Éphrine
Les éphrines, également connues sous les termes de ligands d'éphrine ou de protéines interagissant avec le récepteur de la famille Eph, sont une famille de protéines qui servent de ligands au récepteur Eph. Les récepteurs Eph, à leur tour, composent la plus grande sous-famille connue des protéines de type récepteur à activité tyrosine kinase (RTK). Le couplage éphrine/Eph, est uniquement actif sur les interactions de cellule à cellule de type directe.
La signalisation Eph/éphrine régule une variété de processus biologiques pendant le développement embryonnaire, dont notamment le guidage des cônes de croissance de l'axone, la formation des limites des tissus, la migration cellulaire, ou encore la segmentation.
Par ailleurs, d'autre fonctions spécifiques à la signalisation éphrine/Eph, et se développant au cours de l'âge adulte, ont été récemment observées et caractérisées. Il s'agit essentiellement de la potentialisation à long terme, de l'angiogenèse et de la différenciation des cellules souches.
Description et caractéristiques
Les éphrines, également dénommées « ligands d'éphrine » ou encore « protéines interagissant avec le récepteur de la famille Eph », sont une famille de protéines destinées à servir de ligands au récepteur Eph[1]. Les récepteurs Eph, à leur tour, composent la plus grande sous-famille connue des protéines de type récepteur à activité tyrosine kinase (RTK)[1] - [2].
Les ligands d'éphrine et les récepteurs Eph (Ephs) se présentant comme des protéines à la fois associées à la membrane, à la liaison et à l'activation des voies de signalisation intracellulaire, les formations du couple Eph/éphrine ne peuvent uniquement se produire que par le biais d'une interaction directe de cellule à cellule[1].
Ces protéines transmembranaires sont constituées d'un nombre d'acides aminés variant entre 205 et 340, selon leurs classes ou leurs structures[3]. Par ailleurs, en raison de leur nature transmembranaire, ces molécules protéiques, à contrario de nombreuses autres protéines ou familles de protéines, ne sont pas soumises à sécrétion cellulaire[4].
La signalisation Eph/éphrine régule une variété de processus biologiques pendant le développement embryonnaire, y compris le guidage des cônes de croissance de l'axone[5], la formation des limites des tissus[6], la migration cellulaire et la segmentation[7] - [2] - [4].
En outre, la signalisation d'Eph/éphrine a récemment été identifiée comme jouant un rôle essentiel dans la maintenance de plusieurs processus survenant l'âge adulte, y compris celui de la potentialisation à long terme[8] l'angiogenèse et la différenciation des cellules souches[9], mais également celles des cellules indifférenciées[10].
Classification
Les ligands d'éphrine sont divisés en deux sous-classes : les éphrine A et les éphrine B[11] - [12]. Ces deux sous-classes ont été déterminées en fonction de leur structure et de leur liaison avec la membrane cellulaire[11] - [12]. Les éphrines A sont ancrées à la membrane par une liaison glycosylphosphatidylinositol (GPI) et ne possèdent pas de domaine cytoplasmique tandis que les éphrines B sont attachées à la membrane par un seul domaine transmembranaire qui contient un court motif cytoplasmique de liaison au PDZ (en)[11] - [13]. Les gènes qui codent les protéines éphrines A et éphrine B sont désignés respectivement sous les termes de « EFNA » et « EFNB ». En parallèle, les récepteurs Eph sont à leur tour classés EphA ou EphB en fonction de leur affinité de liaison pour les ligands éphrine A ou éphrine B[14] - [12].
Parmi les huit types d'éphrines ayant été identifiés chez l'homme, il existe cinq ligands éphrine A connus (dénommés « éphrine A 1 à 5 ») et qui interagissent avec neuf EphAs (« EphA 1 à 8 et EphA10 »)[8] - [15] - [12]. La seconde sous-classe est constituée de trois ligands de type éphrine B (« éphrine B 1 à 3 »), lesquels interagissent avec cinq EphB (« EphB1-4 et EphB6 »)[8] - [15] - [12]. Les Ephs d'une sous-classe particulière démontrent une capacité à se lier avec une affinité élevée à toutes les éphrines de la sous-classe correspondante, mais en général ont peu ou pas de liaison croisée aux éphrines de la sous-classe opposée[16]. Cependant, il existe quelques exceptions à cette spécificité de liaison intra-classe, car il a récemment été démontré que l'éphrine B3 est capable de se lier et d'activer le récepteur EPH A4 et l'éphrine A5 peut se lier et activer le récepteur Eph B2[17]. Le couple EphA/éphrine A se révélant comme étant généralement lié avec par une affinité élevée, ce qui peut être partiellement attribuable à une interaction entre les éphrines A et les EphAs par un mécanisme dit de « lock-and-key » ; ce processus nécessitant peu de changement conformationnel des EphA sur la liaison du ligand. En revanche, les EphB se lient généralement avec une affinité inférieure à celle d'EphA/éphrine A car ils utilisent un mécanisme « induit » qui requiert un changement conformationnel plus important d'EphB pour lier les éphrines B[18].
Les éphrines de conformation B1 et B2, se révèlent être de type pro-angiogénique et permettent la multiplication des cellules endothéliales[19]. Par ailleurs, bien que son récepteur EphB4 soit uniquement présent au niveau des réseaux veineux, l'intervention de l'éphrine B2 demeure restreinte aux seuls domaines artériels[19].
