Flux axoplasmique
Le flux axoplasmique ou transport axonal désigne le transport des macromolécules, et en particulier des protéines, le long de l'axone des neurones, soit dans le sens antérograde, du corps cellulaire vers la synapse, soit dans le sens inverse, dit rétrograde. Ce double flux directionnel a été mis en évidence en 1971 par Liliana Lubińska[1]. Suivant les mécanismes impliqués, ces flux peuvent être rapides (quelques microns par seconde) ou lents (environ cent fois moins vite).
L'essentiel des protéines, et notamment des protéines synaptiques, étant synthétisé au niveau du corps cellulaire, une partie de ces molécules doit donc être acheminée sur toute la longueur de l'axone pour pouvoir être utilisée dans la neurotransmission. Le flux axoplasmique permet par exemple de transporter jusqu'à la terminaison axonale, les vésicules synaptiques dont le contenu en neurotransmetteurs a été synthétisé au sein de l'appareil de Golgi. Le transport axonal permet aussi d'évacuer des molécules des axones pour les porter vers le soma, où elles seront dégradées par les lysosomes[2].
Les mécanismes sous-jacents au transport axoplasmique sont encore imparfaitement connus mais ils s'appuient vraisemblablement sur les microtubules et les protéines motrices, kinésine et dynéine, s'agissant des flux rapides tandis que les flux lents reposent sur la contribution de mécanismes multiples (transport actif, diffusion, renouvellement et croissance cytoarchitectonique).
Ce flux permet la régénérescence de neurones sectionnés, et c'est d'ailleurs cette observation qui a conduit à sa découverte[3].
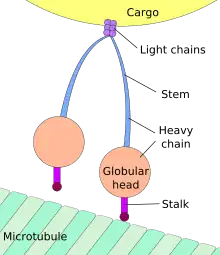
Le transport vers la soma est qualifié de « rétrograde », et celui vers les synapses « transport antérograde[4] - [5]. »
Mécanisme physico-chimique
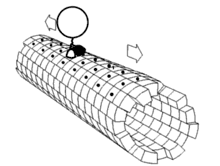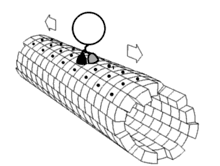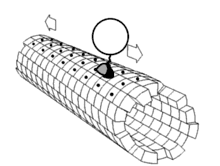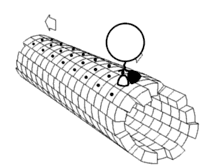
La grande majorité des protéines axonales sont synthétisées dans le soma neuronal puis acheminées le long des axones. On a pu mettre en évidence quelques formes de traduction génétique au sein des axones[6] - [7]. Le transport axonal intervient tout au long de la vie d'un neurone et joue un rôle essentiel dans sa croissance et sa persistance. Les protéines de structure du cytosquelette sont acheminées le long de microtubules (en tubuline) qui ceinturent les axones. Les vecteurs de ces protéines sont la kinésine et la dynéine, protéine motrices qui progressent dans le sens antérograde (depuis le soma vers la pointe de l'axone) et rétrograde (retour au soma), respectivement. Ces protéines motrices assurent aussi bien le transport des mitochondries, des polymères du cytosquelette, des autophagosomes que des vésicule synaptiques, qui contiennent les neurotransmetteurs.
Mode « lent » et mode « rapide »
Les vésicules se déplacent relativement vite (50–400 mm/jour), tandis que le transport des protéines solubles (cytosolique) et de structure est beaucoup plus lent (moins de 8 mm/jour[8]). Si le mécanisme de base du transport axonal rapide est connu depuis des décennies, le mécanisme du transport axonal lent n'a été élucidé que récemment, grâce aux techniques de pointe de l'imagerie médicale[9]. Les techniques de sonde fluorescente (par ex. la microscopie à fluorescence) ont permis de visualiser le transport dans des neurones vivants.
