Taniwhasaurus
Taniwhasaurus (littéralement « lézard de Taniwha ») est un genre éteint de mosasauridés ayant vécu durant le Crétacé supérieur (âges Santonien-Campanien) dans ce qui sont actuellement la Nouvelle-Zélande et l'Antarctique, voire possiblement le Japon et l'Afrique du Sud. Il appartient à la sous-famille des Tylosaurinae, une lignée de mosasaures caractérisée par un long rostre édenté de forme conique.

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Sauropsida |
| Ordre | Squamata |
| Super-famille | † Mosasauroidea |
| Famille | † Mosasauridae |
| Sous-famille | † Tylosaurinae |
Genres de rang inférieur
- † T. 'capensis' Broom, 1912
- † T. 'mikasaensis' Caldwell et al., 2008
- Lakumasaurus Novas et al., 2002
- Yezosaurus Muramoto & Obata, 1977
- Tylosaurus haumuriensis Hector, 1874
- Leiodon haumuriensis Hector, 1874
- Platecarpus oweni Lydekker, 1888
- T. mikasaensis ? Caldwell et al., 2008
- Lakumasaurus antarcticus Novas et al., 2002
- Tylosaurus capensis Broom, 1912
- Yezosaurus mikasaensis Muramoto & Obata, 1977
Taniwhasaurus est un représentant de taille moyenne pour un mosasauridé, les estimations maximales le mettant à environ 7 m de long. Bien que les fossiles attribués à ce genre sont très incomplets et soient principalement connu à partir de matériaux crâniens, la morphologie de Taniwhasaurus est globalement typique des tylosaurinés. Les rares fossiles du squelette axial indiquent que l'animal aurait eu une grande mobilité au niveau de la colonne vertébrale, mais que la queue générerait le principal mouvement propulsif, une méthode de nage proposée pour les autres mosasauridés. La constitution du membre antérieur en forme de pagaie chez Taniwhasaurus indique qu'il aurait eu de puissants palettes natatoires pour nager. Les analyses tomodensitométriques effectuées sur les foramens du museau de T. antarcticus montrent que Taniwhasaurus, à la manière de divers prédateurs aquatiques de nos jours, aurait probablement eu un organe électrosensible capable de détecter les mouvements d'une proie sous l'eau.
L'histoire taxonomique de Taniwhasaurus est complexe, plusieurs espèces ayants été décrits à partir des fossiles trop fragmentaires pour êtres considérés comme viables. Néanmoins, une révision phylogénétique des tylosaurinés publié en 2019 confirme qu'aux moins deux espèces, T. oweni et T. antarcticus, appartiennent au genre, les autres espèces nommées étant problématiques pour être considérées comme des espèces distinctes ou synonymes.
Historique des recherches
Taniwhasaurus oweni

La première espèce connue, Taniwhasaurus oweni, fut découvert durant les années 1860 dans la formation de Conway (en), situé dans les falaises d'Haumuri Bluff (en), à l'est de la Nouvelle-Zélande[1]. Cette formation est datée du Crétacé supérieur, est plus précisément du stade Campanien inférieur et moyen[2]. Les premiers fossiles formellement attribués à ce taxon furent décrite par le naturaliste écossais James Hector en 1874. Le matériel squelettique de T. oweni se composait d'un crâne, de vertèbres et de palettes natatoires, répartis en trois sections distinctes[1]. En 1888, constatant que les fossiles sont incomplets, Richard Lydekker place de manière incertaine T. oweni au sein du genre Platecarpus, étant renommé Platecarpus oweni[3]. En 1897, dans sa révision de la distribution des mosasaures, Samuel Wendell Williston remet Taniwhasaurus comme un genre distinct, mais le considère comme étant toujours proche de Platecarpus[4]. Hector n'ayant pas désigné de fossile holotype pour ce taxon, Samuel Paul Welles et D. R. Gregg désignent le spécimen NMNZ R1536, un crâne fragmenté constitué d'un os frontal et pariétal accompagné d'un os dentaire partiel, comme lectotype de T. oweni en 1971[5]. Le nom de genre Taniwhasaurus est composé du mot en Māori Taniwha (en), et du mot du en grec ancien σαῦρος / saûros, « lézard », le tout voulant littéralement dire « lézard de Taniwha », en référence à une créature aquatique surnaturelle de la mythologie maorie. L'épithète spécifique oweni est nommé en l'honneur du célèbre paléontologue anglais Richard Owen, qui fut le premier à décrire les reptiles marins du Mésozoïque de la Nouvelle-Zélande[6] - [7].
Dans son article, Hector décrit plusieurs restes squelettiques qu'il attribue à un autre mosasaure, qu'il nomme Leiodon haumuriensis[1]. En 1897, Williston suggère de transférer ce taxon au sein du genre Tylosaurus[4], une proposition qui sera effectuée en 1971, étant renommé pour le coup Tylosaurus haumuriensis. Welles et Gregg désignent également le spécimen NMNZ R1532 comme lectotype de Tylosaurus haumuriensis dans l'article[5]. En 1999, des nouveaux matériels crâniens et postcrâniens ont été découverts dans les falaises d'Haumuri Bluff et ces trouvailles sont officialisés par Michael W. Caldwell est ses collègues en 2005. Sur la base d'analyses approfondies de ces fossiles, les chercheurs découvrent qu'il y en fait peu de divergences morphologiques entre les deux taxons de mosasaures de cette localité, les différences étants principalement due à la taille plus imposante du spécimen NMNZ R1532, faisant de Tylosaurus haumuriensis un synonyme junior de T. oweni[2].