Fonctions liées à des cellules nerveuses
Guidage axonal
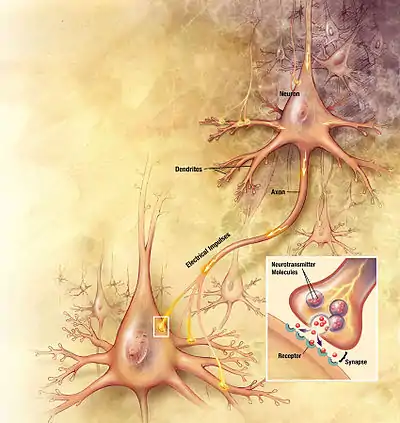
Au cours du développement du système nerveux central (ou neurodéveloppement), la signalisation d'Eph/éphrine joue un rôle essentiel dans la migration de cellule à cellule de plusieurs types d'axones neuronaux vers leurs organes cibles[20] - [21]. La signalisation Eph/éphrine contrôle le guidage des axones neuronaux par leur capacité à inhiber la survie des cônes de croissance axonale, qui repousse l'axone migrant loin du site de l'activation Eph/éphrine[20]. Les cônes de croissance des axones migrateurs ne répondent pas simplement aux niveaux absolus d'Ephs ou d'éphrines dans les cellules qu'ils contactent, mais répondent aux niveaux relatifs de l'expression d'Eph et d'éphrine[22], ce qui permet aux axones migrateurs qui expriment les Ephs ou les éphrines d'être dirigés le long de gradients d'Eph ou d'éphrine exprimés par les cellules vers une destination où la survie axiale du cône de croissance n'est plus complètement inhibée[20].
Bien que l'activation de l'Eph/éphrine soit habituellement associée à la diminution de la survie du cône de croissance et à la répulsion des axones migrateurs, il a récemment été démontré que la survie du cône de croissance ne dépend pas uniquement de l'activation de l'Eph/éphrine, mais plutôt des effets différentiels de la signalisation « avant » par le récepteur Eph ou la signalisation synaptique « inverse » par le ligand de l'éphrine sur la survie du cône de croissance[20] - [23].
D'autre part, des chercheurs ont récemment pu déterminé que l'expression de l'éphrine A5 couplée à son récepteur Eph A5, via l'ARNm, est impliquée dans l'installation le système mésostriatal (en)[21]. La paire éphrine A5/Eph A5 intervient dans le guidage d'axone notamment au sein du tectum mesencephali avant — c'est-à-dire la partie médio-ventral du mésencéphale, ou « cerveau moyen » —, mais également dans la strate de oriens (en) (située dans l'hippocampe)[24] et le thalamus[21]. Cette interaction, qui affecte le développement cérébral au cours de la phase embryonnaire, concerne les neurones de type dopaminergiques[21]. En outre, cette étude a conduit à mettre en évidence qu'un dysfonctionnement de l'activation du couple éphrine A5/Eph A5, pourrait être impliqué dans la maladie de Parkinson[21].
Enfin, une interaction d'adhérence entre les protéines éphrines B et leurs récepteurs associés (EphB) induisent le début de la différenciation des aires pré-synaptiques et de leurs densités après contact synaptique[25].
Signalisation inverse
L'une des rares propriétés observées sur les ligands d'éphrine se révélerait être, pour nombre d'entre eux, la capacité d'initier un signal « inverse » distinct et différent du signal intracellulaire activé dans les cellules exprimant le récepteur Eph[1] - [26]. Bien que les mécanismes par lesquels la signalisation inverse se produisent ne soient pas complètement acquis et appréhendés, il a été démontré que l'éphrine-A et l'éphrine-B ont des réponses cellulaires distinctes de celles associées à l'activation de leurs récepteurs correspondants[1] - [26]. Plus précisément, il a été établi que l'éphrine A5 stimule la propagation du cône de croissance dans les neurones moteurs de la colonne vertébrale[20] ; tandis que l'éphrine-B1, quant à elle, favorise la maturation de l'épine dendritique[27].
Cartographie rétinotopique
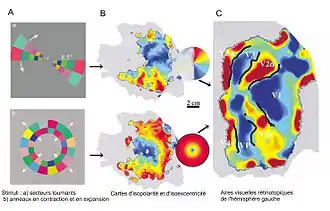
La formation d'une carte rétinotopique, qui s'organise au niveau du colliculus supérieur (« CS ») (structure également connue sous le terme de tectum optique chez les vertébrés inférieurs), requiert une migration appropriée des axones des cellules ganglionnaires rétiniennes (« RGC ») de la rétine vers des régions spécifiques du « CS »[28]. Ce processus rétinien fait l'objet d'une médiation par des gradients d'expression d'Eph et d'éphrine non-seulement dans le « CS », mais également au sein des « RGC » migrateurs quittant la rétine[28]. La diminution de la survie des cônes de croissance axonaux précédemment cités, permet un gradient d'expression du ligand de l'éphrine A dans l'axe de référence postérieur à bas-antérieur dans le CS. Ce mécanisme dirige les axones des RGC migrateurs — ces derniers exprimant alors un niveau élevé de récepteurs EphA — à partir de la région temporelle de la rétine, vers des cibles dans le CS antérieur et les RGC de la rétine nasale qui ont une faible expression EphA vers leur destination finale dans le CS postérieur[29] - [30] - [31]. De même, un gradient d'expression de l'éphrine B1 le long de l'axe médial-ventral du CS, activant ainsi l'EphB dans les aires dorsales et ventrales, dirige respectivement la migration des RGC vers les zones du colliculus supérieur latérales et médianes[32]. De manière générale, pendant la mise en place de la carte rétinotopique au niveau du tectum, le gradient d'interaction éphrine/Eph est très étroitement lié au facteur d'Engrailed, lequel a été auparavant fixé lors de la signalisation axonale[33]. D'autre part, dans un contexte d'arrêt de l'activation d'éphrine B2, au sein du chiasma optique, les croissances axonales sont redirigées vers la partie hétérolatérale de la rétine[34]. À terme, cette anomalie fonctionnelle constitue et aboutie à la mise en place d'un champ visuel de type monoculaire[34].
Élagage synaptique
De récents travaux ont mis en évidence que la signalisation entre l'éphrine et son récepteur Eph, permet de réguler et orienter les branches axiales de la rétine[29] - [30] - [31]. La signalisation ventral avant entre l'éphrine A et l'EphA, suivant l'axe antérieur-postérieur, paraît agir comme un facteur inhibiteur lors de la formation de la branche axiale de la rétine postérieure avec l'aire terminale de l'axone[13]. L'interaction directe éphrine/Eph stimulerait également l'élagage synaptique des axones ayant rejoint et affecté cette même aire terminale[13].