Des études récentes ont montré que la lenteur du mouvement des protéines de structure tien en fait à son caractère saccadé. Ce mécanisme dit de stop and go a été particulièrement étudié[10] pour les polymères de structure du neurofilament. Le mouvement des nutriments solubles (cytosoliques) est plus complexe encore, mais semble de même nature : les protéines solubles se combinent en protéines complexes qui s'associent sporadiquement aux autres protéines cheminant par le canal axonal « rapide[11] - [12] - [13]. »
Le transport antérograde
Le transport antérograde (ou « orthograde ») désigne le flux des molécules ou organites du soma vers les synapses ou la membrane plasmique.
Le mouvement antérograde (lent ou rapide) des vésicules de transport le long des microtubule est assuré par les kinésines[5] - [2]. Le mode lent fait intervenir des kinésines de différents types[9] ; mais le mécanisme qui déclenche les interruptions du transit demeure inconnu.
Il y a deux catégories de transport antérograde lent : la composante lente a (SCa) achemine microtubules et neurofilaments à 0.1-1 mm/jour, et la composante b (SCb) qui transporte plus de 200 protéines diverses et l'actine à la vitesse de 6 mm/jour[9]. Le transport de l'actine dans les axones de la rétine a lieu à 2-3 mm/jour.
Lorsqu'il sort de sa période de latence, l'herpès entame son cycle lytique, et exploite les mécanismes de transport antérograde pour migrer des neurones du ganglion spinal vers la chair ou muqueuse qu'il va contaminer[14].
La protéine précurseur de l'amyloïde (APP), apparentée à celle qui occasionne les plaques séniles de la maladie d'Alzheimer[15], est un récepteur des kinésines, protéines motrices du transport antérograde. Un peptide à 15 acides aminés fixé sur le radical carboxyl cytoplasmique de l'APP fixe la kinésine-1, permet le transport de transmetteurs exogènes dans l'axone géant du calmar[16].
Le traceur au manganèse, un produit de contraste pour la relaxation longitudinale (T1), inventé par Robia Pautler, Elaine Bearer and Russ Jacobs, une fois introduit par injection stéréotactique dans le cerveau d'animaux de laboratoire, migre par transport antérograde et permet de visualiser in vivo le réseau de l'ensemble des neurones.
Transport rétrograde
Le transport rétrograde évacue les molécules/organites des terminaisons des axones vers le soma. Il est assuré par la dynéine cytoplasmique, et sert par exemple à envoyer les signaux chimiques et produits de l'endocytose des endolysosomes de l'axone vers le noyau cellulaire[2]. En moyenne, il développe in vivo une vitesse d'environ 2 μm/sec[17] - [18], fast retrograde transport can cover 10-20 cm/jour[2].
Le transport rétrograde rapide renvoie les vésicules synaptiques usées et d'autres molécules vers le soma. Il transmet les signaux des synapses vers le TRK, récepteur du facteur de croissance du nerf[19]. Certains pathogènes exploitent ce processus pour contaminer le système nerveux : ils pénètrent par les terminaisons distales d'un axone et progressent vers le soma par transport rétrograde. Il en va ainsi pour les toxines du tétanos et le virus de l'herpès, de la rage et de la polio. Dans ces infections, le délai entre l'infection et l'apparition des symptômes correspond au temps mis par les agents pathogènes à atteindre le soma[20]. le virus de l'herpès chemine le long de l'axone dans les deux directions au cours de son cycle de vie, avec une tendance dominante au transport rétrograde pour ses premiers capsides[21].
