Pelagornithidae
Les Pelagornithidae (appelés aussi « oiseaux à pseudo-dents » et « oiseaux à dents osseuses ») forment une famille préhistorique de grands oiseaux de mer. Leurs restes fossiles ont été retrouvés dans le monde entier, au sein de roches datant de la fin du Paléocène jusqu'à la limite entre le Pliocène et le Pléistocène.

Le trait le plus remarquable de ces oiseaux est la présence de pointes semblables à des dents sur les bords de leur bec. Ces pointes, appelées pseudo-dents, étaient des excroissances du prémaxillaire et de l'os mandibulaires et contenaient, contrairement à de véritables dents, des canaux de Volkmann. Les « petites » espèces de Pelagornithidae étaient de la taille des albatros. Les plus grandes avaient une envergure estimée à 5-7 mètres, ce qui les place parmi les plus grands oiseaux volants ayant jamais vécu. Ils étaient les oiseaux dominants de la plupart des océans pendant la majeure partie du Cénozoïque et les derniers Pelagornithidae connus étaient contemporains de l'Homo habilis.
Description et écologie

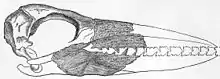
Les plus grands oiseaux volants connus sont les plus grands des Pelagornithidae, le « record » étant détenu par Pelagornis sandersi. La quasi-totalité de leurs restes du Néogène sont immenses[1] mais, au cours du Paléogène, il y a eu un certain nombre de Pelagornithidae, dont le genre Diomedea, qui étaient environ de la taille d'un grand albatros ou même un peu plus petits. L'espèce non décrite désignée sous le nomen nudum « Odontoptila inexpectata », découverte au Maroc et datant de la limite entre le Paléocène et l’Éocène, est le plus petit Pelagornithidae découvert à ce jour. Il était juste un peu plus grand qu'un puffin à menton blanc.
Les Pelagornithidae avaient des os à parois très minces, largement pneumatisés par des sacs aériens, extensions des poumons. De ce fait, la plupart des fossiles osseux sont écrasés. Ces os minces et cette pneumatisation ont permis à ces oiseaux d'atteindre une grande taille tout en restant en-dessous du seuil critique de charge alaire. Bien que 25 kg/m2 soit la charge alaire maximum pour le vol d'un oiseau, il est prouvé que les oiseaux à dents osseuses utilisaient presque exclusivement le vol de gradient : l'extrémité proximale de leur humérus avait une forme diagonale allongée qui rendait difficile le battement d'ailes typique des oiseaux ; leur poids ne peut donc pas être estimé facilement. La zone d'attachement des muscles responsables de l’extension des membres était particulièrement développée et cette anatomie semblait leur permettre de rigidifier leurs ailes au niveau de l'articulation de la cavité glénoïde du coude plus qu'aucun autre oiseau. Cela est particulièrement évident chez les pelagornithides du Néogène, et moins développé pour les formes antérieures du Paléogène. Le sternum avait la forme profonde et courte typique des oiseaux pratiquant le vol de gradient et les attachements osseux étaient solidement ancrés dans la furcula[2] - [3] - [4] - [5] - [6].
Les pattes étaient, proportionnellement, courtes, probablement palmées, et l'hallux vestigiel ou totalement absent. Le tarsométatarse (os de la cheville) ressemblait à celui des albatros tandis que les os de la partie avant étaient similaires à ceux des fulmars. Un second orteil, typique des oiseaux à pseudo-dents, nettement incliné vers l'extérieur, était attaché un peu plus près du genou que les autres. Les pseudo-dents étaient probablement recouvertes par la ramphothèque et il y avait deux sillons qui couraient le long de la partie supérieure du bec, à l'intérieur des crêtes qui portaient les pseudo-dents. Ainsi, lorsque le bec était fermé, seules les « dents » supérieures étaient visibles, les dents inférieures étant cachées par celles-là. À l'intérieur de l’orbite oculaire de quelques-uns, peut-être les plus jeunes espèces, on trouve des glandes à sel bien développées[7] - [3] - [8] - [6] - [9].
Presque aucune partie majeure du corps des Pelagornithidae n’est connue faute d'un fossile bien conservé et le matériel le mieux conservé consiste en un seul os ; il existe cependant par ailleurs, du fait de la longue période durant laquelle ils existèrent et de leur grande taille, quelques restes, écrasés et déformés, d'oiseaux ensevelis après leur mort, accompagnés de plumes fossilisées. De larges parties de crânes et quelques pièces de becs peuvent être trouvées. En , un crâne fossilisé presque complet d'un Odontopteryx (en) présumé, datant de la limite entre le Chasicoien et l'Huayquérien, vers 9 millions d'années BP, fut révélé à Lima. Il avait été trouvé quelques mois plus tôt dans le district d'Ocucaje, dans la province d'Ica, au Pérou. Selon le paléontologue Mario Urbina, découvreur du spécimen, et ses collègues Rodolfo Salas, Ken Campbell et Daniel T. Ksepka, le crâne d'Ocucaje est le mieux préservé des crânes de Pelagornithidae connus[10] - [5].
Écologie et extinction


.jpg.webp)
À l'inverse des oiseaux à « vraies dents » du Mésozoïque, tels Archaeopteryx ou Yanornis, les pseudo-dents des Pelagornithidae ne semblent pas avoir eu des arêtes coupantes dentelées ou spécialisées, et étaient utilisées pour saisir les proies et les déglutir plutôt que pour les déchiqueter. Étant donné que ces pseudo-dents étaient creuses ou, au mieux, remplies de tissus osseux et qu'elles sont souvent usées ou cassées sur les fossiles, on pense qu'elles n'étaient pas extrêmement résistantes même lorsque l'animal était en vie. Les proies de Pelagornithidae étaient donc probablement molles, et étaient composées de cephalopodes[note 1] et de poissons à corps mou[note 2]. Leurs proies étaient elles-mêmes susceptibles d'atteindre des tailles considérables. Bien que certaines reconstitutions montrent les Pelagornithidae comme des oiseaux plongeurs tels les Fous de Bassan et apparentés (Morus), leur os minces et pneumatisés, facilement fracturés comme le montrent les fossiles, font que ce mode d'alimentation est irréaliste et aurait été très dangereux. Il est probable que les proies étaient ramassées près de la surface, tandis que l'oiseau volait ou nageait et que seul le bec était immergé dans la plupart des cas. Leur articulation osseuse de la mâchoire inférieure ressemble à celle des Pélicans qui peuvent ouvrir largement le bec. Finalement, les oiseaux à pseudo-dents pourraient avoir rempli une niche écologique presque identique à celle des plus gros ptérosaures, qui se nourrissaient de poissons, et dont l'extinction à la fin du Crétacé pourrait bien avoir ouvert la voie au règne de 50 millions d'années des Pelagornithidae. À l'instar des albatros modernes, mais à la différence des ptérosaures, les oiseaux à pseudo-dents pourraient avoir utilisé les courants océaniques et la circulation atmosphérique pour établir des circuits circulaires au-dessus des océans ouverts, ne revenant se reproduire que toutes les quelques années seulement. Contrairement aux albatros d'aujourd'hui, qui évitent les courants équatoriaux tropicaux et leurs zones de calme, les Pelagornithidae ont été trouvés dans des zones présentant toutes sortes de climats, et leur présence, qui s'étend sur 40 millions d'années, est attestée de la Belgique et, à travers le Togo, jusqu'à l'Antarctique. Il est démontré que les manchots et les Plotopteridae — animaux plongeurs munis d'ailes qui colonisèrent le plateau continental — se retrouvent presque invariablement en compagnie d'oiseaux à pseudo-dents. Ainsi ces derniers semblent s'être rassemblés en masse dans des régions d'élevage pour se nourrir et se reproduire[3] - [12] - [13] - [14] - [15] - [16] - [17] - [18] - [19] - [5] - [20].