En outre, le processus de signalisation inverse des éphrines B et des EphB induit un phénomène de rétractation au sein du faisceau intrapyramidal[35]. En outre, des observations ont montré que la molécule Ephrin-B3 (en) transforme des signaux inverses dépendant de la phosphorylation de la tyrosine en axones de l'hippocampe qui déclenchent l'élagage des fibres du faisceau intrapyramidal excessives[35] - [36]. Ces éléments tendent à déterminer que les récepteurs EphB, qui s'expriment à la surface des cellules cibles, engendrerait la phosphorylation du domaine tyrosine de l'éphrine B3[35] - [36]. L'interaction entre l'éphrine B3 et la protéine adaptatrice de type cytoplasmique NCK2 génère le recrutement et le couplage de la protéine Dock « 180 » avec les domaines actifs kinases via les enzymes PKA[37] - [38]. À terme, ce mécanisme produit une résorption axonale, ce processus conduisant à son tour à un possible élagage synaptique[39] - [40] - [37] - [38].
Fonctions liées à d'autres types de cellules
Angiogenèse
Les éphrines favorisent l'angiogenèse dans des conditions physiologiques et pathologiques — par exemple : l'angiogenèse du cancer et plus spécifiquement la néovascularisation lors d'une malformation artério-veineuse cérébrale —[41] - [42] - [43]. En particulier, éphrine-B2 et EphB4 déterminent respectivement le devenir artériel et veineux des cellules endothéliales, bien que la régulation de l'angiogenèse atténue l'expression au sein de la voie de signalisation du facteur de croissance de l’endothélium vasculaire (protéine également connue sous l'abréviation anglophone VEGF)[44] - [42] - [43]. L'éphrine-B2 affecte les récepteurs du VEGF (comme le VEGFR3) par des voies de signalisation avant et arrière[44]. Le cheminement de l'éphrine-B2 s'étend à la lymphangiogenèse, conduisant à l'internalisation du VEGFR3 dans les cellules endothéliales lymphatiques cultivées[44]. Bien que le rôle des éphrines dans l'angiogenèse du développement soit établie, ce dernier, au sein du processus de l'angiogenèse tumorale, demeure encore incertain et n'a pas encore été formellement caractérisé et précisé[45]. Sur la base d'observations effectuées sur des souris déficitaires en éphrine-A2, il a été déterminé que cette protéine peut fonctionner dans la signalisation directe de l'angiogenèse tumorale[45]. Néanmoins, cette éphrine ne contribue pas aux déformations vasculaires au cours du développement[45]. En outre, Ephrin-B2 et EphB4 peuvent également contribuer à l'angiogenèse tumorale en surcroît de leurs interventions dans le développement embryonnaire, bien que leur mécanisme précis reste indéterminé[45]. Les paires de récepteurs éphrine B2/EphB4 et éphrine B3/EphB1 contribuent davantage à la vasculogenèse (en) qu'à celle de l'angiogenèse[46]. À contrario, le couple éphrine A1/EphA2 semble contribuer exclusivement à l'angiogenèse[46].
Par ailleurs, il a été également déterminé qu'une mutation de l'allèle codant le liguand DLL4 (en) conduit à une inhibition de plusieurs marqueurs artériels, dont l'éphrine B2[43]. Ce défaut produit à son tour une disparition de la voie de signalisation Notch, transduction essentielle à un système vasculaire efficient[43].
Il a été mis en évidence que plusieurs types d'interactions éphrines et Eph se sont révélés régulés à la hausse dans les cancers humains, y compris les cancers du sein, du côlon et du foie[46]. En contre-point, la régulation négative d'autres types d'éphrines et de leurs récepteurs peut également contribuer à la tumorigénéité ; à savoir EphA1 pour le cancer colorectal et EphB6 pour le mélanome[46]. Dans un contexte d'observation similaire, il a été démontré que différentes éphrines intègrent des voies pourvues de mécanismes identiques afin de compléter la croissance de différentes structures cellulaires[46].
Facteur d'initiation dans la migration des cellules épithéliales de l'intestin
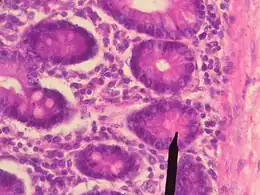
La famille de protéines d'éphrine de classe A et de classe B guide les ligands avec les récepteurs de la surface cellulaire de la famille EphB pour fournir une migration stable, ordonnée et spécifique des cellules épithéliales intestinales des cryptes de Lieberkühn au sein des villosités. Les protéines de type Wnt déclenchent l'expression des récepteurs EphB au fond de la crypte, entraînant une diminution de l'expression de l'Eph et une augmentation de l'expression du ligand de l'éphrine, plus le placement d'une cellule progénitrice est superficiel[47]. La migration est causée par un mécanisme de signalisation bidirectionnelle dans lequel l'engagement du ligand d'éphrine avec le récepteur EphB régule la dynamique du cytosquelette de l'actine pour provoquer une « répulsion ». Les cellules restent en place une fois que l'interaction cesse de s'arrêter. Alors que le mucus sécrétant les cellules en gobelet et les cellules absorbantes se déplacent vers la lumière, les cellules matures de Paneth se déplacent dans la direction opposée, au fond de la crypte, où elles résident[48]. À l'exception de la liaison du ligand d'éphrine à EphA5, toutes les autres protéines de classe A et B ont été trouvées dans l'intestin. Cependant, les protéines d'éphrine A4, A8, B2 et B4 ont les niveaux les plus élevés en phase fœtale et diminuent avec l'âge.