Conséquences d'une discontinuité
À chaque fois que le transport axonal est gêné ou interrompu, une pathologie se déclenche, formant un amas d'axoplasme appelé sphéroïde axonal. Le transport axonal pouvant être affecté d'une multitude de façons, on retrouve ces sphéroïdes axonaux dans différentes classes de maladies : génétiques, traumatiques, ischémiques, infectieuses, dégénératives ; dans les empoisonnements et les leucoencéphalites. Plusieurs maladie neurodégénératives rares résultent de mutations génétiques des protéines motrices, kinésine et dynéine, de sorte qu'il est probable que le transport axonal joue un rôle essentiel dans le développement de la maladie[22] - [23]. Les dysfonctionnements du transport axonal seraient à l'origine des formes sporadiques (courantes) des troubles neurodégénératifs comme la maladie d'Alzheimer et la maladie de Parkinson[9] : on l'a déduit de la présence systématique de gros globules axonaux dans les neurones contaminés, et du fait que les gènes intervenant dans les formes héréditaires de ces maladies influent sur le transport axonal normal ; on n'en a toutefois aucune preuve directe, et d'autres mécanismes (comme une intoxication des synapses) seraient plus craisemblables.
Dans les rétinopathies vasculaires, c'est l'arrêt du flux axoplasmique à la surface des aires ischémiques qui produit une dilatation des fibres nerveuses entraînant une exsudation ou un foisonnement des fibres.
Dans la mesure où un axone est alimenté en protéines vitales et polymères de construction par le transport axoplasmique, les lésions telle la lésion axonale diffuse vont déclencher un processus de dégénérescence wallérienne chez l'axone distal. La chimiothérapie, en altérant les microtubules (qui sont nécessaires à la division cellulaire) endommage les nerfs en affectant le transport axonal.
Dans les infections
Le virus de la rage contamine le système nerveux central par flux axoplasmique rétrograde[24]. La neurotoxine du tétanos est métabolisée à la jonction neuromusculaire en se combinant à la protéine nidogène et est rétro-induite vers le soma par les endosomes transmetteurs[25]. Les virus neurotropes, comme ceux de l’herpès, cheminent à l'intérieur des axones par transport cellulaire, comme l'a montré l'équipe d'Elaine Bearer[26] - [27]. On suspecte d'autres agents infectieux de progresser par transport axonal[28], et de contribuer à déclencher la maladie d'Alzheimer et d'autre maladies dégénératives[29] - [30].
Voir également
- transport intraflagellaire
Notes et références
- (en) Lubińska L et Niemierko S., « Velocity and intensity of bidirectional migration of acetylcholinesterase in transected nerves. », Brain Res., vol. 27, no 2, , p. 329-42 (PMID 4101562, DOI 10.1016/0006-8993(71)90258-7, lire en ligne, consulté le )
- E. Oztas, « Neuronal Tracing », Neuroanatomy, vol. 2, , p. 2–5 (lire en ligne)
- (en) Lubińska L., « On axoplasmic flow. », Int Rev Neurobiol., vol. 17, , p. 241-96 (PMID 49329, DOI 10.1016/s0074-7742(08)60211-1, lire en ligne, consulté le )
- G. Karp, P. van der Geer, Cell and molecular biology: concepts and experiments, John Wiley (réimpr. 4e) (ISBN 978-0-471-46580-5, lire en ligne), 344
- M.F. Bear, B.W. Connors et M.A. Paradso, Neuroscience : exploring the brain, Lippincott Williams & Wilkins, (réimpr. 3e) (ISBN 978-0-7817-6003-4, lire en ligne), p. 41
- M. Giustetto, A.N. Hegde, K. Si, A. Casadio, K. Inokuchi, W. Pei, E.R. Kandel, J.H. Schwartz, « Axonal transport of eukaryotic translation elongation factor 1alpha mRNA couples transcription in the nucleus to long-term facilitation at the synapse », Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no 23, , p. 13680–5 (PMID 14578450, PMCID 263873, DOI 10.1073/pnas.1835674100, Bibcode 2003PNAS..10013680G)