Taniwhasaurus antarcticus
En 2002, un nouveau taxon de mosasauridé est identifié à partir de fossiles découverts dans le membre Gamma de la formation de l'île Snow Hill, situé sur l'île James Ross en Antarctique[8]. La formation est datée dans les stades Santonien-Campanien du Crétacé supérieur, soit une datation similaire à celui de la formation néo-zélandaise de Conway[7]. Cette découverte concerne un spécimen de tylosauriné, catalogué IAA 2000-JR-FSM-1, contenant un crâne, une dent, quelques vertèbres et des fragments de côtes. Le crâne, mesurant 72 cm de long, est à lui-seul presque complet et articulé, ce qui est une première pour les mosasaures d'Antarctique car la grande majorité des fossiles de ces derniers furent souvent des dents isolées. Après analyse du matériel, Fernando E. Novas est ses collègues le nomment Lakumasaurus antarcticus. Le nom de genre Lakumasaurus vient de Lakuma, un esprit des mers de la mythologie du peuple Yagans, et du terme en grec ancien σαῦρος / saûros, « lézard », pour donner littéralement « lézard de Lakuma ». L'épithète spécifique antarcticus fait référence à l'Antarctique, lieu d'où à vécu l'animal[8] - [7].
En 2007, James E. Martin et Marta Fernández remettent en question la validité de Lakumasaurus en tant que genre distinct, notant que les caractéristiques crâniennes sont assez infimes pour justifier une telle proposition. Cependant, ils affirment qu'il y a suffisamment de différences pour classer Lakumasaurus antarcticus en tant que deuxième espèce du genre Taniwhasaurus, étant renommé T. antarcticus[7]. La même année, Martin et ses collègues annoncent la découverte d'un crâne juvénile considéré comme appartenant à la même espèce et datant du Maastrichtien[9], cependant, des études ultérieurs se montrent septiques quant à cette affirmation[10]. Moins de deux ans plus tard, en 2009, les mêmes auteurs publient un article qui décrit les relations phylogénétiques entre les espèces T. antarcticus et T. oweni[11], une relation qui s'avère être encore reconnue actuellement[12].
Taniwhasaurus 'capensis'
Au début du XXe siècle, plusieurs fossiles commencent à être collectés dans la région du Pondoland, en Afrique du Sud. Ces fossiles s'avèrent appartenir à des squamates et tortues marines datant du stade Santonien du Crétacé supérieur. En 1901, l'un des fossiles découverts, étants quelques morceaux fragmentaires de l'os d'une mâchoire, fut référencé comme appartenant à un reptile considéré comme proche du genre Mosasaurus. Cette collection de fossiles fut plus tard donné au paléontologue écossais Robert Broom, qui publie en 1912 un article décrivant les mêmes os, accompagné d'une vertèbre attribué à ce spécimen. Il en conclut que les fossiles appartiendraient à un grand représentant sud-africain du genre Tylosaurus, le nommant Tylosaurus capensis[13].

Pendant tout le reste du XXe siècle, Tylosaurus capensis fut généralement perçu comme une espèce valide au sein du genre, étant principalement identifié grâce à la taille du foramen pariétal et de la suture entre l'os frontal et pariétal[14]. Cependant, les deux caractéristiques sont très variables au sein du genre Tylosaurus et ne sont pas considérés comme diagnostiques au niveau de l'espèce[15]. En 2019, le réexamen des tylosaurinés menée par Paulina Jimenez-Huidobro et Caldwell révèle que le spécimen holotype, catalogué SAM-PK-5265, est plus caractéristique de Taniwhasaurus que de Tylosaurus, mais constatent également que les fossiles sont trop mal conservés pour être identifiés de manière définitive au genre. Néanmoins, l'étude déplace le taxon Tylosaurus capensis vers Taniwhasaurus[16]. En 2022, une révision anatomique des mosasaures d'Afrique du Sud rapproche le spécimen à T. 'mikasaenis' sur la base d'analyses dentaires et le considère comme appartenant au genre, bien que les auteurs affirment toujours que les différences sont trop minces pour justifier une nouvelle espèce[17].