L'expérience réalisée avec des souris mutantes à récepteur Eph a révélé un trouble dans la distribution de différents types de cellules[48]. Des cellules absorbantes de différenciation différente ont été mélangées avec les cellules souches dans les villosités. Sans le récepteur, le ligand d'Ephrin s'est révélé insuffisant pour le placement correct des cellules[49]. Des études récentes, réalisées sur des souris dont l'ephrine est obsolète, ont également montré des preuves du rôle indirect de l'interaction ephrine-eph dans la suppression du cancer colorectal. Le développement de polypes adénomateux créé par la prolifération incontrôlée de cellules épithéliales est contrôlé par l'interaction ephrine-eph. Les souris possédant une mutation APC, autrement dit sans protéines ephrin-B, n'ont pas les moyens d'empêcher la propagation de cellules tumorales positives à l'ephB tout au long de la jonction crypte-villosité[50]. De même, au sein de l'organisme des souris possédant une éphrine B2 dite « knock-out » — ou déficiente, obsolète —, il a été établi que cette anomalie engendre d'importants déficits vasculaires[51].
En parallèle de ces éléments, des recherches effectuées sur des oursins ont permis de constater que la signalisation du couple éphrine/Eph intervient dans la migration et l'insertion des immunocytes au sein de l'épythélium[52].
Stimulateur des interactions entre cellules osseuses
De récents travaux de recherches, effectués sur des souris de laboratoire, ont permis d'établir que les interactions de cellulo-cellulaires de types osseux, processus indispensables à la formation des os (autrement dit l'ostéogenèse), ne pouvaient se réaliser, ou en tous les cas être correctement maintenues, sans l'activation du couple éphrine/Eph[53]. De manière générale, lors d'une ostéogenèse en phase embryonnaire, l'association des ligands d'éphrine et de leurs récepteurs permet de réguler la migration, le développement et l'adhésion des cellules souches de type mésenchymateuse[53]. Des études, effectuées par technique de mise en culture cellulaire, ont mis en évidence que des éphrines B1 non-fonctionnelles peuvent notamment induire une dysplasie craniofrontonasale (en), une pathologie qui fait partie des maladies rares[53]. Ce type de maladie rare est observée chez l'espèce humaine mais également chez les rongeurs[53].
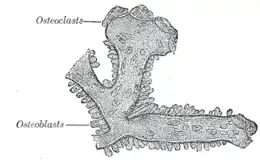
Les éphrines B2 activent la phase remodelage osseux[53]. Dans ce contexte, l'envoi du signal bidirectionnel[Note 1] se produisant entre l'éphrine B2 et l'EphB4, récepteur présent à la surface des ostéoblastes, annule et détruit la résorption osseuse des cellules ostéoclastiques — monocytes de type macrophages —[53] - [55]. Ce mécanisme permet alors de faciliter la formation osseuse ostéoblastique, processus conduisant à une transition normale entre deux ces deux états (résorption puis formation osseuses)[53]. En amont, l'hormone parathyroïdienne, également connue sous le nom de « parathormone », active l'éphrine B2 sur la surface des cellules ostéoblastiques[53]. Par conséquent, cette hormone participe également à la formation des ostéoblastes[53].
D'autre part, à contrario de l'éphrine B2, l'éphrine de type A2 se comporte comme un « inhibiteur de couplage »[53]. Au cours de l'ostéognèse, ce facteur d'inhibition caractéristique de l'éphrine A2 trouve son origine dans sa capacité de produire une signalisation inverse[53]. Or cette même signalisation inverse favorise la croissance et le développement des ostéoclastes[53]. Cet autre mécanisme, induit par l'activation du couple éphrine A2/Eph A2 à la surface des ostéoblastes, annule ainsi la formation et la minéralisation efficiente des os crâno-frontonasal[53].
Enfin, un dysfonctionnement des éphrines et de leurs récepteurs Eph peut également engendrer l'apparition de pathologies tels que l'arthrose, la polyarthrite rhumatoïde, le myélome multiple et l'ostéosarcome[53].
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Ephrin » (voir la liste des auteurs).
Notes
- À l'instar de la signalisation inverse, celle de type bidirectionnelle se révèle être l'une des spécificités caractérisant le couplage éphrine/Eph[54].
Références
- (en) Erika M. Lisabeth, Giulia Falivelli et Elena B. Pasquale, « Eph Receptor Signaling and Ephrins », Cold Spring Harber Perspectives Biology, vol. 9, no 5, 2013 sep (PMCID PMC3753714, DOI 10.1101/cshperspect.a009159, lire en ligne, consulté le ).
- (en) Rüdiger Klein, « Eph/ephrin signalling during development », Development, no 139, , p. 4105-4109 (DOI 10.1242/dev.074997, lire en ligne, consulté le ).
- Jean Foucrier et Guillaume Bassez, « Molécules membranaires : Éphrine/Eph », dans Jean Foucrier et Guillaume Bassez, PACES UE2 Reproduction et Embryologie, Ediscience, , 368 p. (lire en ligne), page 130.
- Pierre Corvol et Nicolas Postel-Vinay, « Naissance : le vaisseau embryonnaire », dans Pierre Corvol et Nicolas Postel-Vinay, L’arbre vasculaire : Nouvelles voies de guérison, Odile Jacob, , 288 p. (lire en ligne), page 63.
- (en) J. Egea et R. Klein, « Bidirectional Eph-ephrin signaling during axon guidance », Trends in Cell Biology, vol. 17, no 5, , pages 230–238 (PMID 17420126, DOI 10.1016/j.tcb.2007.03.004).
- (en) N. Rohani, L. Canty, O. Luu, F. Fagotto et R. Winklbauer, « EphrinB/EphB signaling controls embryonic germ layer separation by contact-induced cell detachment », PLoS Biology, Hamada Hiroshi, vol. 9, no 3, , e1000597 (PMID 21390298, PMCID 3046958, DOI 10.1371/journal.pbio.1000597).