- K. Si, M. Giustetto, A. Etkin , R. Hsu , et al., « A neuronal isoform of CPEB regulates local protein synthesis and stabilizes synapse-specific long-term facilitation in aplysia », Cell, vol. 115, no 7, , p. 893–904 (PMID 14697206, DOI 10.1016/s0092-8674(03)01021-3, S2CID 15552012)
- (en) Sandra Maday, Alison E. Twelvetrees, Armen J. Moughamian et Erika L.F. Holzbaur, « Axonal Transport: Cargo-Specific Mechanisms of Motility and Regulation », Neuron, vol. 84, no 2, , p. 292–309 (PMID 25374356, PMCID 4269290, DOI 10.1016/j.neuron.2014.10.019)
- S. Roy , B. Zhang , V.M. Lee , J.Q. Trojanowski, « Axonal transport defects: a common theme in neurodegenerative diseases », Acta Neuropathologica, vol. 109, no 1, , p. 5–13 (PMID 15645263, DOI 10.1007/s00401-004-0952-x, S2CID 11635065)
- A. Brown, « Axonal transport of membranous and nonmembranous cargoes: a unified perspective », The Journal of Cell Biology, vol. 160, no 6, , p. 817–21 (PMID 12642609, PMCID 2173776, DOI 10.1083/jcb.200212017)
- Scott DA, Das U, Tang Y, Roy S, « Mechanistic logic underlying the axonal transport of cytosolic proteins », Neuron, vol. 70, no 3, , p. 441–54 (PMID 21555071, PMCID 3096075, DOI 10.1016/j.neuron.2011.03.022)
- S. Roy , M.J. Winton , M.M. Black , J.Q. Trojanowski , V.M. Lee, « Rapid and intermittent cotransport of slow component-b proteins », The Journal of Neuroscience, vol. 27, no 12, , p. 3131–8 (PMID 17376974, PMCID 6672457, DOI 10.1523/JNEUROSCI.4999-06.2007)
- Andreï V. Kouznetsov, « Analytical solution of equations describing slow axonal transport based on the stop-and-go hypothesis », Central European Journal of Physics, vol. 9, no 3, , p. 662–673 (DOI 10.2478/s11534-010-0066-0, Bibcode 2011CEJPh...9..662K)
- D.J. Holland, M. Miranda-Saksena, R.A. Boadle, P. Armati, A.L. Cunningham, « Anterograde transport of herpes simplex virus proteins in axons of peripheral human fetal neurons: an immunoelectron microscopy study », Journal of Virology, vol. 73, no 10, , p. 8503–11 (PMID 10482603, PMCID 112870, DOI 10.1128/JVI.73.10.8503-8511.1999)
- P. Satpute-Krishnan , J.A. DeGiorgis , M.P. Conley , M. Jang , E.L. Bearer, « A peptide zipcode sufficient for anterograde transport within amyloid precursor protein », Proceedings of the National Academy of Sciences of the United States of America, vol. 103, no 44, , p. 16532–7 (PMID 17062754, PMCID 1621108, DOI 10.1073/pnas.0607527103, Bibcode 2006PNAS..10316532S)
- Seamster PE, Loewenberg M, Pascal J, Chauviere A, Gonzales A, Cristini V, Bearer EL, « Quantitative measurements and modeling of cargo-motor interactions during fast transport in the living axon », Physical Biology, vol. 9, no 5, , p. 055005 (PMID 23011729, PMCID 3625656, DOI 10.1088/1478-3975/9/5/055005, Bibcode 2012PhBio...9e5005S)
- K.L. Gibbs, B. Kalmar, J.N. Sleigh, L. Greensmith, G. Schiavo, « In vivo imaging of axonal transport in murine motor and sensory neurons », Journal of Neuroscience Methods, vol. 257, , p. 26–33 (PMID 26424507, PMCID 4666412, DOI 10.1016/j.jneumeth.2015.09.018)
- James Sleigh et Giampietro Schiavo, « Older but not slower: aging does not alter axonal transport dynamics of signalling endosomes in vivo », Matters, vol. 2, no 6, (DOI 10.19185/matters.201605000018)
- B. Cui, C. Wu , L. Chen , A. Ramirez , E.L. Bearer , W.P. Li , W.C. Mobley, S. Chu, « One at a time, live tracking of NGF axonal transport using quantum dots », Proceedings of the National Academy of Sciences of the United States of America, vol. 104, no 34, , p. 13666–71 (PMID 17698956, PMCID 1959439, DOI 10.1073/pnas.0706192104, Bibcode 2007PNAS..10413666C)
- Kenneth Saladin, Anatomy and Physiology: The Unity of Form and Function, New York, McGraw-Hill, (réimpr. 6e), 445 p..