Taniwhasaurus 'mikasaensis'
En , une grande partie avant du crâne d'un mosasaure fut découvert sur une rive de la rivière Ikushumbetsu à Hokkaidō, au Japon. Ce spécimen a été trouvé dans une concrétion flottante, et sa formation d'origine a été identifiée à la formation de Kashima, situé dans le groupe de Yezo (en), la localité étant la zone exposée de ce même lieu. Tout comme les sites précédemment mentionnés, la formation d'où fut trouvé cet animal est datée du stade Santonien-Campanien. Le spécimen, catalogué MCM.M0009, fut nommé Yezosaurus mikasaensis dans un communiqué de presse publié par K. Muramoto et Ikuwo Obata le [18], avant d'être classé à tort comme un dinosaure du groupe des tyrannosauroïdes dans un article publié par Muramoto en décembre de la même année[19]. Le nom de genre Yezosaurus vient de Yezo, le groupe contenant la formation de Kashima d'où le taxon a été découvert, et du grec ancien σαῦρος / saûros, « lézard », le tout voulant littéralement dire « lézard du Yezo ». L'épithète spécifique mikasaensis est nommé d'après la ville de Mikasa, lieu situé à proximité du site de la découverte[18] - [19]. Bien que ces deux publications ne peuvent pas être considérées comme valide du point de vue de la CINZ, Obata et Muramoto étaient effectivement perçus comme les auteurs de la description originale de Y. mikasaensis. Toujours la même année, et ceux avant même que le spécimen soit nommé, le ministère japonais de l'éducation décide de considérer le fossile comme trésor national du pays[20] - [21].
En 2008, le fossile est complètement réidentifié par Caldwell et ses collègues comme étant un mosasaure, et est classé comme une nouvelle espèce de Taniwhasaurus, étant renommé T. mikasaensis, gardant ainsi l'épithète spécifique d'Obata et Muramoto[20]. En 2019, la révision phylogénétique des tylosaurinés menée par Jimenez et Calwell considère toujours le spécimen comme un représentant du genre Taniwhasaurus, mais l'attribution à une quelconque espèce reste incertaine, le fossile étant insuffisant pour le classer soit dans T. mikasaensis, soit dans T. oweni[16]. En 2020, des scans 3D ont été effectués sur des répliques du spécimen, le véritable fossile nécessitant une autorisation spéciale du ministère japonais de l'éducation[21].
Description
Description simplifiée
Bien que les fossiles de Taniwhasaurus soient incomplet pour visualiser de manière formelle l'animal, les rares éléments squelettiques montrent que ce dernier a une anatomie grandement caractéristique des tylosaurinés[2] - [8] - [7]. Il s'avère aussi que le genre a une taille et un poids plutôt réduits pour un mosasaure, T. oweni ayant une longueur maximale estimée à environ 7 m pour un poids de 3 tonnes, contre 13 m pour 5,5 tonnes chez Mosasaurus et 14 m pour 6,7 tonnes chez Tylosaurus[N 1] - [22].
Squelette postcrânien

Le nombre exact de vertèbres chez Taniwhasaurus est inconnu, néanmoins, les rares fossiles concernants cette partie du corps inclut les vertèbres cervicales, dorsales, lombaires et caudales[N 2] - [1] - [2] - [8] - [23]. Comme chez les autres tylosaurinés, les condyles articulaires des vertèbres cervicales de Taniwhasaurus sont en légère dépression[8]. L'arc neural de l'atlas possède des processus qui auraient assuré la protection de la moelle épinière et le fixage des muscles qui maintiennent la tête. L'épine neurale de l'axis est robuste et allongée, culminant de manière postéro-dorsale en une pointe large, aplatie et incomplète qui portait probablement une coiffe cartilagineuse[2]. Les vertèbres dorsales sont procèles, et sont caractérisée pour avoir un plus grande diamètre au niveau antérieur que postérieur. Les surfaces articulaires sont placées de manière oblique vers l'arrière par rapport à l'axe général de la colonne vertébrale. L'arc neural est en continuité avec les parties antérieures des centra, et articulées par des processus transerves audacieuses. Le condyle des vertèbres dorsales est large et circulaire tandis que la parapophyse robuste s'étend latéralement sur une certaine distance[2].
Les vertèbres caudales ont des épines neurales hautes et droites qui ne possèdent aucun processus ou de facettes zygosphéno-zygantrales (en), une articulation que l'on retrouve chez la plupart des squamates. Les vertèbres caudales ont un tube neural de taille réduite et de forme triangulaire. Le centrum est raccourci du côté rostro-caudal mais est allongé dorso-ventralement et comprimé latéralement, ce qui donne un condyle ventralement ovale plutôt que circulaire comme on le voit dans les vertèbres présacrées[2] - [23]. Les vertèbres caudales de Taniwhasaurus ont des centra craniocaudal non fusionné au arc hémaux, ce qui est un cas typique chez les tylosaurinés[8]. Les arc hémaux s'articulent avec des hémapophyses profondes mais ne fusionnent pas avec elles. Distalement, les moitiés droite et gauche fusionnent à mi-chemin de la pointe ventrale de l'élément, créant une grande crête antérieure sur la colonne vertébrale[2] - [23].