- (en) A. Davy et P. Soriano, « Ephrin signaling in vivo : look both ways », Developmental Dynamics, vol. 232, no 1, , p. 1–10 (PMID 15580616, DOI 10.1002/dvdy.20200).
- (en) K. Kullander et R. Klein, « Mechanisms and functions of Eph and ephrin signalling », Nature Reviews Molecular Cell Biology, vol. 3, no 7, , pages 475 à 486 (PMID 12094214, DOI 10.1038/nrm856).
- (en) S. Kuijper, C.J. Turner et R.H. Adams, « Regulation of angiogenesis by Eph-ephrin interactions », Trends in Cardiovascular Medicine, vol. 17, no 5, , p. 145–151 (PMID 17574121, DOI 10.1016/j.tcm.2007.03.003).
- (en) M. Genander et J. Frisén, « Ephrins and Eph receptors in stem cells and cancer », Current Opinion in Cell Biology, vol. 22, no 5, , p. 611–616 (PMID 20810264, DOI 10.1016/j.ceb.2010.08.005).
- (de) Christoph Wagener et Oliver Müller, « EPH/Ephrin system », dans Christoph Wagener et Oliver Müller, Molekulare Onkologie : Entstehung, Progression, klinische Aspekte ; 95 Tabellen, Georg Thieme Verlag, , 404 p. (lire en ligne), pages 372 et 373.
- Larsen et al. 2017, p. 128.
- (en) Dina Arvanitis et Alice Davy, « Eph/ephrin signaling : networks », Genes & Development, (DOI 10.1101/gad.1630408, lire en ligne, consulté le ).
- (en) Collectif, « Unified Nomenclature for Eph Family Receptors and Their Ligands, the Ephrins », Cell, vol. 90, no 3, , pages 403 et 404 (DOI 10.1016/S0092-8674(00)80500-0, lire en ligne, consulté le ).
- (en) M.E. Pitulescu et R.H. Adams, « Eph/ephrin molecules--a hub for signaling and endocytosis », Genes & Development, vol. 24, no 22, , p. 2480–2492 (PMID 21078817, PMCID 2975924, DOI 10.1101/gad.1973910).
- (en) E.B Pasquale, « The Eph family of receptors », Current Opinion in Cell Biology, vol. 9, no 5, , p. 608–615 (PMID 9330863, DOI 10.1016/S0955-0674(97)80113-5).
- (en) J.P. Himanen, M.J. Chumley, M. Lackmann, C. Li et al., « Repelling class discrimination : ephrin-A5 binds to and activates EphB2 receptor signaling », Nature Neuroscience, vol. 7, no 5, , p. 501–509 (PMID 15107857, DOI 10.1038/nn1237).
- (en) J.P. Himanen, « Ectodomain structures of Eph receptors », Seminars in Cell & Developmental Biology, vol. 23, no 1, , p. 35–42 (PMID 22044883, DOI 10.1016/j.semcdb.2011.10.025).
- Jean-Yves Artigou et Jean-Jacques Monsuez, « Biologie et physilogie cardiovasculaire et de l'hémostase : Différenciation vasculaire en veine ou en artère », dans Jean-Yves Artigou et Jean-Jacques Monsuez, Cardiologie et maladies vasculaires, Elsevier Masson, , 1639 p. (lire en ligne), page 19.
- (en) T. Marquardt, R. Shirasaki, S. Ghosh, S.E. Andrews et al., « Coexpressed EphA receptors and ephrin-A ligands mediate opposing actions on growth cone navigation from distinct membrane domains », Cell, vol. 121, no 1, , p. 127–139 (PMID 15820684, DOI 10.1016/j.cell.2005.01.020).
- Claire Deschamps, Étude de l'expression de la molécule de guidage éphrine-A5 dans le cerveau de souris au cours du développement : implication dans la mise en place de la voie mésostriatale, Université de Poitiers, , 224 p. (lire en ligne [PDF]).
- (en) M. Reber, P. Burrola et G. Lemke, « A relative signalling model for the formation of a topographic neural map », Nature, vol. 431, no 7010, , p. 847–853 (PMID 15483613, DOI 10.1038/nature02957).
- (en) T.J. Petros, J.B. Bryson et C. Mason, « Ephrin-B2 elicits differential growth cone collapse and axon retraction in retinal ganglion cells from distinct retinal regions », Developmental Neurobiology, vol. 70, no 11, , p. 781–794 (PMID 20629048, PMCID 2930402, DOI 10.1002/dneu.20821).
- (en) Gianmaria Maccaferri, « Stratum oriens horizontal interneurone diversity and hippocampal network dynamics », The Journal of Physiology, vol. 562, , p. 73 à 80 (PMCID 1665470, DOI 10.1113/jphysiol.2004.077081, lire en ligne, consulté le ).
- George J. Augustine, David Fitzpatrick, William Hall, Anthony-Samuel LaMantia et al., « La formation des cartes topographiques », dans George J Augustine, David Fitzpatrick, William Hall, Anthony-Samuel LaMantia, Dale Purves et Léonard White, Neurosciences, De Boeck Superieur, , 848 p. (lire en ligne), page 522.
- Larsen et al. 2017, p. 246.
- (en) I. Segura, C.L. Essmann, S. Weinges et A. Acker-Palmer, « Grb4 and GIT1 transduce ephrinB reverse signals modulating spine morphogenesis and synapse formation », Nature Neuroscience, vol. 10, no 3, , p. 301–310 (PMID 17310244, DOI 10.1038/nn1858).
- (en) J.W. Triplett et D.A. Feldheim, « Eph and ephrin signaling in the formation of topographic maps », Seminars in Cell & Developmental Biology, vol. 23, no 1, , p. 7–15 (PMID 22044886, PMCID 3288406, DOI 10.1016/j.semcdb.2011.10.026)
- (en) D.G. Wilkinson, « Multiple roles of EPH receptors and ephrins in neural development », Nature Reviews. Neuroscience, vol. 2, no 3, , p. 155–164 (PMID 11256076, DOI 10.1038/35058515).
