Neurite
Un neurite fait référence à toute projection du corps cellulaire d'un neurone. Cette projection peut être soit un axone soit une dendrite. Ce terme est fréquemment utilisé dans le cadre de la biologie du développement en parlant de cellules nerveuses immatures, notamment en culture, car il est alors difficile de distinguer les deux types de prolongements[1].
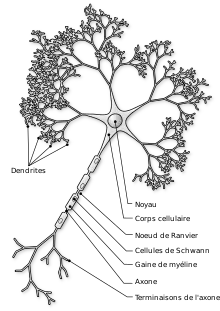
Développement et croissance des neurites
Le développement d'un neurite nécessite l'interaction complexe de signaux extracellulaires et intracellulaires. Le long d'un neurite se trouve des récepteurs détectant les signaux de croissance positifs et négatifs[2]. Le neurite en développement additionne ces signaux de croissance afin de déterminer dans quelle direction ce dernier doit se développer[2]. Bien que tous les signaux de croissance ne soient pas encore connus, plusieurs ont été identifiés et caractérisés. Parmi ces signaux extracellulaire connus, on trouve la nétrine, la sémaphorine, l'éphrine et la collapsine, qui sont tous des inhibiteurs de la croissance des neurites[2] - [3] - [4].
Les jeunes neurites sont souvent remplis de faisceaux de microtubules dont la croissance est stimulée par des facteurs neurotrophiques, tels que le facteur de croissance nerveuse (NGF)[5]. Les protéines Tau permettent elles d'aider à la stabilisation des microtubules en se liant à ces derniers, les protégeant des protéines de coupure[6]. Même après la stabilisation des microtubules, le cytosquelette du neurone reste dynamique.
Dans le neurite qui deviendra l'axone, les filaments d'actine conservent leurs propriétés dynamiques afin de pousser les faisceaux de microtubules vers l'extérieur pour étendre l'axone[7]. Dans tous les autres neurites cependant, les filaments d'actine sont stabilisés par la myosine ce qui empêche le développement de plusieurs axones[8].
La croissance des neurites peut aussi être induite par la molécule d'adhésion des cellules neurales (N-CAM). Cette dernière peut se combiner avec une autre N-CAM (dimérisation) et se fixer sur un récepteur du facteur de croissance des fibroblastes pour en stimuler l'activité tyrosine kinase[9].
Pour finir, des champs électriques endogènes de faible intensité peuvent être utilisés pour faciliter et diriger la croissance des neurites, démontré chez les modèles murins et xénopes. La co-culture de neurones avec du tissu glial électriquement aligné permet également de diriger la croissance des neurites grâce à sa richesse en neurotrophines. .
Établissement de la polarité du neurone
In vitro
Si l'on place un neurone de mammifère indifférencié en culture, ce dernier rétractera tous ses neurites qu'il a déjà développés[10].
Après une mise en culture pendant 0,5 à 1,5 jours, plusieurs neurites mineurs commenceront à sortir du corps cellulaire de ce neurone[10]. Entre le jour 1,5 et le jour 3, l'un des neurites mineurs commencera à devenir considérablement plus grand que les autres, ce neurite finira par devenir l'axone . Aux jours 4 à 7, les neurites mineurs restants commenceront à se différencier en dendrites[10]. Au jour 7, le neurone devient complètement polarisé, avec des dendrites et un axone fonctionnels[10].
In vivo
Un neurite croissant in vivo est entouré de milliers de signaux extracellulaires qui, à leur tour, peuvent être modulés par des centaines de voies intracellulaires. Les mécanismes par lesquels ces signaux chimiques affectent la différenciation des neurites in vivo ne sont pas tous compris avec précision. Néanmoins, l'on sait que 60% du temps le premier neurite qui dépasse du corps cellulaire deviendra l'axone, 30% du temps ce premier neurite ne deviendra pas l'axone, et 10% du temps le neurite qui deviendra l'axone dépasse du corps cellulaire en même temps qu'un ou plusieurs autres neurites[10].
L'hypothèse du "touch and go" tente d'expliquer la mise en place de cette polarité. Elle propose l'idée qu'un neurite mineur puisse s'étendre et grandir vers l'extérieur jusqu'au contact avec un axone déjà développé d'un autre neurone. À ce stade, le neurite commencerait à se différencier en un axone[10]. Cependant, ce modèle n'explique pas comment le premier axone à l'origine des autres puisse se développer.