- E.L. Bearer, X.O. Breakefield, D. Schuback, T.S. Reese, J.H. LaVail, « Retrograde axonal transport of herpes simplex virus: evidence for a single mechanism and a role for tegument », Proceedings of the National Academy of Sciences of the United States of America, vol. 97, no 14, , p. 8146–50 (PMID 10884436, PMCID 16684, DOI 10.1073/pnas.97.14.8146, Bibcode 2000PNAS...97.8146B)
- S. Maday, A.E. Twelvetrees, A.J. Moughamian , E.L. Holzbaur, « Axonal transport: cargo-specific mechanisms of motility and regulation », Neuron, vol. 84, no 2, , p. 292–309 (PMID 25374356, PMCID 4269290, DOI 10.1016/j.neuron.2014.10.019)
- Sleigh JN, Rossor AM, Fellows AD, Tosolini AP, Schiavo G, « Axonal transport and neurological disease », Nat Rev Neurol, vol. 15, no 12, , p. 691–703 (PMID 31558780, DOI 10.1038/s41582-019-0257-2, S2CID 203437348, lire en ligne)
- E. Mitrabhakdi, S. Shuangshoti, P. Wannakrairot, R.A. Lewis, K. Susuki, J. Laothamatas, T. Hemachudha, « Difference in neuropathogenetic mechanisms in human furious and paralytic rabies », Journal of the Neurological Sciences, vol. 238, nos 1–2, , p. 3–10 (PMID 16226769, DOI 10.1016/j.jns.2005.05.004, S2CID 25509462)
- K. Bercsenyi, N. Schmieg, J.B. Bryson et al., « Tetanus toxin entry. Nidogens are therapeutic targets for the prevention of tetanus », Science, vol. 346, no 6213, , p. 1118–23 (PMID 25430769, DOI 10.1126/science.1258138, S2CID 206560426, lire en ligne)
- Satpute-Krishnan P, DeGiorgis JA, Bearer EL, « Fast anterograde transport of herpes simplex virus: role for the amyloid precursor protein of alzheimer's disease », Aging Cell, vol. 2, no 6, , p. 305–18 (PMID 14677633, PMCID 3622731, DOI 10.1046/j.1474-9728.2003.00069.x)
- Cheng SB, Ferland P, Webster P, Bearer EL, « Herpes simplex virus dances with amyloid precursor protein while exiting the cell », PLOS ONE, vol. 6, no 3, , e17966 (PMID 21483850, PMCID 3069030, DOI 10.1371/journal.pone.0017966, Bibcode 2011PLoSO...617966C)
- Bearer EL, Satpute-Krishnan P, « The role of the cytoskeleton in the life cycle of viruses and intracellular bacteria: tracks, motors, and polymerization machines », Current Drug Targets. Infectious Disorders, vol. 2, no 3, , p. 247–64 (PMID 12462128, PMCID 3616324, DOI 10.2174/1568005023342407)
- R.F. Itzhaki, R. Lathe ,B.J. Balin et al., « Microbes and Alzheimer's Disease », Journal of Alzheimer's Disease, vol. 51, no 4, , p. 979–84 (PMID 26967229, PMCID 5457904, DOI 10.3233/JAD-160152)
- « No place like asphalt for these hardy microbes », New Scientist, vol. 206, no 2757, , p. 15 (DOI 10.1016/s0262-4079(10)60991-8)