- (en) H.J Cheng, M. Nakamoto, A.D. Bergemann et J.G. Flanagan, « Complementary gradients in expression and binding of ELF-1 and Mek4 in development of the topographic retinotectal projection map », Cell, vol. 82, no 3, , p. 371–381 (PMID 7634327, DOI 10.1016/0092-8674(95)90426-3).
- (en) U. Drescher, C. Kremoser, C. Handwerker et al., « In vitro guidance of retinal ganglion cell axons by RAGS, a 25 kDa tectal protein related to ligands for Eph receptor tyrosine kinases », Cell, vol. 82, no 3, , p. 359–370 (PMID 7634326, DOI 10.1016/0092-8674(95)90425-5).
- (en) F. Mann, S. Ray, W. Harris et C. Holt, « Topographic mapping in dorsoventral axis of the Xenopus retinotectal system depends on signaling through ephrin-B ligands », Neuron, vol. 35, no 3, , p. 461–473 (PMID 12165469, DOI 10.1016/S0896-6273(02)00786-9).
- J.M.W. Slack, « 16 - Développement du système nerveux : Connexions neuronales dans le système visuel », dans J.M.W. Slack, Biologie du développement, De Boeck Supérieur, , 482 p. (lire en ligne), pages 343 à 345.
- Larsen et al. 2017, p. 222.
- Clara A. Amegandjin, « 2. Récepteurs Eph et éphrines », dans Clara A. Amegandjin, Localisation régionale et subcellulaire du récepteur EphA7 dans l’hippocampe et le cervelet du rat adulte, Université de Montréal, , 117 p. (lire en ligne [PDF]), pages 8 à fin chapitre.
- (en) Dominique Bagnard, Axon Growth and Guidance, Springer Science & Business Media, , 170 p. (lire en ligne), pages 32 à 40.
- (en) Lei Shi, « Dock protein family in brain development and neurological disease », Communicative & Integrative Biology, vol. 6, no 6, (DOI 10.4161/cib.26839, lire en ligne, consulté le ).
- Alexandre Moreau, « III. Rôle de l’interaction Nck2/PAK3 dans la modulation de la transmission synaptique excitatrice : contrôle sérotoninergique de la balance excitation-inhibition dans le cortex visuel. », dans Alexandre Moreau, Neuromodulation des réseaux neuronaux, Université Paris-Sud, (lire en ligne [PDF]), pages 157 à 180.
- (en) N. Xu et M. Henkemeyer, « Ephrin-B3 reverse signaling through Grb4 and cytoskeletal regulators mediates axon pruning », Nature Neuroscience, vol. 12, , p. 268–276 (PMID 19182796, PMCID 2661084, DOI 10.1038/nn.2254).
- (en) Inji Park et Hyun-Shik Lee, « EphB/ephrinB Signaling in Cell Adhesion and Migration », Mol Cells, (PMCID PMC4314128, DOI 10.14348/molcells.2015.2116, lire en ligne, consulté le ).
- (en) O. Salvucci et G. Tosato, « Essential roles of EphB receptors and EphrinB ligands in endothelial cell function and angiogenesis », Advances in Cancer Research, vol. 114, no 2, , p. 21–57 (PMID 22588055, PMCID 3500853, DOI 10.1016/B978-0-12-386503-8.00002-8)
- (en) J. Bai, Y.J. Wang, L. Liu et Y.L. Zhao, « Ephrin B2 and EphB4 selectively mark arterial and venous vessels in cerebral arteriovenous malformation », The Journal of International Medical Research, vol. 42, no 2, , p. 405–15 (PMID 24517927, DOI 10.1177/0300060513478091).
- Jacques Robert, « Ontogenèse vasculaire : Marqueurs moléculaires spécifiques des artères et des veines », dans Jacques Robert, L'Angiogenèse, John Libbey Eurotext, , 152 p. (lire en ligne), pages 22 à 26.
- (en) O. Salvucci et G. Tosato, « Essential roles of EphB receptors and EphrinB ligands in endothelial cell function and angiogenesis », Advances in Cancer Research, vol. 114, no 2, , p. 21–57 (PMID 22588055, PMCID 3500853, DOI 10.1016/B978-0-12-386503-8.00002-8)
- (en) E.B. Pasquale, « Eph receptors and ephrins in cancer: bidirectional signalling and beyond », Nature Reviews. Cancer, vol. 10, no 3, , p. 165–80 (PMID 20179713, PMCID 2921274, DOI 10.1038/nrc2806).
- (en) Birgit Mosch, Bettina Reissenweber, Christin Neuber et Jens Pietzsch, « Eph Receptors and Ephrin Ligands : Important Players in Angiogenesis and Tumor Angiogenesis », Journal of Oncology, vol. 2010, , p. 1–12 (ISSN 1687-8450, DOI 10.1155/2010/135285).
- (en) Bruce Alberts, Alexander Johnson, Julian lewis, Raff Martin, Keith Roberts et Peter Walter, Molecular Biology of the Cell, Garland Sciences, (ISBN 978-0-8153-4105-5), pages 1440–1441.
- (en) Eduard Batlle, « Wnt signalling and EphB-ephrin interactions in intestinal stem cells and CRC progression », Scientific Report, (lire en ligne [PDF], consulté le ).
- S. Islam, A.M. Loizides, J.J. Fialkovich, R.J. Grand RJ et R.K. Montgomery, « Developmental expression of Eph and ephrin family genes in mammalian small intestine », Digestive Diseases and Sciences, vol. 55, no 9, , p. 2478–88 (PMID 20112066, PMCID 3947671, DOI 10.1007/s10620-009-1102-z).
- (en) Mara Pitulescu, « Eph/ephrin molecules-a hub for signaling and endocytosis », Genes & Development, (DOI 10.1101/gad.1973910, lire en ligne [PDF], consulté le ).