Les côtes de T. oweni sont aplaties et quelque peu dilatées à leur insertion. Les rares côtes préservées de montre des surfaces articulaires convexes et ils s'avèrent être articulée sur une surface rugueuse, placée sur les parties antérieures et supérieures des centra vertébraux[1]. Bien que la ceinture scapulaire soit incomplètement connu chez Taniwhasaurus, il s'avère être de morphologie globalement similaire à ce que l'on retrouve chez les tylosaurinés en général[24]. Le coracoïde est beaucoup plus grand que l'omoplate, et ces deux os sont de forme convexe. La plaque du coracoïde est mince et distale par rapport au foramen du coracoïde, mais il n'y a aucune présence d'émargination sur le bord médial[2]. L'humérus est très court par rapport à sa largeur, étant de forme aplatie et ayant une articulation du coude très recourbé. Ce même humérus possède des crêtes musculaires prononcées. Les carpes sont de forme remarquablement plate et mince, leurs bordures étants relevées et rugueuses. Les rares fragments de phalanges indiquent que ces derniers auraient été de forme cylindrique et allongée. Cela suggère que Taniwhasaurus aurait eu un humérus musclé et puissant qui aurait été court et large, avec des os en forme de pagaie, indiquant qu'il aurait été un nageur efficace[1].
Classification
Taniwhasaurus fut toujours classé au sein des mosasaures, mais la description initiale publiée par Hector en 1874 ne l'attribue à aucun sous-taxon de cette famille[1] - [N 3]. En 1888, Taniwhasaurus est déplacé dans le genre Platecarpus par Lydekker, le considérant un synonyme junior[3]. En 1897, Williston nomme la sous-famille des Platercarpinae et classe Taniwhasaurus dans ce groupe, le considérant comme proche de certains genres tels que Platecarpus et Plioplatecarpus[4]. En 1967, le paléontologue Dale Russell synonymise les Platecarpinae avec les Plioplatecarpinae en raison du principe de priorité et de leurs définitions taxonomiques similaires[24] - [N 4]. C'est en 1971 que Taniwhasaurus est déplacé au sein des Tylosaurinae par Welles et Gregg, sur la base de caractéristiques crâniennes le rapprochant du genre Tylosaurus[5]. Les découvertes ultérieures d'autres tylosaurinés, précédemment mentionnés comme appartenant à des genres distincts et qui sont maintenant considérés comme synonymes de Taniwhasaurus, confirmeront la proposition de Welles et Gregg sur la position phylogénétique de ce genre[2] - [7] - [11] - [16] - [28] - [17]. Les membres de cette sous-famille, incluant le genre apparenté Tylosaurus et peut-être même Kaikaifilu, sont caractérisées par un rostre de forme conique et allongé qui ne porte pas de dents[2] - [20] - [11]. En 2019, dans leur révision phylogénétique de ce groupe, Jiménez-Huidobro et Caldwell estiment que Taniwhasaurus ne peut pas être considéré avec certitude comme étant monophylétique, car certaines espèces nommées ont des fossiles trop fragmentaires pour être attribuées de manière concrète au genre. Cependant, ils considèrent qu'en ignorant le matériel problématique, Taniwhasaurus forme un taxon bel et bien monophylétique et distinct de Tylosaurus[16]. Une étude publiée en 2020 par Daniel Madzia et Andrea Cau suggère une relation paraphylétique de Tylosaurus, considérant que Taniwhasaurus aurait évolué à partir de ce dernier, vers environ 84 millions d'années. Cependant, cette affirmation ne semble pas être cohérente avec les précédentes analyses phylogénétiques menées sur les deux genres[28].
Le cladogramme suivant est modifié à partir de l'analyse phylogénétique menée par Jiménez-Huidobro & Caldwell (2019), basée d'après les espèces de tylosaurinés avec les matériaux suffisamment connu pour modéliser des relations précises[16] - [N 5] :
| ◄ Mosasauroidea |
| ||||||||||||||||||||||||
Paléobiologie
Système neurovasculaire rostral
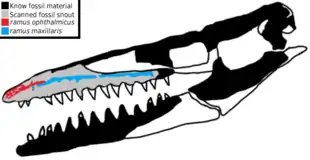
Une étude publiée en 2020 sur la base de la tomodensitométrie du rostre de l'holotype de T. antarcticus révèle la présence de plusieurs foramens internes situées dans la partie la plus en avant du museau. Ces foramens, les ramus maxillaris et les ramus ophthalmicus sont abondamment ramifiés et ont pour particularités d'êtres directement reliées au nerf trijumeau, indiquant qu'ils aurait envoyé des informations sensibles de la peau du museau au cerveau. Cela signifie que Taniwhasaurus aurait eu un organe électrosensible capable de détecter le moindre mouvement d'une proie sous l'eau. Ce système neurovasculaire est comparable à ceux présents chez divers tétrapodes aquatiques vivants ou éteints, tels que les cétacés, les crocodiliens, les plésiosaures et les ichthyosaures, qui sont utilisées pour traquer des proies dans des conditions de faibles luminosités[29].
L'étude mentionne que T. antarcticus est le premier mosasaure identifié à se disposer de tels structures pouvant expliquer ceci, mais il est probable que ce type d'organe soit présent chez les genres apparentés[29]. En effet, plusieurs mosasaures ont de grands foramens semblables à ceux présents chez Taniwhasaurus[24], ce qui semble indiquer une condition répandue au sein du groupe. De plus, les tylosaurinés semblent afficher le plus grand foramen au niveau du museau parmi les mosasaures. Cette condition peut être mise en corrélation avec le rostre édenté qui caractérise la morphologie de cette sous-famille, mais d'autres études sont nécessaires pour valider ces deux hypothèses[29].