Quels que soient les signaux extracellulaires impliqués dans la formation d'axones, ils sont transmit par, au moins, 4 voies différentes :
- La voie Rac-1
- La voie médiée par Ras
- La voie cAMP-foie kinase B1
- La voie protéine kinase calcium/calmoduline-dépendante [10]
Une déficience dans l'une de ces voies peut conduire à l'incapacité de développer un axone[10].
Après avoir formé l'axone, le neurone doit empêcher tous les autres neurites de devenir également des axones. C'est ce qu'on appelle l'inhibition globale[10]. Il a été suggéré que l'inhibition globale est obtenue par un signal rétroactif négatif libéré par l'axone nouvellement développé et dirigé vers les autres neurites[11]. Cependant, aucune molécule de signalisation n'a été découverte[10].
Alternativement, il a été suggéré que l'accumulation de facteurs de croissance axonale dans le neurite devenant l'axone induit l'épuisement du dit facteur de croissance pour les autres neurites[12]. Cela forcerait les autres neurites à devenir des dendrites à la suite de concentrations insuffisantes de facteurs de croissance[12].
Voir également
Liens externes
Notes et références
- Flynn, « The cytoskeleton and neurite initiation », Bioarchitecture, vol. 3, no 4, , p. 86–109 (ISSN 1949-0992, PMID 24002528, PMCID 4201609, DOI 10.4161/bioa.26259)
- Valtorta et Leoni, « Molecular mechanisms of neurite extension », Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, vol. 354, no 1381, , p. 387–394 (ISSN 0962-8436, PMID 10212488, PMCID 1692490, DOI 10.1098/rstb.1999.0391)
- Niclou, Franssen, Ehlert et Taniguchi, « Meningeal cell-derived semaphorin 3A inhibits neurite outgrowth », Molecular and Cellular Neurosciences, vol. 24, no 4, , p. 902–912 (ISSN 1044-7431, PMID 14697657, DOI 10.1016/s1044-7431(03)00243-4)
- Luo, Raible et Raper, « Collapsin: a protein in brain that induces the collapse and paralysis of neuronal growth cones », Cell, vol. 75, no 2, , p. 217–227 (ISSN 0092-8674, PMID 8402908, DOI 10.1016/0092-8674(93)80064-l)
- Bear, Mark F; Connors, Barry W.; Paradiso, Michael A., Neuroscience, Exploring the Brain, Philadelphia : Lippincott Williams & Wilkins; Third Edition (February 1, 2006). (ISBN 0-7817-6003-8)
- Qiang, Yu, Andreadis et Luo, « Tau Protects Microtubules in the Axon from Severing by Katanin », The Journal of Neuroscience, vol. 26, no 12, , p. 3120–3129 (ISSN 0270-6474, PMCID 6674103, DOI 10.1523/JNEUROSCI.5392-05.2006, lire en ligne)
- (en) Xiao, Peng, Wan et Tang, « The Atypical Guanine Nucleotide Exchange Factor Dock4 Regulates Neurite Differentiation through Modulation of Rac1 GTPase and Actin Dynamics », Journal of Biological Chemistry, vol. 288, no 27, , p. 20034–20045 (ISSN 0021-9258, PMID 23720743, PMCID 3707701, DOI 10.1074/jbc.M113.458612)
- Toriyama, Kozawa, Sakumura et Inagaki, « Conversion of a signal into forces for axon outgrowth through Pak1-mediated shootin1 phosphorylation », Current Biology, vol. 23, no 6, , p. 529–534 (ISSN 1879-0445, PMID 23453953, DOI 10.1016/j.cub.2013.02.017)
- (en) Vladimir Berezin, Structure and Function of the Neural Cell Adhesion Molecule NCAM, Springer Science & Business Media, (ISBN 978-1-4419-1170-4, lire en ligne)
- (en) Takano, Xu, Funahashi et Namba, « Neuronal polarization », Development, vol. 142, no 12, , p. 2088–2093 (ISSN 0950-1991, PMID 26081570, DOI 10.1242/dev.114454)
- (en) Arimura et Kaibuchi, « Neuronal polarity: from extracellular signals to intracellular mechanisms », Nature Reviews Neuroscience, vol. 8, no 3, , p. 194–205 (ISSN 1471-003X, PMID 17311006, DOI 10.1038/nrn2056)
- (en) Inagaki, Toriyama et Sakumura, « Systems biology of symmetry breaking during neuronal polarity formation », Developmental Neurobiology, vol. 71, no 6, , p. 584–593 (ISSN 1932-846X, PMID 21557507, DOI 10.1002/dneu.20837)