- J.M.W. Slack, « 19 - Cellules souches et croissance vasculaire : 2.4.1. Les vaisseaux sanguins », dans J.M.W. Slack, Biologie du développement, De Boeck Supérieur, , 482 p. (lire en ligne), page 420.
- (en) Oliver A. Krupke, Ivona Zysk, Dan O. Mellott et Robert D. Burke, « Eph and Ephrin function in dispersal and epithelial insertion of pigmented immunocytes in sea urchins embryons », Research article, (DOI 10.7554/eLife.16000, lire en ligne, consulté le ).
- (en) Koichi Matsuo et Natsuko Otaki, « Bone cell interactions through Eph/ephrin : Bone modeling, remodeling and associated diseases », Cell and Adhesion migration, Taylor & Francis - Landes Biosciences, vol. 6, no 2, , p. 148–156 (PMCID PMC3499314, DOI 10.4161/cam.20888, lire en ligne, consulté le ).
- Larsen et al. 2017, p. 129.
- Roland Baron, « L'ostéoclaste et les mécanismes moléculaires de la résorption osseuse », médecine/sciences, Inserm, vol. 17, no 12, , p. 1260 à 1269 (lire en ligne [PDF], consulté le ).
Pour approfondir
Bibliographie
![]() : document utilisé comme source pour la rédaction de cet article.
: document utilisé comme source pour la rédaction de cet article.
- (en) Dina Arvanitis et Alice Davy, « Eph/ephrin signaling : networks », Genes & Development, (DOI 10.1101/gad.1630408, lire en ligne, consulté le ).

- Thomas Jungas, Caractérisation de la signalisation éphrine/Eph dans la division cellulaire, Université de Toulouse, , 239 p. (lire en ligne [PDF]).
- J.M.W. Slack, Biologie du développement, De Boeck Supérieur, , 482 p. (lire en ligne).
- (en) Egea J et Klein R, « Bidirectional Eph-ephrin signaling during axon guidance », Trends in Cell Biology, vol. 17, no 5, , p. 230–238 (PMID 17420126, DOI 10.1016/j.tcb.2007.03.004).
- William Larsen, Philip R. Brauer, Gary C. Schoenwolf et Philippa Francis-West, Embryologie humaine, De Boeck Supérieur, , 560 p. (lire en ligne).
- (en) Koichi Matsuo et Natsuko Otaki, « Bone cell interactions through Eph/ephrin : Bone modeling, remodeling and associated diseases », Cell and Adhesion migration, Taylor & Francis - Landes Biosciences, vol. 6, no 2, , p. 148–156 (PMCID PMC3499314, DOI 10.4161/cam.20888, lire en ligne, consulté le ).
- (en) N. Rohani, L. Canty, O. Luu, F. Fagotto et R. Winklbauer, « EphrinB/EphB signaling controls embryonic germ layer separation by contact-induced cell detachment », PLoS Biology, Hamada Hiroshi, vol. 9, no 3, , e1000597 (PMID 21390298, PMCID 3046958, DOI 10.1371/journal.pbio.1000597).
- (en) N. Rohani, L. Canty, O. Luu, F. Fagotto et R. Winklbauer, « EphrinB/EphB signaling controls embryonic germ layer separation by contact-induced cell detachment », PLoS Biology, Hamada Hiroshi, vol. 9, no 3, , e1000597 (PMID 21390298, PMCID 3046958, DOI 10.1371/journal.pbio.1000597).
- (en) A. Davy et P. Soriano, « Ephrin signaling in vivo : look both ways », Developmental Dynamics, vol. 232, no 1, , p. 1–10 (PMID 15580616, DOI 10.1002/dvdy.20200).
- (en) K. Kullander et R. Klein, « Mechanisms and functions of Eph and ephrin signalling », Nature Reviews Molecular Cell Biology, vol. 3, no 7, , pages 475 à 486 (PMID 12094214, DOI 10.1038/nrm856).
- (en) S. Kuijper, C.J. Turner et R.H. Adams, « Regulation of angiogenesis by Eph-ephrin interactions », Trends in Cardiovascular Medicine, vol. 17, no 5, , p. 145–151 (PMID 17574121, DOI 10.1016/j.tcm.2007.03.003).
- (en) M. Genander et J. Frisén, « Ephrins and Eph receptors in stem cells and cancer », Current Opinion in Cell Biology, vol. 22, no 5, , p. 611–616 (PMID 20810264, DOI 10.1016/j.ceb.2010.08.005).
- (en) Collectif, « Unified Nomenclature for Eph Family Receptors and Their Ligands, the Ephrins », Cell, vol. 90, no 3, , pages 403 et 404 (DOI 10.1016/S0092-8674(00)80500-0, lire en ligne, consulté le ).
- (en) M.E. Pitulescu et R.H. Adams, « Eph/ephrin molecules--a hub for signaling and endocytosis », Genes & Development, vol. 24, no 22, , p. 2480–2492 (PMID 21078817, PMCID 2975924, DOI 10.1101/gad.1973910).
- (en) E.B Pasquale, « The Eph family of receptors », Current Opinion in Cell Biology, vol. 9, no 5, , p. 608–615 (PMID 9330863, DOI 10.1016/S0955-0674(97)80113-5).
- (en) J.P. Himanen, M.J. Chumley, M. Lackmann, C. Li et al., « Repelling class discrimination : ephrin-A5 binds to and activates EphB2 receptor signaling », Nature Neuroscience, vol. 7, no 5, , p. 501–509 (PMID 15107857, DOI 10.1038/nn1237).
- (en) J.P. Himanen, « Ectodomain structures of Eph receptors », Seminars in Cell & Developmental Biology, vol. 23, no 1, , p. 35–42 (PMID 22044883, DOI 10.1016/j.semcdb.2011.10.025).
- (en) T. Marquardt, R. Shirasaki, S. Ghosh, S.E. Andrews et al., « Coexpressed EphA receptors and ephrin-A ligands mediate opposing actions on growth cone navigation from distinct membrane domains », Cell, vol. 121, no 1, , p. 127–139 (PMID 15820684, DOI 10.1016/j.cell.2005.01.020).