Mécanique du cou
Les prézygapophyses de T. antarcticus ne sont pas autant développées, ce indiquerait que cette musculature serait moins prononcée que chez les autres mosasaures. Les prézygapophyses des vertèbres cervicales marquent l'emplacement des muscles longissimus et semi-épineux, qui produisent en partie les flexions latérales du corps chez les reptiles. Le peu de développement de crêtes au niveau des cervicales indiquent que la surface de préhension des muscles nommés serait par conséquent plus petite que chez les autres mosasaures, ainsi que la force produite par ces muscles. T. antarcticus aurait donc eu une grande capacité de mouvement latéral du cou, bien que les muscles qui y sont ancrés n'auraient pas eu une grande force. Dans le même sens, les prézygapophyses réduites indiqueraient que les vertèbres cervicales avaient une connexion plus lâche les unes aux autres, car elles présentent une réduction de surface d'articulation entre eux. Le genre apparenté Tylosaurus n'aurait pas eu une mobilité du cou trop prononcée en raison des épines neurales penchées vers l'arrière, qui attachent plus étroitement une vertèbre avec une autre au moyen de ligaments et de la musculature axiale. Bien que les vertèbres n'ont pas été retrouvés avec les épines neurales complètes chez Taniwhasaurus, les valeurs de compression des centra indiquent que bien qu'il ait pu avoir une certaine restriction au mouvement latéral, cela aurait été de toute façon plus prononcé[23].
Mobilité

L'analyse des vertèbres dorsales et caudales est complexe en raison la mauvaise préservation du matériel fossile de Taniwhasaurus, qui ne préserve pas les apophyses transverses ni les épines neurales. Plusieurs révisions anatomiques réalisée grâces au vertèbres caudales accordent une grande importance aux épines neurales et hemales, les conclusions ne pouvant pas être facilement appliquées pour Taniwhasaurus. Les vertèbres caudales de T. antarcticus suivent un schéma très similaire à celui trouvé à la fois chez Plotosaurus et Tylosaurus, où la robustesse des vertèbres est prise comme paramètre pour quantifier la raideur vertébrale dans différentes zones. Certaines des régions caudales peut être distingué, comme c'est le cas des vertèbres pygales. Ceux-ci sont interprétés comme une zone d'appui qui aurait une grande flexibilité. Cette partie des vertèbres caudales est constituée de vertèbres très similaires en morphologie les uns aux autres, et n'est représenté dans Taniwhasaurus que par des vertèbres caudales intermédiaires[23].
Les vertèbres caudales terminales seraient le support de la nageoire caudale et, comme chez Plotosaurus, ceux-ci ont une section subcirculaire dans la région antérieur et se transforme en une forme ovoïde comprimée latéralement en arrière. Cependant, cette configuration ne permet pas d'évaluer s'il y a ou non une tendance au nombre élevée de vertèbres pygales au détriment des caudales intermédiaires, comme on le voit dans les mosasaurinés dérivées. Il fut suggéré que les Rusellosaurina, le clade incluant les tylosaurinés et les lignées apparentés, avaient un squelette axial plésiomorphe et que par conséquent leur nage serait moins développé[14], tout le contraire des mosasaurinés, qui eux auraient eu une nage carangiforme, c'est-à-dire des formes où la queue est la principale source de propulsion, tandis que la partie la plus antérieure du corps maintient le mouvement restreint[23]. Cependant, une thèse publié en 2017 prouve que Tylosaurus avait eu une nage puissante et rapide, due notamment à la régionalisation des vertèbres caudales, bien que moins marquée que chez le mosasaurinés plus dérivées[30].
Les analyses concernant les vertèbres dorsales et caudales chez Plotosaurus et Tylosaurus est similaire à celle que l'on retrouve chez les cétacés actuels, et que donc ceux-ci auraient aussi une forme de nage carangiforme. Les mesures relatives des centra vertébraux, des proximité morphologiques et phylogénétiques avec Tylosaurus, semblent indiquer que la queue de T. antarcticus aurait aussi un rôle très important en déplacement, confortant cette hypothèse. Cependant les vertèbres cervicales de Taniwhasaurus montre une gamme de mouvement inhabituel dans une nage carangiforme, peut-être plus large que dans tout autre mosasaure en raison de la compression latérale des centra vertébraux de cette zone, mais aussi à leur longueur. Basée sur ces preuves, il est admis que bien que toute la colonne vertébrale de T. antarcticus aurait eu une grande mobilité, la queue serait la principale source de propulsion, soutenant la tendance à des formes plus carangiformes, plaçant Taniwhasaurus quelque part entre les formes basales en forme d'anguille et les formes dérivées des carangiformes[23]. Cela se trouve dans l'accord avec la position phylogénétique de ce taxon[16].