On prétend parfois que, à l'instar d'autres oiseaux de mer (par exemple les oiseaux inaptes au vol tels que les Plotopteridae), l'évolution des cétacés et des pinnipèdes les conduisirent à supplanter les oiseaux à pseudo-dents, entraînant ainsi leur disparition. Bien que cette explication puisse convenir pour les Plotopteridae, elle est moins convaincante concernant les Pelagornithidae, pour deux raisons. La première est que Pelargonis continua à prospérer 10 millions d'années après l'apparition des types modernes de baleines et, au Miocène moyen, il coexista avec Aglaocetus et Eobalaenoptera dans l'Atlantique, au large de ce qui est aujourd'hui la côte Est des États-Unis, tandis que, dans le Pacifique, Osteodontornis (en) vivait dans les mêmes mers que Balaenula et Morenocetus. L'ancêtre du grand cachalot, Aulophyseter et/ou Orycterocetus, apparut dans les deux océans de l'hémisphère nord à la même période, tandis que Brygmophyseter shigensis, un cachalot de taille moyenne, parcourait le Pacifique Nord. En ce qui concerne les pinnipèdes du Miocène, divers morses anciens[note 3] et les ancêtres des otaries, tels Thalassoleon, peuplaient le nord-est, tandis que le léopard de mer Acrophoca était une espèce remarquable du sud-est du Pacifique. En second lieu, les pinnipèdes sont limités aux eaux proches des côtes, tandis que les oiseaux à pseudo-dents parcourent les mers loin au large et, comme les grands cétacés et les grands carnivores, ils s'inscrivent dans la stratégie K, avec des densités de population modérées à très faibles[21] - [22] - [11].
Conséquemment, la compétition pour la nourriture entre les oiseaux à pseudo-dents, les cétacés et les pinnipèdes ne pouvait être très sévère. Cependant, les oiseaux et les pinnipèdes ont besoin de terrains plats proches de l'eau pour élever leur progéniture, et la concurrence pour les aires de reproduction pourrait avoir affecté la population aviaire. À cet égard, la spécialisation des oiseaux pratiquant le vol de gradient limitait le nombre de sites propres à la nidification, mais, d'un autre côté, les hautes-terres costales auraient pu fournir des zones de reproduction inaccessibles aux pinnipèdes, comme cela se passe aujourd'hui pour les albatros, aux Galapagos ou à Torishima. Les oiseaux à dents osseuses ont besoin de forts courants ascendants pour décoller, ce qui suppose des sites en altitude, limitant la compétition avec les colonies de pinnipèdes. En ce qui concerne les lieux de reproduction, des fragments de coquilles géantes provenant des montagnes de Famara à Lanzarote, dans les îles Canaries, ont été attribués provisoirement à ces oiseaux du Miocène supérieur. L'argile de Londres, datant de l'Yprésien, dans l'île de Sheppey, où les fossiles ne sont pas rares, s'est déposée dans une mer épicontinentale à une époque très chaude avec des niveaux marins élevés. Les sites de reproduction présumés ne pouvaient pas être aussi loin au large que certaines colonies le sont aujourd'hui, alors que la région était entourée par les Alpes, les Monts Grampians et les Alpes scandinaves, dans une mer moins large que l'actuelle mer des Caraïbes. Les oiseaux à pseudo-dents du Néogène sont courants le long des côtes américaines près des Appalaches et des cordillères et ils étaient élevés probablement non loin en mer ou même sur les montagnes elles-mêmes. La présence d'os médullaires dans la mine Lee Creek en Caroline du Nord, est remarquable car, chez les oiseaux, on ne trouve en général cela que chez les femelles qui couvent, ce qui montre que les lieux de reproduction n'étaient probablement pas éloignés. Les îles d'origine volcanique du Pacifique ont été érodées au cours des millions d'années écoulées, effaçant les restes des colonies d'élevage de Pelagornithidae qui auraient pu exister en plein air. L'île Necker, par exemple, était d'une taille significative il y a 10 millions d'années, lorsqu'Osteodontornis parcourait le Pacifique[23] - [24] - [25] - [26] - [27] - [17] - [18] - [28].
Il n'y a pas d'explication simple et unique quant à l'extinction des oiseaux à pseudo-dents. Un scénario de changement écologique global — exacerbé par le début d'un âge de glace et des changements dans les courants océaniques dus à des surrections liées aux mouvements des plaques tectoniques (entraînant l'émergence du courant circumpolaire antarctique et la fermeture de l'isthme de Panama) — est le plus probable ; les oiseaux à pseudo-dents, survivants de la faune du Paléogène, n'auraient pas pu s'y adapter. À cet égard, il peut être significatif de remarquer que certains genres de cétacés, comme les dauphins primitifs Kentriodontidae ou les baleines à dents de requin, qui se sont développés en même temps que les Pelagornithidae, se sont éteints à peu près au même moment. En outre, la diversité moderne des genres de pinnipèdes et de cétacés apparaît autour du Mio-Pliocène, ce qui suggère que de nombreuses niches écologiques sont apparues ou sont devenues vacantes. En outre, ce qui a causé la perturbation du Miocène moyen et la crise de salinité messinienne a également eu une incidence non négligeable sur le réseau trophique des océans de la Terre, ce dernier événement entraînant une extinction généralisée des oiseaux de mer. Cette combinaison de facteurs fit que les animaux du Néogène remplacèrent finalement les derniers survivants de la faune du Paléogène durant le Pliocène. Les oiseaux à pseudo-dents les plus anciens se trouvent le plus souvent dans les mêmes dépôts que les spécimens de Plotopteridae et de Sphenisciformes (manchots), tandis que les formes récentes sont associées aux pingouins, albatros et Procellariidae, lesquels connurent une radiation évolutive importante coïncidant avec la disparition du réseau trophique du Paléogène, qui en est probablement la cause. Quoique les fossiles soient nécessairement incomplets, il semble que les cormorans et les mouettes aient été très rarement associés aux Pelagornithidae[15] - [16] - [25] - [29] - [19] - [22] - [30].
Indépendamment de la cause de leur extinction ultime, les oiseaux à pseudo-dents furent, durant leur longue existence, eux-mêmes des proies pour de grands prédateurs. Il y avait très peu de grands oiseaux susceptibles de les attaquer en vol ; on connait la Limnofregata, datant de l'Éocène inférieur, co-existante avec les Pelagornithidae, qui aurait pu attaquer à l'occasion les petites espèces, sachant que les Fregatidae (« frégates ») actuelles pratiquent de même à l’encontre des albatros. Datant de l'Éocène moyen à l'Éocène supérieur, on trouve, dans la mine de Lee Creek quelques restes d'oiseaux à pseudo-dents probablement victimes de requins. L'abondante faune de requins de Lee Creek chassait près de la surface et elle comprenait le requin plat-nez (Notorynchus cepedianus), le requin-taureau (Carcharias taurus), des représentants du genre Isurus et Cosmopolitodus, ainsi que des Carcharodons (requins blancs)[note 4], des requins-tigre (Galeocerdo), des représentants de l'espèce Hemipristis elongata, des requins-citron (Negaprion brevirostris), des requins-marteau (Sphyrnidae). En fonction de l’âge des fossiles des oiseaux, ils auraient pu aussi co-exister avec des poissons-scie (Pristis), des Odontaspis, des Lamniformes et des Parotodus. Il convient de remarquer que les fossiles d'oiseaux plongeurs les plus petits, Alcidae (mergules, guillemots, pingouins, stariques, macareux), plongeons et cormorans, ainsi que les albatros se retrouvent beaucoup plus fréquemment dans les pelotes stomacales de requins que les oiseaux à pseudo-dents, ce qui conforte l'hypothèse qu'ils avaient une faible densité de populations et qu'ils se nourrissaient en vol[25] - [31].
Apparence externe




Rien n'est sûr concernant la couleur de ces oiseaux, car ils n'ont pas de descendance vivante. On peut cependant faire des inférences basées sur leur arbre phylogénétique.
S'ils font partie du groupe des « oiseaux aquatiques supérieurs », ils devaient probablement avoir un plumage similaire à celui de la reconstitution d'Osteodontornis orri présentée ci-contre. Les Procellariiformes et Pelecaniformes moderne (ou Ciconiiformes, si on admet que les Pelecaniformes en font partie), n'ont pratiquement pas de caroténoïde ou de couleur structurelle dans leurs plumes et ils manquent généralement de phéomélanine. Ainsi, les couleurs les plus communes chez ces oiseaux sont le noir, le blanc et les nuances de gris. Certains ont des taches iridescentes ou des teintes rougeâtres ou brunâtres mais cela est limité et les espèces chez qui on les trouve (butors, ibis, ombrettes…) sont en général des espèces d'eau douce[24].
S'ils ressemblent aux Galloanserae, la phéomélanine est beaucoup plus fréquente mais les lignées les plus basales des Anseriformes ont typiquement une couleur noire et grise ou noire et blanche. Parmi les oiseaux océaniques en général, la partie supérieure tend à être plus sombre que la partie inférieure (y compris pour les ailes). Quoique certains pétrels soient uniformément gris foncé, une combinaison de plusieurs gris pour la partie supérieure et de blanc pour la partie inférieure et la tête est la plus commune chez les oiseaux marins et relève peut-être de la symplésiomorphie ou, plus probablement, est un caractère adaptatif procurant un camouflage, en particulier pour leur éviter de se profiler sur le ciel lorsque leurs proies les observent depuis la mer. On peut noter qu'au moins la première rémige, mais aussi, souvent, les pennes, sont noires, même si le reste du plumage est complètement blanc, comme chez les pélicans ou chez l'Étourneau de Rothschild (Leucopsar rothschildi). Cela est dû au fait que la mélanine se polymérise, rendant les plumes noires extrêmement robustes, ce qui leur donne une meilleure résistance aux efforts mécaniques encaissés lors du vol. Chez les oiseaux pratiquant le vol dynamique (vol de gradient) dans des vents forts, tels les oiseaux à pseudo-dents, on peut s'attendre à trouver des extrémités alaires et, peut-être, des queues, noires[note 5].
Concernant les parties dénudées, tous les parents proches des Pelagornithidae ont, le plus souvent, des teintes rougeâtres plutôt claires, en particulier sur le bec. Les incertitudes phylogénétiques qui entourent les Pelagornithidae ne permettent pas de savoir s'ils avaient un sac gulaire similaire à celui des pélicans. S'ils en avaient un, il était probablement rouge ou orange, et pourrait avoir été utilisé pour les parades nuptiales. Le dimorphisme sexuel était probablement presque absent, comme typiquement chez les Ansériformes basaux et les oiseaux aquatiques supérieurs[24].
Taxonomie, systématique et évolution
Les noms « pseudodontornes » ou « pseudodontornithidés » renvoient au genre Pseudodontornis (en), qui servit pendant un temps comme nom de la famille. Le nom de « Pelagornithidae » actuellement utilisé est antérieur à celui de « Pseudodontornithidae » et les auteurs modernes préfèrent par conséquent « pélagornithidés » plutôt que « pseudodontornithidés » ou « pseudodontornes » . Ce dernier nom peut cependant être trouvé dans la littérature du milieu du xxe siècle[32] - [33] - [34].
Historiquement, les os disparates des oiseaux à pseudo-dents furent répartis en six groupes : un certain nombre de genres, décrit à partir des os de pattes furent placés dans une famille nommée Cyphornithidae, considérée comme proche de la famille des pélicans (Pelicanidae). Ils ont ensuite été placés dans une super-famille des « Pelacanides », dans le sous-ordre Pelacanae, puis, plus tard (après que le rang taxonomique ait été fixé à la norme actuelle), parmi les Pelecanoidea dans le sous-ordre Pelecani. Par la suite, certains ont été rangés dans la pseudo-famille Cladornithidae, dans un sous-ordre des Pelecaniformes. Les genres connus grâce aux crânes furent assignés à une ou deux familles (Odontopterygidae) et, parfois, aussi, aux Pseudodontornithidae dans un sous-ordre des Pelecaniformes, Odontopteryges ou Odontopterygia. Pendant ce temps, Pelagornis, décrit à partir des os des ailes, était traditionnellement placé dans la famille monotypique des Pelecaniformes nommée Pelagornithidae. Le sternum de Gigantornis (en) le fit placer dans la famille des albatros, Diomedeidae, dans l'ordre des « oiseaux marins à narines tubulaires », nommée Procellariiformes[35] - [36] - [37] - [3] - [23] - [33] - [1].
La plus grande confusion taxonomique et systématique affecta Dasornis (en). Le genre fut établi sur la base d'une importante partie d'un crâne, et fut longtemps placé parmi les Gastornis simplement à cause de sa taille. Argillornis (aujourd'hui reconnu comme appartenant au genre Dasornis), fut décrit à partir d'os des ailes, et fut généralement inclus parmi les Sulae, en tant que membre d'Elopterygidae, une autre famille invalide, et n’est pas considéré comme un oiseau de type moderne par les auteurs actuels. Des os de tarsométatarse furent placés dans le genre Neptuniavis et assignés aux Procellariidae. En 2008, tous ces fossiles ont désormais montré qu'ils appartiennent au genre d'oiseaux à pseudo-dents Dasornis[35] - [38] - [39] - [40] - [41] - [5] - [1].
Systématique et phylogénie
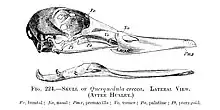

On voit aussi l’empreinte supra-orbitale de la glande à sel.
La systématique des oiseaux à pseudo-dents est sujette à d'âpres débats. À l'origine, ils furent considérés comme apparentés avec les (désormais paraphylétiques) Pelecaniformes (les pélicans et d'autres tels que Morus et Fregatidae) et les Procellariiformes (oiseaux marins à narines tubulaire tels que les albatros et les pétrels), à cause de la similarité globale de leur anatomie. Quelques-uns des premiers fossiles du grand Dasornis furent pris pour ceux d'un ratite puis d'un diatryma. Ils furent invoqués comme argument concernant la similarité entre ces deux groupes ; de fait, les pélicans et les oiseaux à narines tubulaires, comme, par exemple, les autres Pelecaniformes (cormorans et apparentés), qui sont aujourd'hui séparés entre Phalacrocoraciformes, Ciconiiformes (cigognes, héron, ibis et Pelecaniformes typiques) et Gaviiformes (plongeons et autres), semblent être issus d'une radiation évolutive, et forment possiblement un clade d'« oiseaux marins supérieurs ». Les Pelagornithidae ne sont cependant pas considérés comme un chaînon manquant entre pélicans et albatros, mais sont considérés comme nettement plus proches des premiers et, dans une approche écomorphologique, seulement convergents avec les derniers[42] - [43] - [44] - [45] - [46] - [1].
En 2005, une analyse cladistique suggère une étroite relation entre les oiseaux à pseudo-dents et les Anseriformes. Ces derniers ne font pas partie des « oiseaux aquatiques supérieurs » mais des Galloanserae, une lignée basale d'oiseaux neognathes. Certaines caractéristiques, principalement du crâne, soutiennent cette hypothèse. Par exemple, les pelagornithides n'ont pas de crête sur le bas de leur os palatin, alors que les Neoaves (le « clade-frère » des Galloanserae, qui inclut les « oiseaux marins supérieurs » et les « oiseaux terrestres supérieurs ») ont un telle crête. De même, comme les canards, les oies et les cygnes, les pelagornithides ont seulement deux et pas trois condyles sur le processus de l'os carré mandibulaire, avec le condyle moyen au-dessus du condyle latéral. Leur articulation basiptérygoïde est similaire à celle des Galloanseres. Ils présentent une large plate-forme sur le côté du parasphénoïde, comme les Anseriformes. Chez les oiseaux à pseudo-dents, le point d'attache de la partie coronoïdale du muscle adducteur externe de la machoire est située au milieu, le processus rostroptégyroïde possède un support à sa base et l'os mésethmoïde présente une profonde dépression pour la conque caudale, à l'instar des Anseriformes[45] - [5] - [1].
En ce qui concerne les autres parties du squelette, la synapomorphie des Pelagornitidae et des Galloanseres se retrouve essentiellement dans les bras et les os des « mains » ; l'ulna présente une partie supérieure fortement convexe à son extrémité côté coude — à l'extrémité de laquelle sont attachés les muscles scapulotricipitaux —, un cotyle dorsal en pointe et une légère dépression pour abriter le ménisque entre l'ulna et le radius ; vers le coude, le sillon intercondylien de l'ulna devient large et il est bordé par une longue crête sinueuse du côté ventral. Le radius, quant à lui, possède un bord ventral convexe sur le cotyle huméral, qui se continue sur le bord inférieur de la proéminence à laquelle s'attache le muscle biceps brachial ; vers la partie supérieure du radius, la surface devient plate et triangulaire à l'approche de la surface articulaire de l'ulna. Le carpométacarpe des oiseaux à pseudo-dents et celui des Anseriformes présente une apophyse pisiforme développée qui s'étend loin des doigts depuis la trochlée carpale le long de la face avant de l'os. Sur la face inférieure du carpométacarpe, il existe une longue et fine symphyse du métacarpe distal, et le gros os métacarpien présente une crête intermédiaire qui, à son extrémité extérieure, se courbe vers le haut, et l'articulation du pouce a un bouton bien développé sur le côté postérieur de sa surface articulaire. Les os de la patte et du pied, comme on s'y attend concernant des oiseaux qui ne sont pas spécialisés dans la nage, à l'inverse des Anseriformes, montrent moins de similarités avec ces derniers. Le tibiotarse montre une grande incision entre les condyles, et le condyle central est plus étroit que le condyle latéral et fait saillie vers l’avant ; le tarsométatarse possède un foramen vasculaire distal avec une ouverture sur la surface plantaire et une trochlée au milieu des orteils, allongée, légèrement oblique, dirigée vers le dessous du pied et pointue à son extrémité[45].
On ne sait pas vraiment quoi déduire des apomorphies supposées relier Anseriformes et oiseaux à pseudo-dents car, d'un autre côté, le sternum, l'humérus distal, les os des jambes et des pieds des Pelagornithidae semblent présenter des apomorphies avec les « oiseaux marins supérieurs ». Alors que le détail des os du neurocrâne est considéré comme très informatif concernant la phylogénétique, les caractéristiques des crânes dans chacun des deux groupes sont très similaires quant à la façon dont le bec s’attache au crâne, suggérant une pression de sélection similaire sur la fonction consistant a prélever des proies près de la surface maritime. Les traits apparemment non-néoaviens des Pelagornithidae pourraient n'être que des plésiomorphies ataviques ; dans la mesure où les « oiseaux marins supérieurs » sont des Neoaves très anciens et qu'aucun des membres basaux supposés de leur radiation évolutive (cf. Graculavidae) n'a été inclus dans l'analyse, on ne sait pas de manière certaine quand les conditions dérivées typiques des Neoaves ont été acquises. Chez les oiseaux, les caractéristiques des os du pied sont fortement dépendantes des conditions de sélection, avec des effets de convergence évolutive qui inhibent ou invalident les analyses cladistiques ; cependant, les autapomorphies apparentes de l’avant-bras et des os des « mains » sont difficiles à expliquer par autre chose qu'une relation effective. L'emplacement des glandes à sel à l'intérieur de l'orbite oculaire chez Osteodontornis, Pelagornis, et probablement d'autres, montre que, quel que soit leur apparentement, les Pelagornithidae s'adaptent à un environnement océanique de manière indépendante de celle des Sphenisciformes et des oiseaux à narines tubulaires, qui ont des glandes à sel supra-orbitales. Leur hallux vestigiel ou absent — à l'instar des canards mais à la différence des pélicans, lesquels ont leur quatre orteils développés et palmés — va à l'encontre d'un apparentement étroit avec les pélicans. On sait désormais que les pélicans sont plus proches des cigognes (qui possèdent un hallux mais ne sont pas palmées) que des oiseaux à pseudo-dents et ont évolué vers des pattes palmées indépendamment. Présentant simultanément des pattes palmées et un hallux hypotrophié apomorphique et paraphylétique, l'absence d'halux chez les oiseaux à pseudo-dents ne donne que peu d'informations sur leur apparentement[47] - [3] - [48] - [49] - [45] - [50] - [5] - [1] - [9] - [51].
Alors que les Galloanserae géants étaient communs et diversifiés, particulièrement au Paléogène, Gastornis et Dromornithidae étaient des oiseaux terrestres inaptes au vol ; il est sans doute significatif que les seuls autres oiseaux à dents osseuses connus jusqu'à présent soient deux espèces disparues de moa-nalo (en) des îles Hawaï, du genre Thambetochen (en), des canards (Anatinae) géants inaptes au vol. En tout cas, l'analyse cladistique de 2005 utilise un échantillon représentatif de Procellariiformes et les intègre dans un clade en accord avec le consensus actuel. Les relations censément étroites entre les oiseaux à dents osseuses et ceux à narines tubulaires peuvent être abandonnées. Pour ce qui concerne les Pelecaniformes, l'analyse ne permet pas d'établir une phylogénie correcte et n'inclut pas le Bec-en-sabot du Nil (Balaeniceps rex, un chaînon manquant entre pélicans et cigognes) ; à l'évidence, la radiation évolutive de la lignée pélican-cigogne est infirmée par l'analyse. En outre, les Galloanserae ne sont pas monophylétiques. En 2007, une analyse cladistique plus complète de l'anatomie des oiseaux, incluant quelques formes fossiles (mais pas les taxons cruciaux[note 6] du Crétacé supérieur, connus en général à partir de restes fragmentaires), résout le problème de l'évolution radiative des « oiseaux marins supérieurs » de meilleure manière ; la problématique des caractéristiques des membres inférieurs résiste encore cependant à l'analyse[45] - [53] - [1].
Dans la mesure où les relations entre Galloanserae et « oiseaux marins supérieurs » restent irrésolues, les oiseaux à pseudo-dents sont ici placés par compromis dans un ordre distinct, Odontopterygiformes plutôt que dans un sous-ordre pelecaniforme/ciconiiforme ou anseriforme Odontopterygia ou même une famille d'Anseriformes, de Ciconiiformes ou de Pelecaniformes. Une telle solution a peu de chances d'être totalement erronée quel que soit le cas de figure, car les oiseaux à pseudo-dents sont bien distincts des Presbyornithidae et Scopidae, aujourd'hui considérés comme des divergences basales de, respectivement, les Anseriformes et le groupe pélican-cigogne. Cela prévoit aussi une marge de manœuvre si la séparation des Pelagornithidae en plusieurs familles s'avère appropriée. Lorsque Boris Spulski établit l'ordre Odontopterygia en 1910, il le fait en partie parce qu'il remarque quelques similarités entre les oiseaux à pseudo-dents et les Anseriformes listés ci-dessous. Dasornis fut longtemps pris pour un diatryma (Gastornithiformes (en)), aujourd'hui supposé être très proche des Anseriformes. Le pelagornithide Paleochenoides mioceanus (en) fut lui aussi considéré comme un Anseriforme et ce fut la même chose pour le cygne de l'Oligocène, Guguschia nailiae. Dans ce dernier cas, cependant, une analyse « beaucoup plus convaincante »[54] le plaçant hors des Galloanseres fut publiée dès l'année suivante. La plupart des ossements de pelagornithidiques non reconnus ont d'abord été assignés à des familles d'oiseaux marins supérieurs, généralement aux (désormais paraphylétiques) Pelecaniformes, mais le tarsométatarse en particulier fut généralement confondu avec celui d'un procellariiforme. Les Odontopterygiformes furent d'abord proposés lorsqu'Osteodontornis fut décrit comme le premier, et toujours le seul, squelette raisonnablement complet d'un de ces oiseaux. Hildegarde Howard a constaté que, quoique la combinaison de ses caractéristiques ressemblât à celle d'autres oiseaux, elle était différente de celles de tous les néognathes connus[55] - [3] - [43] - [47] - [45] - [5] - [56] - [9] - [51].
Genres et spécimens non identifiés
Du fait de l'état fragmentaire et de l'écrasement de la plupart des restes des oiseaux à pseudo-dents, il n'est pas certain, pour une douzaine de genres, qu'ils soient valides. Seuls les becs sont suffisamment robustes et distinctifs pour permettre une délimitation taxonomique correcte et même ces pièces sont le plus souvent brisées. Par exemple, Argilliornis et Neptuniavis furent récemment considérés comme étant des os des ailes et des pattes de Dasornis qui était, jusque-là, connu seulement à partir d'os crâniens. La taille est généralement considérée comme un marqueur fiable de la diversité du genre, mais il reste à déterminer si les petits spécimens ne concernent pas de jeunes oiseaux[57] - [5] - [58].
De manière provisoire, les genres suivants sont reconnus[23] - [59] - [58] :
- Pseudodontornis (Paléocène supérieur ?–? Oligocène supérieur de Charleston, Caroline du Nord, É.-U.) – polyphylétique (espèce-type de Palaeochenoides/Pelagornis) ?
- Odontoptila (Paléocène supérieur-Éocène inférieur d'Ouled Abdoun, Maroc) – un nomen nudum - Odontopteryx n. sp. 1" d'après Bourdon (2005). Ensemble d'os crâniens et corporels provenant des collections de l'OCP et de la Rhinopolis Association. Ce sont les plus petits oiseaux à pseudo-dents connus à la date de mi-2009[45] - [60] - [61].
- Odontopteryx (Paléocène supérieur-Éocène inférieur de Ouled Abdoun, Maroc – Éocène moyen d'Ouzbékistan) – comprend Neptuniavisminor et, peut-être, Pseudodontornis longidentata, P. tschulensis et Macrodontopteryx.
- Dasornis (Éocène inférieur - Argile de Londres de l'île de Sheppey, Angleterre) – comprend Argillornis, Lithornis emuinus et Neptuniavis miranda ; comprend peut-être Odontopteryx gigas (un nomen nudum), Pseudodontornis longidentata et Gigantornis.
- Macrodontopteryx (Éocène inférieur - Argile de Londres de l'île de Sheppey, Angleterre) – inclut peut-être Pseudodontornis longidentata et/ou appartient à Odontopteryx.
- cf. Odontopteryx (Éocène inférieur de Virginie, É.-U.)[49] - [33] - [62].
- Gigantornis (Éocène moyen d'Ameki, Nigeria) – appartient peut-être à Dasornis.
- cf. Odontopteryx (Éocène moyen du Mexique) - Spécimen MHN-UABCS Te5/6–517. Extrémité distale de l'humérus d'un petit oiseau à pseudo-dents, environ de la taille d'un pélican brun (Pelecanus occidentalis), trouvé dans la formation de Tepetate, près d'El Cien, Basse-Californie du Sud, Mexique[49].
- Pelagornithidae gen. et esp. indét. (Éocène moyen de Mount Discovery, Antarctique) – idem grand spéciment de Seymour Island Dasornis/Gigantornis ? - Une partie d'humérus d'une espèce de grande taille[63].
- Pelagornithidae gen. et esp. indét. (Éocène moyen d'Etterbeek, Belgique) – Dasornis/Macrodontopteryx ? - une espèce de taille moyenne, auparavant au moins partiellement rattachée à Argillornis[38] - [61] - [64] - [30].
- Aequornis (Éocène moyen de Kpogamé-Hahotoé, Togo) – un nomen nudum d'une espèce de grande taille[60] - [61].
- Pelagornithidae gen. et esp. indét. (Éocène supérieur de La Meseta, île de Seymour, Antarctique) – deux espèces ? similaires au spéciment de Mount Discovery, Dasornis/Gigantornis, Odontopteryx ? Une pièce large, une petite pièce de la machoire supérieure et une petite partie de la machoire inférieure[65] - [66] - [67] - [63] - [6].
- Pelagornithidae gen. et esp. indét. (Éocène supérieur, France) - Mammal Paleogene zone (en) MP19 (Priabonien). Pas plus de détails[33].
- Pelagornithidae gen. et esp. indét. (Éocène supérieur du Kazakhstan) – appartient peut-être à Zheroia. Pièce de bec[61].
- Pelagornithidae gen. et esp. indét. (Éocène des îles Shetland du Sud, Atlantique Sud)[33].
- cf. Dasornis, comme Argillornis[68] (Éocène supérieur-Oligocène inférieur de l'Orégon, É.-U.) – Cyphornis ? Spécimen LACM 128462, extrémité proximale d'un ulna gauche, provenant de la « formation de Keasey », Orégon, É.-U. ; vraisemblablement aussi LACM 127875, fragments de l'extrémité proximale d'un humérus, partie proximale droite d'un ulna et radius d'un même individu provenant de la « formation de Pittsburg Bluff », Orégon, É.-U. Une très grande espèce, sans doute le plus grand oiseau à pseudo-dents connu. Warheit (2001) liste deux espèces et leur donne un âge datant de l'Éocène moyen, mais ceci est erroné[68] - [49] - [62].
- cf. Macrodontopteryx (Oligocène inférieur, Hamstead, Angleterre) – appartient peut-être à Proceriavis. Radius distal d'une espèce de grande taille. Les restes de Proceriavis sont une vertèbre cervicale (spécimen du British Museum of Natural History réf. A-4413) et, peut-être, une phalange[69] - [70] - [71]. E. helveticus est un lapsus.
- Pelagornithidae gen. et esp. indét. (Oligocène inférieur du Japon). Une pointe de bec, provenant de la « formation d'Iwaki », et du matériel additionnel du « groupe de Kishima », Kyūshū[13] - [72] - [6].
- Caspiodontornis (Oligocène supérieur de Pirəkəşkül, Azerbaïdjan) – appartient peut-être à Guguschia.
- Palaeochenoides (Oligocène supérieur de Caroline du Sud, É.-U.) – inclut peut-être Pseudodontornis longirostris ou à Pelagornis.
- Pelagornithidae gen. et esp. indét. (Oligocène supérieur de Caroline du Sud, É.-U.). Une espèce de grande taille, comparable en taille à Osteodontornis et Pelagornis[16].
- Pelagornithidae gen. et esp. indét. (Oligocène supérieur de Yamaga, Kitakyushu, Japon) – Osteodontornis ?. Humérus distal gauche et quelques os de l'aile. Le Miocène inférieur avancé par Warheit (2001) est probablement une erreur[72] - [49] - [6].
- Tympanonesiotes (Oligocène supérieur ou Miocène inférieur de Cooper River, É.-U.).
- Cyphornis (Miocène inférieur de Carmanah Point, île de Vancouver, Canada) – inclut peut-être Osteodontornis.
- Osteodontornis (Miocène inférieur - Pliocène inférieur) – appartient peut-être à Cyphornis.
- Pelagornis (Miocène inférieur de l'Armagnac, France – Péistocène inférieur de Ahl al Oughlam, Maroc) – inclut peut-être Pseudodontornis longirostris et Palaeochenoides.
- Pelagornithidae gen. et esp. indét. (Miocène inférieur ? – Pliocène inférieur de l'est des É.-U.) – deux ou trois espèces ? Pelagornis ? « Pelagornithidae sp. A » et « Pelagornithidae sp. B » chez Warheit (2001) ; « Pelagornis sp. 1 » et « Pelagornis sp. 2 » chez Olson & Rasmussen (2001). Principalement des pièces de fémur et d'humérus d'un oiseau un peu plus petit qu'Osteodontornis et d'autres légèrement plus grands que Tympanonesiotes ainsi que quelques autres os. Le spécimen 476044 de l'USNM, un tarsométatarse, est plus petit et relève peut-être d'une troisième espèce[32] - [73] - [25].
- cf. Osteodontornis (Miocène moyen de Capadare, Cueva del Zumbador, Venezuela). Spécimen MBLUZ-P-5093, une très grande prémaxillaire[17].
- cf. Osteodontornis/Pelagornis (? Miocène moyen/supérieur de North Canterbury, Nouvelle-Zélande). Spécimen CMNZ AV 24,960, une pièce proximale d'humérus (initialement identifiée comme distale) d'une espèce de grande taille[74] - [75] - [76].
- cf. Pelagornis (Miocène moyen de Bahía Inglesa, Chili – Pliocène inférieur du Chili et du Pérou) – deux espèces ? Museo Paleontológico de Caldera 1001 to 1006 (divers morceaux de crânes et de becs, extrémité proximale d'un ulna gauche et deux vertèbres cervicales du Miocène moyen de la « formation de Bahia Inglesa » ; auparavant assigné à Pseudodontornis longirostris par erreur. UOP/01/81 (première phalange d'un second doigt gauche), UOP/01/79 et UOP/01/80 (tarsométatarses endommagés et partie distale d'un coracoïde droit de la limite Miocène-Pliocène de la formation de Bahía Inglesa. Le MNHN possède une partie proximale d'un carpométacarpe et l'extrémité d'un humérus droit provenant de la « formation de Pisco »[77] - [78] - [79] - [17] - [19].
- cf. Osteodontornis (Miocène moyen de Pisco –? Pliocène inférieur du Pérou) – deux espèces ? Spécimen 210 de l'Université nationale principale de San Marcos (fragments de bec et une vertèbre cervicale) ; spécimen 666 (tête proximale d'un humérus droit) ; spécimen 667 (partie proximale de l'ulna) d'un oiseau un peu plus petit que Pelagornis miocaenus, attribué auparavant par erreur à Pseudodontornis. Le crâne bien préservé dévoilé en 2009 présente des pseudo-dents similaires à celles de Osteodontornis, mais appartient semble-t-il à un oiseau plus grand[80] - [79] - [19].
- Pseudodontornis stirtoni (Miocène ou Pliocène de Motunau Beach, Nouvelle-Zélande) – parfois Neodontornis.
- Pelagornithidae gen. et esp. indét. (Pliocène inférieur de Yushima, Maesawa, Japon) – Osteodontornis ? Un humérus droit fragmentaire[13] - [72] - [49].
- cf. Pseudodontornis stirtoni (Pliocène moyen ; mudstone de Tangahoe, Hawera Nouvelle-Zélande). Radius proximal droit (McKee collection A080 183) et humérus distal droit (McKee collection A111 182) d'une espèce de grande taille[81] - [68].
- Pelagornithidae gen. et esp. indét. (Pléistocène inférieur de Dainichi, Kakegawa, Japon) – Osteodontornis ? Spécimen du Mizunami Fossil Museum 1801, un fémur droit distal d'une espèce de grande taille. À l'origine, identifié à tort comme un albatros[82] - [13] - [72].
- Pelagornis sp. (Pliocène supérieur de Californie, É.-U.)[83].
D'autres oiseaux du Paléogène (et, dans un cas, éventuellement du Crétacé supérieur), typiquement des taxons connus uniquement à partir des restes les plus fragmentaires, pourraient également être des Pelagornithidae. Ils ne sont pas classés ici, mais la grande taille des fossiles et les similitudes connues de certains os des oiseaux à pseudo-dents avec ceux d'autres lignées mériteraient une étude plus approfondie. Les genres en question sont Laornis, Proceriavis, Manu (en) et Protopelicanus (en)[84] - [85] - [86].
Bibliographie
- (en) Gerald Mayr, Paleogene Fossil Birds, Heidelberg & New York, Springer-Verlag, (ISBN 978-3-540-89627-2 et 3-540-89627-9).
- (en) Michael Begon, Colin A. Townsend et John Lander Harper, Ecology : From Individuals to Ecosystems, Wiley-Blackwell, , 4e éd. (ISBN 978-1-4051-1117-1, lire en ligne).
- (en) Robert W. Boessenecker et N. Adam Smith, « Latest Pacific basin record of a bony-toothed bird (Aves, Pelagornithidae) from the Pliocene Purisima Formation of California, U.S.A. », Journal of Vertebrate Paleontology, vol. 31, no 3, , p. 652–657.
- (en) Estelle Bourdon, « Osteological evidence for sister group relationship between pseudo-toothed birds (Aves: Odontopterygiformes) and waterfowls (Anseriformes) », Naturwissenschaften, vol. 92, no 12, , p. 586–591 (PMID 16240103, DOI 10.1007/s00114-005-0047-0, résumé).
- Estelle Bourdon, L'avifaune du Paléogène des phosphates du Maroc et du Togo : diversité, systématique et apports à la connaissance de la diversification des oiseaux modernes (Neornithes) (thèse de doctorat, Muséum national d'histoire naturelle), (lire en ligne).
- (en) Pierce Brodkorb, « Catalogue of fossil birds. Part 1 (Archaeopterygiformes through Ardeiformes) », Bulletin of the Florida State Museum, Biological Sciences, vol. 7, no 4, , p. 179–293 (lire en ligne).
- (en) Pierce Brodkorb, « Catalogue of Fossil Birds: Part 3 (Ralliformes, Ichthyornithiformes, Charadriiformes) », Bulletin of the Florida State Museum, vol. 11, no 3, , p. 99–220 (lire en ligne).
- (es) Martín Chávez et Marcelo Stucchi, « El registro de Pelagornithidae del Pacífico sudeste », dans Actas del Congreso Latinoamericano de Paleontología de Vertebrados 1, (lire en ligne), p. 26.
- (es) Martín Chávez, Marcelo Stucchi et Mario Urbina, « El registro de Pelagornithidae (Aves: Pelecaniformes) y la Avifauna Neógena del Pacífico Sudeste », Bulletin de l'Institut Français d’Études Andines, vol. 36, no 2, , p. 175–197 (lire en ligne).
- (en) Leslie Christidis et Walter E. Boles, Systematics and Taxonomy of Australian Birds, Collingwood, Victoria, Australia, CSIRO Publishing, , 277 p. (ISBN 978-0-643-06511-6, lire en ligne).
- (en) Josep del Hoyo (éd.), Andrew Elliott (éd.) et Jordi Sargatal (éd.), Handbook of the Birds of the World, vol. 1 : Ostrich to Ducks, Barcelone, Lynx Edicions, , 696 p. (ISBN 84-87334-10-5).
- (en) Denis Geraads, « The late Pliocene locality of Ahl al Oughlam, Morocco: vertebrate fauna and interpretation », Transactions of the Royal Society of South Africa, vol. 61, no 2, , p. 97–101 (DOI 10.1080/00359190609519958, lire en ligne).
- (en) James L. Goedert, « Giant Late Eocene Marine Birds (Pelecaniformes: Pelagornithidae) from Northwestern Oregon », Journal of Paleontology, vol. 63, no 6, , p. 939–944 (JSTOR 1305659).
- (en) Gerardo González-Barba, Tobias Schwennicke, James L. Goedert et Lawrence G. Barnes, « Earliest Pacific Basin record of the Pelagornithidae (Aves, Pelecaniformes) », J. Vertebr. Paleontol., vol. 22, no 2, , p. 722–725 (DOI 10.1671/0272-4634(2002)022[0722:EPBROT]2.0.CO;2, JSTOR 4524266).
- (en) James A. Hopson, « Pseudodontornis and other large marine birds from the Miocene of South Carolina », Postilla, vol. 83, , p. 1–19 (lire en ligne).
- (en) Hildegarde Howard, « A gigantic "toothed" marine bird from the Miocene of California », Santa Barbara Museum of Natural History Bulleting (Geology Department), vol. 1, , p. 1–23.
- (en) International Commission on Zoological Nomenclature (ICZN), International Code of Zoological Nomenclature, Londres, International Trust for Zoological Nomenclature, London, , 4e éd., 306 p. (ISBN 0-85301-006-4, lire en ligne).
- (en) Urless N. Lanham, « Notes on the phylogeny of the Pelecaniformes », The Auk, vol. 64, no 1, , p. 65–70 (DOI 10.2307/4080063, lire en ligne).
- (en) Bradley C. Livezey et Richard L. Zusi, « Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion », Zoological Journal of the Linnean Society, vol. 149, no 1, , p. 1–95 (PMID 18784798, PMCID 2517308, DOI 10.1111/j.1096-3642.2006.00293.x).
- (en) Hirosige Matsuoka, Fujio Sakakura et Fumio Ohe, « A Miocene pseudodontorn (Pelecaniformes: Pelagornithidae) from the Ichishi Group of Misato, Mie Prefecture, Central Japan », Paleontological Research, vol. 2, no 4, , p. 246–252 (lire en ligne).
- (en) Gerald Mayr, « A skull of the giant bony-toothed bird Dasornis (Aves: Pelagornithidae) from the Lower Eocene of the Isle of Sheppey », Palaeontology, vol. 51, no 5, , p. 1107–1116 (DOI 10.1111/j.1475-4983.2008.00798.x).
- (en) Gerald Mayr, Cornelis J. Hazevoet, Pedro Dantas et Mário Cachāo, « A sternum of a very large bony-toothed bird (Pelagornithidae) from the Miocene of Portugal », J. Vertebr. Paleontol., vol. 28, no 3, , p. 762–769 (DOI 10.1671/0272-4634(2008)28[762:ASOAVL]2.0.CO;2, lire en ligne).
- (en) Joseph W.A. McKee, « A pseudodontorn (Pelecaniformes: Pelagornithidae) from the middle Pliocene of Hawera, Taranaki, New Zealand », New Zealand Journal of Zoology, vol. 12, no 2, , p. 181–184 (DOI 10.1080/03014223.1985.10428278, lire en ligne).
- (de) K. Meunier, « Korrelation und Umkonstruktionen in den Größenbeziehungen zwischen Vogelflügel und Vogelkörper » [« Corrélation et restructuration dans la relation de taille entre l'aile et le corps aviaire »], Biologia Generalis, no 19, , p. 403–443.
- (en) Jirí Mlíkovský, « Tertiary avian localities of the United Kingdom », Acta Universitatis Carolinae: Geologica, vol. 39, , p. 759–771 (lire en ligne).
- (en) Jirí Mlíkovský, Cenozoic Birds of the World, Part 1 : Europe, Prague, Ninox Press, (lire en ligne) — La fusion des espèces doit être regardée avec une extrême précaution. La répartition des espèces est généralement considérée comme correcte. Voir cette recension : (en) Cécile Mourer-Chauvire, « Cenozoic Birds of the World, Part 1: Europe », The Auk, vol. 121, no 2, , p. 623-627 (lire en ligne).
- (en) Jirí Mlíkovský, « Early Miocene birds of Djebel Zelten, Libya », Časopis Národního muzea, Řada přírodovědná (Journal du musée national, série Histoire naturelle), no 172, , p. 114–120 (lire en ligne).
- (en) Jirí Mlíkovský, « Evolution of the Cenozoic marine avifaunas of Europe », Annalen des Naturhistorischen Museums Wien A, no 111, , p. 357–374 (lire en ligne).
- (en) Storrs L. Olson, chap. 2 « The Fossil Record of Birds », dans D.S. Farner et Kenneth C. Parkes (éds.), Avian Biology, (lire en ligne), p. 79–252.
- (en) Storrs L. Olson et Pamela C. Rasmussen, « Miocene and Pliocene Birds from the Lee Creek Mine, North Carolina », dans Clayton E. Ray et David J. Bohaska (éds.), Geology and Paleontology of the Lee Creek Mine, North Carolina, III, coll. « Smithsonian Contributions to Paleobiology » (no 90), (lire en ligne), p. 233–307.
- (en + ja) Keiichi Ono, « Pliocene Tubinare Bird from Kakegawa, Shizuoka Prefecture, Japan », Memoirs of the National Science Museum, vol. 13, no 1, , p. 29–34 (lire en ligne).
- (en) Keiichi Ono, « A Bony-Toothed Bird from the Middle Miocene, Chichibu Basin, Japan », Bulletin of the National Science Museum Series C: Geology & Paleontology, vol. 15, no 1, , p. 33–38 (lire en ligne).
- (en) « Taxonomic name search form », Paleobiology Database, (consulté le ).
- (en) D. Palmer, The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals, Londres, Marshall Editions, (ISBN 1-84028-152-9).
- (en) Jordan P. Price et David A. Clague, « How old is the Hawaiian biota? Geology and phylogeny suggest recent divergence », Proc. R. Soc. B, vol. 269, no 1508, , p. 2429–2435 (PMID 12495485, PMCID 1691179, DOI 10.1098/rspb.2002.2175).
- (en) Robert W. Purdy, Vincent P. Schneider, Shelton P. Applegate, Jack H. McLellan, Robert L. Meyer et Bob H. Slaughter, « The Neogene Sharks, Rays, and Bony Fishes from Lee Creek Mine, Aurora, North Carolina », dans Clayton E. Ray et David J. Bohaska (éds.), Geology and Paleontology of the Lee Creek Mine, North Carolina, III, coll. « Smithsonian Contributions to Paleobiology » (no 90), (lire en ligne), p. 71–202.
- (en) Pamela C. Rasmussen, « Early Miocene Avifauna from the Pollack Farm Site, Delaware », Delaware Geological Survey Special Publication, vol. 21, , p. 149–151 (lire en ligne).
- (es) R. Rincón, D. Ascanio et Marcelo Stucchi, « Primer registro de la familia Pelagornithidae (Aves: Pelecaniformes) para Venezuela », Boletín de la Sociedad Venezolana de Espeleología, vol. 37, , p. 27–30 (lire en ligne).
- (en) R.J. Scarlett, « Bone of a presumed odontopterygian bird from the Miocene of New Zealand », New Zealand Journal of Geology and Geophysics, vol. 15, no 2, , p. 269–274 (DOI 10.1080/00288306.1972.10421960, lire en ligne).
- (en) Appy Sluijs, Stefan Schouten, Mark Pagani, Martijn Woltering, Henk Brinkhuis, Sinninghe S. Jaap, Gerald R. Dickens, Matthew Huber, Gert-Jan Reichart, Rüdiger Stein, Jens Matthiessen, Lucas J. Lourens, Nikolai Pedentchouk, Jan Backman, Kathryn Moran et the Expedition 302 Scientists, « Subtropical Arctic Ocean temperatures during the Palaeocene/Eocene thermal maximum », Nature, vol. 441, no 7093, , p. 610–613 (PMID 16752441, DOI 10.1038/nature04668, lire en ligne) — notes complémentaires.
- (en) Jeffrey D. Stilwell, Craig M. Jones, Richard H. Levy et David M. Harwood, « First fossil bird from East Antarctica », Antarctic Journal of the United States, vol. 33, no 1, , p. 12–16 (lire en ligne).
- (en) Witmer Stone, « Recent Literature – Wetmore on Palæochenoides mioceanus Shufeldt », The Auk, vol. 35, no 2, , p. 251 (DOI 10.2307/4072894, lire en ligne).
- (en) Darren Naish, « A good year for terror birds and mega-ducks », The Tetrapod Zoology blog, .
- (es) Eduardo Pedro Tonni, « Un pseudodontornitido [Pelecaniformes, Odontopterygia] de gran tamaño, del Terciario temprano de Antártida », Ameghiniana, vol. 17, no 3, , p. 273–276 (résumé).
- (es) Eduardo Pedro Tonni et Claudia Patricia Tambussi, « Nuevos restos de Odontopterygia (Aves: Pelecaniformes) del Terciario temprano de Antártida », Ameghiniana, vol. 21, nos 2–4, , p. 121–124 (lire en ligne).
- (en) Digital Nomenclator Zoologicus, version 0.86, vol. 3, uBio, (lire en ligne), p. 387.
- (en) Stig A. Walsh (Discours prononcé lors du 48e symposium annuel sur la paléontologie des vertébrés et l'anatomie comparatives, le , Portsmouth, UK.), « Big-chested birds – exciting new avian material from the Neogene of Chile »,
- (en) Stig A. Walsh et Julian P. Hume, « A new Neogene marine avian assemblage from north-central Chile », J. Vertebr. Paleontol., vol. 21, no 3, , p. 484–491 (DOI 10.1671/0272-4634(2001)021[0484:ANNMAA]2.0.CO;2, lire en ligne).
- (en) Kenneth I. Warheit, « A Review of the Fossil Seabirds from the Tertiary of the North Pacific: Plate Tectonics, Paleoceanography, and Faunal Change », Paleobiology, vol. 18, no 4, , p. 401–424 (JSTOR 2400827).
- (en) Kenneth I. Warheit, « The seabird fossil record and the role of paleontology in understanding seabird community structure », dans E.A Schreiber et J. Burger (éds.), Biology of marine birds, Boca Raton, Florida, US, CRC Press, (ISBN 0-8493-9882-7, lire en ligne), p. 17–55.
- (en) Alexander Wetmore, « The Relationships of the Fossil Bird Palaeochenoides mioceanus », Journal of Geology, vol. 25, no 6, , p. 555–557 (lire en ligne).
- (en) Alexander Wetmore, « A check-list of the fossil and prehistoric birds of North America and the West Indies », Smithsonian Miscellaneous Collections, vol. 131, no 5, , p. 1–105 (lire en ligne).
- (en) Arthur Smith Woodward (éd.), A Guide to the Fossil Mammals and Birds in the Department of Geology and Palaeontology of the British Museum (Natural History), Londres, William Clowes and Sons Ltd., , 9e éd. (lire en ligne).
Voir aussi
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- Les seiches (Sepiida) et les calmars (Teuthida) se sont diversifiés durant le Paléogène, et les Argonautoida (Octopodes pelagiques modernes) datent de la même époque voire d'un peu plus tôt. Les octopodes basaux Keuppia, Palaeoctopus et Styletoctopus du Crétacé supérieur étaient aussi, partiellement, pélagiques. Dans la mesure où les premiers Pelagornithidae vivaient presque certainement au Crétacé supérieur, les Vampyromorphida alors diversifiés (seul, aujourd'hui subsiste le Vampire des abysses, Vampyroteuthis infernalis), étaient les proies potentielles des premiers oiseaux à pseudo-dents ; bien que les plus grands Pelagornithidae ont été trouvés dans des couches du Néogène, il est possible que des juvéniles de Tusoteuthis aient pu servir de nourriture à ces oiseaux[11].
- Les anguilles (Anguillidae) et les congres (Congridae) sont attestés depuis l'Éocène et ont peut-être leurs origines au début du Paléogène voire un peu avant. Les poissons-chats (Plotosidae), les Ophidiiformes (donzelles et autres), certains Blennioidei et peut-être les Zoarcidae sont supposés être d'âge similaire avec une origine de leurs lignées remontant au Crétacé supérieur. Si les oiseaux à pseudo-dents ont leur origine au Crétacé, comme cela est probable, leurs proies initiales pourraient avoir été les Enchodontoidei, lesquels s'éteignirent à la fin du Mésozoïque[11].
- Tels que Desmatophoca, Gomphotaria, Imagotaria, Pelagiarctos et Pliopedia. Quelques-uns avaient quatre défenses et certains, aucune[11].
- Le grand requin mégalodon (Carcharocles megalodon) aurait probablement trouvé que même le plus grand des oiseaux à pseudo-dents ne valait pas la peine de faire l'effort de le chasser…
- Les oiseaux aquatiques à queue courte et les « oiseaux aquatiques supérieurs » ont souvent des queues légères ou blanches[24].
- En particulier l'énigmatique Laornis edvardsianus[52].
Références
- Mayr 2009, p. 59.
- Meunier 1951.
- Hopson 1964.
- Olson 1985, p. 200.
- Mayr 2008.
- Mayr 2009, p. 58.
- Smith Woodward 1909, p. 86-87.
- Olson 1985, p. 142.
- Mayr et alii 2008.
- Olson 1985, p. 194-195.
- Paléodatabase.
- Olson 1985, p. 200–201.
- Ono 1989.
- del Hoyo et alii 1992, p. 198, 204.
- Warheit 1992.
- Warheit 2001.
- Rincón, Ascanio et Stucchi 2003.
- Sluijs et alii 2006.
- Chávez, Stucchi et Urbina 2007.
- Mayr 2009, p. 56–58, 217–218.
- Begon et alii 2005, p. 123–124.
- Mayr 2009, p. 217–218.
- Olson 1985, p. 195–199.
- del Hoyo et alii 1992.
- Olson et Rasmussen 2001.
- Price et Clague 2002.
- Mlíkovský 2002, p. 81–83.
- Mayr 2009, p. 6, 56.
- Geraads 2006.
- Mlíkovský 2009.
- Purdy et alii 2001.
- Olson 1985, p. 198.
- Mlíkovský 2002, p. 81.
- Mayr 2009, p. 55-59.
- Lanham 1947.
- Wetmore 1956, p. 12–14.
- Brodkorb 1963, p. 241, 262–264.
- Brodkorb 1963, p. 248–249.
- Brodkorb 1967, p. 141-143.
- Olson 1985, p. 195.
- Mlíkovský 2002, p. 78, 81–83.
- Smith Woodward 1909, p. 87.
- Brodkorb 1967, p. 142.
- Olson 1985, p. 195, 199.
- Bourdon 2005.
- Christidis et Boles 2008, p. 100.
- Wetmore 1918.
- Olson 1985, p. 199–200.
- González-Barba et alii 2002.
- Christidis et Boles 2008, p. 100, 105.
- Naish 2009.
- Mayr 2009, p. 21.
- Livezey et Zusi 2007.
- Stone 1918.
- Howard 1957.
- Mayr 2009.
- Olson 1985, p. 194–195.
- Mayr 2009, p. 55–59.
- Mlíkovský 2002, p. 81–84.
- Bourdon 2006.
- Mayr 2009, p. 56.
- Mayr 2009, p. 57.
- Stilwell et alii 1998.
- Mlíkovský 2002, p. 83.
- Olson 1985, p. 196, 199.
- Tonni 1980.
- Tonni et Tambussi 1985.
- Goedert 1989.
- Mlíkovský 1996.
- Mlíkovský 2002, p. 269.
- Mayr 2009, p. 31.
- Matsuoka et alii 1998.
- Rasmussen 1998.
- Scarlett 1972.
- Olson 1985, p. 199.
- Mlíkovský 2002, p. 84.
- Walsh 2000.
- Walsh et Hume 2001.
- Chávez et Stucchi 2002.
- Palmer 1999, p. 180.
- McKee 1985.
- Ono 1980.
- Boessenecker et Smith 2011.
- Olson 1985, p. 173, 202, 208.
- Mlíkovský 2002, p. 269–270.
- Mayr 2009, p. 21, 31, 77, 80.