- (en) M. Reber, P. Burrola et G. Lemke, « A relative signalling model for the formation of a topographic neural map », Nature, vol. 431, no 7010, , p. 847–853 (PMID 15483613, DOI 10.1038/nature02957).
- (en) T.J. Petros, J.B. Bryson et C. Mason, « Ephrin-B2 elicits differential growth cone collapse and axon retraction in retinal ganglion cells from distinct retinal regions », Developmental Neurobiology, vol. 70, no 11, , p. 781–794 (PMID 20629048, PMCID 2930402, DOI 10.1002/dneu.20821).
- (en) J.W. Triplett et D.A. Feldheim, « Eph and ephrin signaling in the formation of topographic maps », Seminars in Cell & Developmental Biology, vol. 23, no 1, , p. 7–15 (PMID 22044886, PMCID 3288406, DOI 10.1016/j.semcdb.2011.10.026).
- (en) D.G. Wilkinson, « Multiple roles of EPH receptors and ephrins in neural development », Nature Reviews. Neuroscience, vol. 2, no 3, , p. 155–164 (PMID 11256076, DOI 10.1038/35058515).
- (en) H.J Cheng, M. Nakamoto, A.D. Bergemann et J.G. Flanagan, « Complementary gradients in expression and binding of ELF-1 and Mek4 in development of the topographic retinotectal projection map », Cell, vol. 82, no 3, , p. 371–381 (PMID 7634327, DOI 10.1016/0092-8674(95)90426-3).
- (en) U. Drescher, C. Kremoser, C. Handwerker et al., « In vitro guidance of retinal ganglion cell axons by RAGS, a 25 kDa tectal protein related to ligands for Eph receptor tyrosine kinases », Cell, vol. 82, no 3, , p. 359–370 (PMID 7634326, DOI 10.1016/0092-8674(95)90425-5).
- (en) F. Mann, S. Ray, W. Harris et C. Holt, « Topographic mapping in dorsoventral axis of the Xenopus retinotectal system depends on signaling through ephrin-B ligands », Neuron, vol. 35, no 3, , p. 461–473 (PMID 12165469, DOI 10.1016/S0896-6273(02)00786-9).
- (en) O. Salvucci et G. Tosato, « Essential roles of EphB receptors and EphrinB ligands in endothelial cell function and angiogenesis », Advances in Cancer Research, vol. 114, no 2, , p. 21–57 (PMID 22588055, PMCID 3500853, DOI 10.1016/B978-0-12-386503-8.00002-8).
- (en) J. Bai, Y.J. Wang, L. Liu et Y.L. Zhao, « Ephrin B2 and EphB4 selectively mark arterial and venous vessels in cerebral arteriovenous malformation », The Journal of International Medical Research, vol. 42, no 2, , p. 405–15 (PMID 24517927, DOI 10.1177/0300060513478091).
- (en) O. Salvucci et G. Tosato, « Essential roles of EphB receptors and EphrinB ligands in endothelial cell function and angiogenesis », Advances in Cancer Research, vol. 114, no 2, , p. 21–57 (PMID 22588055, PMCID 3500853, DOI 10.1016/B978-0-12-386503-8.00002-8).
- (en) E.B. Pasquale, « Eph receptors and ephrins in cancer: bidirectional signalling and beyond », Nature Reviews. Cancer, vol. 10, no 3, , p. 165–80 (PMID 20179713, PMCID 2921274, DOI 10.1038/nrc2806).
- (en) Birgit Mosch, Bettina Reissenweber, Christin Neuber et Jens Pietzsch, « Eph Receptors and Ephrin Ligands : Important Players in Angiogenesis and Tumor Angiogenesis », Journal of Oncology, vol. 2010, , p. 1–12 (ISSN 1687-8450, DOI 10.1155/2010/135285).
- (en) Bruce Alberts, Alexander Johnson, Julian lewis, Raff Martin, Keith Roberts et Peter Walter, Molecular Biology of the Cell, Garland Sciences, (ISBN 978-0-8153-4105-5), pages 1440–1441.
- (en) Eduard Batlle, « Wnt signalling and EphB-ephrin interactions in intestinal stem cells and CRC progression », Scientific Report, (lire en ligne [PDF], consulté le ).
- S. Islam, A.M. Loizides, J.J. Fialkovich, R.J. Grand RJ et R.K. Montgomery, « Developmental expression of Eph and ephrin family genes in mammalian small intestine », Digestive Diseases and Sciences, vol. 55, no 9, , p. 2478–88 (PMID 20112066, PMCID 3947671, DOI 10.1007/s10620-009-1102-z).
- (en) Mara Pitulescu, « Eph/ephrin molecules-a hub for signaling and endocytosis », Genes & Development, (DOI 10.1101/gad.1973910, lire en ligne [PDF], consulté le ).
- (en) I. Segura, C.L. Essmann, S. Weinges et A. Acker-Palmer, « Grb4 and GIT1 transduce ephrinB reverse signals modulating spine morphogenesis and synapse formation », Nature Neuroscience, vol. 10, no 3, , p. 301–310 (PMID 17310244, DOI 10.1038/nn1858).
Articles connexes
Liens externes
- (en) « Conserved Protein Domain Family Ephrin_ectodomain », sur le site du National Center for Biotechnology Information (NCBI) (consulté le ).
- (en) « Ephrin ectodomain family », sur le site Superfamily (consulté le ).
- (en) « Family : Ephrin receptor ligand binding domain », sur le site de la base de données Scop (consulté le ).
- (en) « Ephrin receptor-binding (ephrin RBD) domain signature and profile », sur le site de la base de données PROSITE, SIB Swiss Institute Bioinformatics (consulté le ).
- (en) « Ephrin receptor-binding domain (IPR001799) », sur le site de la base de données InterPro (consulté le ).