Paléoécologie
Les archives fossiles montrent que Taniwhasaurus a eu une distribution principalement répandue dans ce qui fut autrefois le Gondwana, ses fossiles ayant été trouvés dans des dépôts marins concernant les parties de l'ancien supercontinent. Bien que T. 'mikasaensis' fut retrouvé au Japon, les deux espèces actuellement valides T. antarcticus et T. oweni auraient été probablement endémique du Gondwana[7] - [20].
Nouvelle-Zélande
T. oweni est connu de la formation de Conway (en), est plus particulièrement d'Haumuri Bluff (en), une localité contenant des fossiles datant du Campanien inférieur et moyen. La partie précise du site atteint une épaisseur maximale de 240 m et du point de vue lithologique, l'unité est un siltite gris massif lâchement cimenté avec des interlits localement limités de grès fin. Le noyau des concrétions présents dans la formation semblent être des os fossilisés, des coquillages ou même du bois, indiquant que l'environnement de dépôt aurait été la zone inférieure d'un estran[2]. Relativement peu de grands vertébrés sont connus au sein du site d'après les sources, le seul clairement identifié étant la grande raie rajiforme Australopristis (en)[31].
Antarctique
T. antarcticus est connu des dépôts datant du Campanien supérieur de la péninsule Antarctique, dans la formation de l'île Snow Hill, situé sur l'île James Ross[8] - [7]. Le taxon est principalement connu du membre Gamma, un site très diversifié contenant de nombreux fossiles de faunes marines et terrestres. Ce lieu se compose d'environ 200 m de grès et de coquina à l'intérieur du plateau, dominée principalement par les mollusques du groupe des bivalves et des gastéropodes. Les grès sont pour la plupart à grain fin, bien triés, formant des lits massifs ou stratifiés en parallèle, avec une stratification occasionnelle des vagues et des ondulations de courant. Plusieurs poissons osseux sont présents, notamment les ichtyodectiformes, les aulopiformes, les albuliformes, ainsi qu'un téléostéen indéterminée. Les poissons cartilagineux sont principalement représentés par les holocéphales et les requins. Les holocéphales comprennent les chimères, les callorhinchidés, les rhinochimaeridés ainsi que l'imposante espèce Edaphodon snowhillensis, qui est l'un des plus grands chimériformes ayant été identifié à ce jour. Les requins présents dans la zone incluent les hexanchiformes, les lamniformes, les squatiniformes, les squaliformes et les synechodontiformes. Plusieurs reptiles marins sont connus de cette localité[32], mais les mosasaures ne semblent pas être aussi diversifiés que dans les autres formations géologiques voisines d'Antarctique[10]. Les seuls clairement identifiés au sein du membre Gamma sont T. antarcticus ainsi qu'une espèce indéterminée du genre Hainosaurus. Les seuls plésiosaures connus du membre Gamma sont des incertae sedis appartenant soit aux élasmosauridés ou sont considérés comme indéterminée. Les dinosaures sont aussi répertoriés dans cette formation, notamment l'ankylosaure Antarctopelta, l'ornithopode Trinisaura ainsi qu'un sauropode lithostrotien non nommé, ce dernier étant le premier sauropode connu d'Antarctique[32].
Notes et références
Notes
- Ces estimations sont basées sur les plus grandes espèces connues, à savoir Mosasaurus hoffmannii et Tylosaurus proriger, les autres espèces de ces deux genres étant de taille plus ou moins réduite[22].
- La majorité des vertèbres connus de Taniwhasaurus proviennent de T. oweni[1] - [2], celles de T. antarcticus n'étant connu que par une vertèbre cervicale, dorsale et caudale[8] - [7] - [23].
- Dans l'article original publié par Hector en 1874, Taniwhasaurus est classé de manière simplifiée dans l'ordre des Pythonomorpha (en), un taxon censé englober les mosasaures et les ancêtres des serpents[1] - [25]. La validité de ce clade de squamates fait toujours débat, certains auteurs considérants les mosasaures comme étant plus proche des varans. Cependant, des analyses phylogénétiques récentes soutiennent que les serpents seraient les plus proches parents actuels des mosasaures, une position se rapprochant de la définition originelle du taxon[26] - [27].
- La famille Plioplatecarpidae fut nommé par Louis Dollo en 1884[25], tandis que le taxon Platecarpinae fut nommé par Williston en 1897[4]. Selon les règlements de la CINZ, saufs exceptions justifiables, si un taxon s'avère être un synonyme junior d'un autre taxon précédemment nommé, alors le premier nom doit être gardé. Russell, constatant que la définition des deux taxons sont similaires, fait déplacer les Plioplatecarpidae en tant que sous-famille, le renommant en Plioplatecarpinae[24].
- Plusieurs analyses phylogénétiques ont été effectué sur Taniwhasaurus et Tylosaurus, incluant même les espèces problématiques, cependant, les analyses plus strictes ne gardent que les espèces disposant du matériel fossile le mieux préservée[16].
Références
- (en) James Hector, « On the fossil Reptilia of New Zealand », Transactions and Proceedings of the Royal Society of New Zealand, vol. 6, , p. 333-358 (lire en ligne).
- (en) Michael W. Caldwell, Robert Holmes, Gorden L. Bell Jr. et Joan Wiffen, « An unusual tylosaurine mosasaur from New Zealand: A new skull of Taniwhasaurus oweni (Lower Haumurian: Upper Cretaceous) », Journal of Vertebrate Paleontology, vol. 25, no 2, , p. 393-401 (DOI 10.1671/0272-4634(2005)025[0393:AUTMFN]2.0.CO;2, JSTOR 4524453, S2CID 130434185, lire en ligne).
- (en) Richard Lydekker, Catalogue of the Fossil Reptilia and Amphibia in the British Museum. Part I, Londres, 270 p. (lire en ligne).
- (en) Samuel W. Williston, « Range and distribution of the mosasaurs with remarks on synonymy », Kansas University Quarterly, vol. 6, no 4, , p. 177-185 (lire en ligne).
- (en) S. P. Welles et D. R. Gregg, « Late Cretaceous marine reptiles of New Zealand », Records of the Canterbury Museum, vol. 9, , p. 1-111.
- (en) Ben Creisler, « Mosasauridae Translation and Pronunciation Guide » [archive du ], sur Dinosauria On-line, .
- (en) James E. Martin et Marta Fernández, « The synonymy of the Late Cretaceous mosasaur (Squamata) genus Lakumasaurus from Antarctica with Taniwhasaurus from New Zealand and its bearing upon faunal similarity within the Weddellian Province », Geological Journal, vol. 42, no 2, , p. 203-211 (DOI 10.1002/gj.1066, S2CID 128429649, lire en ligne [PDF]).
- (en) Fernando E. Novas, Marta S. Fernandez, Zulma B. de Gasparini, Juan M. Lirio, Héctor J. Nuñez et Pablo Puerta, « Lakumasaurus antarcticus, n. gen. et sp., a new mosasaur (Reptilia, Squamata) from the Upper Cretaceous of Antarctica », Ameghiniana, vol. 39, no 2, , p. 245-249 (hdl 11336/136746, S2CID 128304133, lire en ligne).
- (en) J. Martin, A. Kihm, M. Fernández, M. Reguero et J. Case, « A juvenile mosasaur (Taniwhasaurus antarcticus) from the Late Cretaceous of Antarctica », Journal of Vertebrate Paleontology, vol. 27, , p. 112A (DOI 10.1080/02724634.2007.10010458).
- (en) Martin S. Fernandez et Zulma Gasparini, « Campanian and Maastrichtian mosasaurs from Antarctic Peninsula and Patagonia, Argentina », Bulletin de la Société Géologique de France, vol. 183, no 2, , p. 93-102 (DOI 10.2113/gssgfbull.183.2.93, S2CID 129228056).
- (en) Marta Fernández et James E. Martin, « Description and phylogenetic relationships of Taniwhasaurus antarcticus (Mosasauridae, Tylosaurinae) from the upper Campanian (Cretaceous) of Antarctica », Cretaceous Research, vol. 30, no 3, , p. 717-726 (DOI 10.1016/j.cretres.2008.12.012, S2CID 129028759).
- (en) Paulina Jiménez-Huidobro et Michael W. Caldwell, « A New Hypothesis of the Phylogenetic Relationships of the Tylosaurinae (Squamata: Mosasauroidea) », Frontiers in Earth Science, vol. 7, , p. 47 (DOI 10.3389/feart.2019.00047
 , Bibcode 2019FrEaS...7...47J, S2CID 85513442).
, Bibcode 2019FrEaS...7...47J, S2CID 85513442). - (en) Robert Broom, « On a species of Tylosaurus from the Upper Cretaceous beds of Pondoland », Annals of the South African Museum, vol. 1, , p. 332-333 (lire en ligne).
- (en + fr) Theagarten Lingham-Soliar, « The tylosaurine mosasaurs (Reptilia, Mosasauridae) from the Upper Cretaceous of Europe and Africa », Bulletin de l'Institut royal des sciences naturelles de Belgique, vol. 62, , p. 171-194 (lire en ligne [archive du ] [PDF]).
- (en) Paulina Jimenez-Huidobro et Michael W. Caldwell, « Reassessment and reassignment of the early Maastrichtian mosasaur Hainosaurus bernardi Dollo, 1885, to Tylosaurus Marsh, 1872 », Journal of Vertebrate Paleontology, vol. 36, no 3, , e1096275 (DOI 10.1080/02724634.2016.1096275, S2CID 87315531).
- (en) Paulina Jiménez-Huidobro et Michael W. Caldwell, « A New Hypothesis of the Phylogenetic Relationships of the Tylosaurinae (Squamata: Mosasauroidea) », Frontiers in Earth Science, vol. 7, , p. 47 (DOI 10.3389/feart.2019.00047 , Bibcode 2019FrEaS...7...47J, S2CID 85513442)
- (en) Megan R. Woolley, Anusuya Chinsamy et Michael W. Caldwell, « Unraveling the taxonomy of the South African mosasaurids », Frontiers in Earth Science, vol. 10, , p. 971968 (DOI 10.3389/feart.2022.971968 , Bibcode 2022FrEaS..10.1968W, S2CID 254565690).
- (ja) K. Muramoto et Ikuwo Obata, « A way to dinosaur –discovery of Yezosaurus– », Hokuensya, Sapporo, , p. 115.
- (ja) K. Muramoto, « A discovery of a skull fossil of a large reptile », Kaseki No Tomo, vol. 16, no 12, , p. 2.
- (en) Michael W. Caldwell, Takuya Konishi, Ikuwo Obata et Kikuwo Muramoto, « A new species Of Taniwhasaurus (Mosasauridae, Tylosaurinae) from the upper Santonian-lower Campanian (Upper Cretaceous) of Hokkaido, Japan », Journal of Vertebrate Paleontology, vol. 28, no 2, , p. 339-348 (DOI 10.1671/0272-4634(2008)28[339:ANSOTM]2.0.CO;2, JSTOR 20490955, S2CID 129446036).
- (en) Kumiko Matsui et Tomoki Karasawa, « 3D models related to the publication: Interacting with the inaccessible: utilization of multimedia-based visual contents of Japan’s National Monument, the Taniwhasaurus mikasaensis (Mosasauridae) holotype for educational workshops at Mikasa City Museum », MorphoMuseuM, vol. 6, , e106 (DOI 10.18563/journal.m3.106 , S2CID 241798505).
- (en) Gregory S. Paul, The Princeton Field Guide to Mesozoic Sea Reptiles, Princeton University Press, , 166-176 p. (ISBN 978-0-69-119380-9, DOI 10.2307/j.ctv2hnkc6h, S2CID 251553177).
- (es + en) Gerardo Álvarez Herrera, « Análisis osteológico de Taniwhasaurus antarcticus (Mosasauroidea, Tylosaurinae). Implicancias paleobiológicas », Universidad de Buenos Aires, (lire en ligne [PDF]).
- (en) Dale A. Russell, Systematics and morphology of American mosasaurs, vol. 23, New Haven, Bulletin of the Peabody Museum of Natural History, , 240 p. (OCLC 205385, lire en ligne).
- Louis Dollo, « Le Mosasaure », Revue des Questions Scientifiques, vol. 16, , p. 648-653 (lire en ligne).
- (en) Tod W. Reeder, Ted M. Townsend, Daniel G. Mulcahy, Brice P. Noonan, Perry L. Wood, Jr., Jack W. Sites, Jr. et John J. Wiens, « Integrated Analyses Resolve Conflicts over Squamate Reptile Phylogeny and Reveal Unexpected Placements for Fossil Taxa », PLOS ONE, vol. 10, no 3, , e0118199 (PMID 25803280, PMCID 4372529, DOI 10.1371/journal.pone.0118199 , Bibcode 2015PLoSO..1018199R).
- (en) R. Alexander Pyron, « Novel Approaches for Phylogenetic Inference from Morphological Data and Total-Evidence Dating in Squamate Reptiles (Lizards, Snakes, and Amphisbaenians) », Systematic Biology, vol. 66, no 1, , p. 38-56 (PMID 28173602, DOI 10.1093/sysbio/syw068 , S2CID 3697004, lire en ligne [PDF]).
- (en) Daniel Madzia et Andrea Cau, « Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas », PeerJ, vol. 8, , e8941 (PMID 32322442, PMCID 7164395, DOI 10.7717/peerj.8941 ).
- (en) Gerardo Álvarez-Herrera, Federico Agnolin et Fernando Novas, « A rostral neurovascular system in the mosasaur Taniwhasaurus antarcticus », The Science of Nature, vol. 107, no 3, , p. 19 (PMID 32333118, DOI 10.1007/s00114-020-01677-y, Bibcode 2020SciNa.107...19A, hdl 11336/133328 , S2CID 216111650, lire en ligne).
- (en) J. A. Carpenter, « Locomotion and skeletal morphology of Late Cretaceous mosasaur, Tylosaurus proriger », Georgia Southern University, (résumé, lire en ligne).
- (en) David M. Martill et Nizar Ibrahim, « Aberrant rostral teeth of the sawfish Onchopristis numidus from the Kem Kem beds (?early Late Cretaceous) of Morocco and a reappraisal of Onchopristis in New Zealand », Journal of African Earth Sciences, vol. 64, , p. 71-76 (ISSN 1464-343X, DOI 10.1016/j.jafrearsci.2011.11.009, Bibcode 2012JAfES..64...71M, S2CID 129451161).
- (en) Marcelo A. Reguero, Zulma Gasparini, Eduardo B. Olivero, Rodolfo A. Coria, Marta S. Fernández, José P. O. Gorman, Soledad Gouiric-Cavalli, Carolina Acosta Hospitaleche, Paula Bona, Ari Iglesias, Javier N. Gelfo, María E. Raffi, Juan José Moly, Sergio N. Santillana et Magalí Cárdenas, « Late Campanian-Early Maastrichtian Vertebrates From The James Ross Basin, West Antarctica: Updated Synthesis, Biostratigraphy, And Paleobiogeography », Anais da Academia Brasileira de Ciências, vol. 94, no 1, , e20211142 (PMID 35674550, DOI 10.1590/0001-3765202220211142 , S2CID 249359371).
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :