Tyrannosauroidea
Tyrannosauroïdes
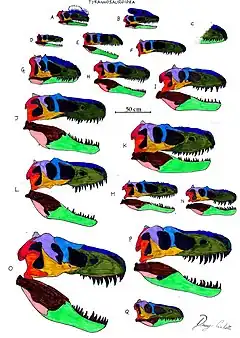
Taxons de rang inférieur
- †Pantyrannosauria
- †Alectrosaurus
- †Aviatyrannis
- †Bagaraatan
- †Dilong
- †Jinbeisaurus
- †Moros
- †Santanaraptor
- †Suskityrannus
- †Timimus
- †Timurlengia
- †Xiongguanlong
- †Teihivenator
- †Timurlengia
- † Stokesosauridae
- † Eutyrannosauria (dont Tyrannosauridae)
- †Proceratosauridae
- Classement incertain :
Les Tyrannosauroidea, ou tyrannosauroïdes (« de la forme des tyrannosaures ») forment une super-famille (ou clade) éteinte de dinosaures théropodes coelurosauriens comprenant les Tyrannosauridae ainsi que des genres plus basaux qui leur sont étroitement apparentés.
À la fin du Crétacé, ils étaient les grands prédateurs dominants dans l'hémisphère nord, culminant avec l'apparition du gigantesque Tyrannosaurus. Les fossiles de Tyrannosauroidea ont été recueillis dans ce que sont maintenant les continents de l'Amérique du Nord, de l'Europe, de l'Asie, de l'Amérique du Sud et l'Australie.
Les Tyrannosauroidea étaient des carnivores bipèdes, comme l'étaient la plupart des théropodes, et ont été caractérisés par de nombreuses particularités du squelette, notamment le crâne et le bassin. Au début de leur existence, les Tyrannosauroidea étaient de petits prédateurs avec de longues pattes avant munies de trois doigts. Les genres du Crétacé supérieur sont devenus beaucoup plus grands, incluant certains des plus grands prédateurs terrestres de tous les temps, mais la plupart de ces genres plus récents avaient des pattes avant proportionnellement plus petites avec seulement deux doigts. Des plumes primitives ont été identifiées dans les restes fossiles de deux espèces, et sont peut-être également présentes chez d'autres Tyrannosauroidea. Des crêtes osseuses remarquables dans la variété de formes et de tailles, présentes sur les crânes de nombreux Tyrannosauroidea, ont peut-être servi de fonction ornementale.
Description
Les Tyrannosauroidea varient considérablement en taille, mais il y a une tendance générale à l'augmentation de la taille au fil du temps. Les premiers Tyrannosauroidea sont de petits animaux[1]. Un spécimen de Dilong, dont la croissance était presque achevée, mesure 1,6 mètre de long[2], et un Guanlong adulte 3 mètres[3]. Des dents découvertes dans le Crétacé inférieur (140 à 136 millions d'années) de Hyōgo au Japon, semblent provenir d'animaux d'une longueur d'environ 5 mètres, indiquant peut-être une augmentation de la taille au début de la lignée[4]. Un spécimen immature d'Eotyrannus a une longueur plus de 4 mètres, et celle d'un Appalachiosaurus subadulte est estimée à plus de 6 mètres[1], indiquant que les deux genres atteignaient de plus grandes tailles. Les tyrannosaures du Crétacé supérieur varient de 9 mètres pour Albertosaurus et Gorgosaurus jusqu'à plus de 13 mètres pour Tyrannosaurus, ce dernier pouvant avoir pesé plus de 6 400 kilogrammes. Une étude de 2010 a conclu que les Tyrannosauroidea étaient « de petite à taille moyenne » dans leurs 80 premiers millions d'années, mais étaient « parmi les plus grands carnivores terrestres ayant jamais vécu » dans leurs 20 derniers millions d'années d'existence[5].

Les crânes des premiers Tyrannosauroidea sont longs, bas et de faible constitution, semblables à d'autres coelurosauriens, tandis que les espèces plus récentes ont des crânes plus grands et plus massifs. Malgré les différences de forme, certaines caractéristiques du crâne se trouvent chez tous les Tyrannosauroidea connus. L'os prémaxillaire est très grand, coupant la pointe du museau, une caractéristique qui a évolué de façon convergente chez les abélisauridés. Les os du nez sont typiquement fusionnés, voûtés légèrement vers le haut et souvent très grossièrement texturés sur leur surface supérieure. Les dents prémaxillaires à l'avant de la mâchoire supérieure sont façonnées différemment du reste de la dentition, de taille plus petite et ayant une section transversale en forme de D. Dans la mâchoire inférieure, une nervure proéminente sur le surangulaire s'étend latéralement de juste en dessous de l'articulation de la mâchoire, à l'exception de Guanlong, un membre basal des Tyrannosauroidea[1] - [2] - [3].

Les Tyrannosauroidea avaient le cou en forme de S et de longues queues, comme la plupart des autres théropodes. Les genres les plus anciens ont de longues pattes avant, environ 60 % de la longueur du membre postérieur chez Guanlong, avec les trois doigts typiques des coelurosauriens[3]. Les longues pattes antérieures ont persisté jusqu'au moins Eotyrannus, du Crétacé inférieur[6], mais sont inconnues pour Appalachiosaurus[7]. Les tyrannosauridés dérivés ont des membres antérieurs fortement réduits, l'exemple le plus extrême étant Tarbosaurus, genre découvert en Mongolie, où l'humérus est d'une longueur de seulement un quart de celle du fémur[1]. Le troisième doigt de la patte antérieure s'est également réduit également au fil du temps. Ce doigt est non réduit chez Guanlong, genre basal[3], et pour Dilong il est plus mince que les deux autres doigts[2]. Eotyrannus possède également trois doigts fonctionnels sur chaque main[6]. Les tyrannosauridés n'ont que deux doigts, bien que le métacarpe vestigial du troisième est présent chez quelques spécimens bien conservés[8]. Comme pour la plupart des coelurosauriens, le deuxième doigt de la main est le plus grand, même si le troisième doigt n'est pas présent.
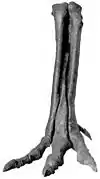
Les membres postérieurs de tous les Tyrannosauroidea, comme la plupart des théropodes, possèdent quatre doigts, bien que le premier orteil (hallux) ne touche pas le sol. Les membres postérieurs des Tyrannosauroidea sont plus longs par rapport à la taille du corps que ceux de presque tous les autres théropodes, et montrent des proportions caractéristiques d'animaux pouvant courir rapidement, avec des tibias et métatarses allongés[1].
Les caractéristiques du bassin des Tyrannosauroidea comprennent une encoche concave à l'extrémité supérieure avant de l'os iliaque, une arête verticale bien définie sur la surface extérieure de l'os iliaque, s'étendant vers le haut à partir de l'acetabulum (cotyle), et une énorme « botte » sur l'extrémité du pubis, d'une taille supérieure à plus de la moitié de la tige du pubis elle-même[1]. Ces caractéristiques se retrouvent chez tous les Tyrannosauroidea connus, y compris les membres basaux Guanlong[3] et Dilong[2]. Le pubis n'est pas connu pour Aviatyrannis et Stokesosaurus mais leur l'os iliaque présente les caractères typiques des Tyrannosauroidea[9].
Classification
Tyrannosaurus a été nommé par Henry Fairfield Osborn en 1905, avec la famille Tyrannosauridae[10]. Le nom est dérivé des mots grecs anciens τυραννος [tyrannos] (« tyran ») et σαυρος [sauros] (« lézard »). Le nom de la super-famille Tyrannosauroidea a été publié dans un article de 1964 par le paléontologue britannique Alick Walker[11]. Le suffixe -oidea, couramment utilisé dans le nom de superfamilles des animaux, est dérivé du grec ειδος [eidos], signifiant « forme ».
Les scientifiques ont généralement inclus dans la super-famille Tyrannosauroidea les tyrannosaures et leurs ancêtres immédiats[11] - [12]. Cependant, avec l'avènement de la taxonomie phylogénétique en paléontologie des vertébrés, le clade a reçu plusieurs définitions plus explicites. La première est formulée par Paul Sereno en 1998, où Tyrannosauroidea est définie comme un groupe-souche, comprenant toutes les espèces partageant un ancêtre commun plus récent avec Tyrannosaurus rex qu'avec les oiseaux modernes (Neornithes)[13]. Pour rendre le groupe plus exclusif, Thomas Holtz redéfinit celui-ci en 2004 par l'inclusion de toutes les espèces plus proches de Tyrannosaurus rex que d'Ornithomimus velox, Deinonychus antirrhopus ou Allosaurus fragilis[1]. Sereno publie une nouvelle définition en 2005, en excluant Ornithomimus edmontonicus, Velociraptor mongoliensis et Troodon formosus du clade Tyrannosauroidea[14]. Cette définition est adoptée dans un article de 2010[5].
Deux études publiées en 2012 et 2014 ont suggéré que le clade des Megaraptora, habituellement considérés comme des Allosauroidea, soient des Tyrannosauroidea basaux[15] - [16]. Cette classification est contestée par d'autres auteurs[17] - [18].
Phylogénie

Loewen et ses collègues ont réalisé en 2013 une classification comprenant pratiquement toutes les espèces connues de Tyrannosauroidea[19], conduisant au cladogramme ci-après.
En 2016, Stephen Brusatte et Thomas Carr ont effectué une revue de l'analyse phylogénétique des Tyrannosauroidea et de leur évolution (maximum de parcimonie)[20] :
- Tyrannosauroidea
En 2018, un nouvel arbre a été publié par Rafael Delcourt et Orlando Nelson Grillo[21]. Ils y ont placé Santanaraptor et Timimus, deux genres dont les placements étaient controversés [21]. Ils ont également nommé deux nouveaux sous-clades : les Pantyrannosauria qui constituent le groupe frère des procératosauridés, et les Eutyrannosauria qui comprennent les genres les plus dérivés dont les tyrannosauridés [21].
- Tyrannosauroidea
- Proceratosauridae
- Pantyrannosauria
Répartition géographique
Les Tyrannosauroidea vivaient sur le super-continent de Laurasie, qui s'est séparé du Gondwana au Jurassique moyen, ainsi que sur les continents de l'hémisphère nord, qui se sont séparés de la Laurasie plus tard au cours de l'ère mésozoïque. Les plus anciens Tyrannosauroidea connus vivaient au Jurassique moyen, il y a environ 168—166 millions d'années, et sont représentés par les genres Proceratosaurus du Bathonien d'Angleterre[22] et son proche parent Kileskus, découvert dans des strates du même âge en Russie[23]. D'autres genres ont vécu au Jurassique supérieur, tels Guanlong du Nord-ouest de la Chine[3], Stokesosaurus de l'Ouest des États-Unis et Aviatyrannis du Portugal. Certains fossiles actuellement attribués à Stokesosaurus pourraient appartenir au genre Aviatyrannis, étant donné les grandes similitudes dans les faunes de dinosaures du Portugal et d'Amérique du Nord au cours de cette période. Si Iliosuchus, du Jurassique moyen d'Angleterre, est en fait un membre des Tyrannosauroidea, il serait le premier connu et pourrait suggérer que la super-famille est originaire d'Europe[9].
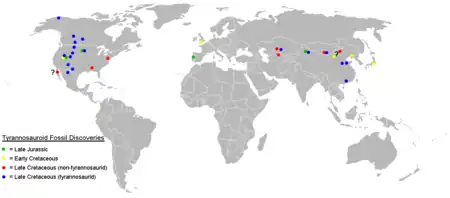
Les Tyrannosauroidea du Crétacé inférieur ont été découverts sur les trois continents de l'hémisphère nord. Eotyrannus en Angleterre[6] et Dilong dans le nord de la Chine[3] sont les deux seuls genres nommés de cette époque, et des dents prémaxillaires du début du Crétacé ont été recueillies dans la Formation de Cedar Mountain dans l'Utah[24] et dans le Groupe de Tetori au Japon[25].
Vers le milieu du Crétacé, les fossiles de Tyrannosauroidea ne se trouvent plus en Europe, suggérant une extinction localisée sur ce continent[1]. Des dents de Tyrannosauroidea et de possibles corps fossiles sont connus des Grès du Dakota en Amérique du Nord[24], ainsi que dans des formations géologiques du Kazakhstan, du Tadjikistan et d'Ouzbékistan[26] à partir du Crétacé moyen.
Les premiers restes incontestables de tyrannosauridés proviennent du Campanien (Crétacé supérieur) d'Amérique du Nord et d'Asie. Deux sous-familles sont reconnues. Les albertosaurinés ne sont connus qu'en Amérique du Nord, tandis que les tyrannosaurinés se trouvent sur les deux continents[1]. Des fossiles de tyrannosauridés ont été trouvés en Alaska, lequel pourrait avoir servi de pont terrestre permettant leur dispersion entre les deux continents[27]. Des Tyrannosauroidea non tyrannosauridés comme Alectrosaurus et éventuellement Bagaraatan étaient contemporains de tyrannosauridés en Asie, alors qu'ils sont absents de l'ouest de l'Amérique du Nord[1]. Au Crétacé supérieur, la voie maritime intérieure de l'Ouest divisait en deux l'Amérique du Nord et isolait la partie occidentale du continent appelée Laramidia. L'absence de tyrannosauridés dans la partie orientale du continent — Appalachia — suggère que la famille a évolué après l'apparition de la mer intérieure nord-américaine, permettant aux membres basaux des Tyrannosauroidea, tels que Dryptosaurus et Appalachiosaurus, de survivre dans l'est comme une population relicte jusqu'à la fin du Crétacé[7].
Des représentants basaux des Tyrannosauroidea pourraient aussi avoir été présents dans ce qui est maintenant le sud-est de l'Australie au cours de l'Aptien au début du Crétacé. Le spécimen NMV P186046, constitué d'un pubis partiel (os de la hanche) d'une forme retrouvée chez les Tyrannosauroidea, a été découvert à Dinosaur Cove dans l'état de Victoria, indiquant que les Tyrannosauroidea ne seraient pas limités aux continents septentrionaux[28]. Cette hypothèse a été contestée par d'autres auteurs, indiquant que la forme du pubis n'est pas propre aux Tyrannosauroidea et que le spécimen pourrait être un membre du groupe plus large des coelurosauriens ou d'un autre clade de théropodes[29].
Si l'hypothèse que les Megaraptora soient des Tyrannosauroidea est averée, ceux-ci auraient un représentant provenant de l'Albien d'Australie, à savoir Australovenator, découvert dans la Formation de Winton, datée d'environ 95 millions d'années[30].
Paléobiologie
Plumes
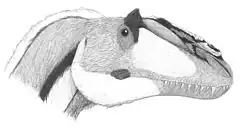
De longues structures filamenteuses ont été préservées avec des restes de squelettes de nombreux coelurosauriens provenant de la formation d'Yixian du Crétacé inférieur et d'autres formations géologiques du Liaoning, en Chine[31]. Ces filaments ont généralement été interprétés comme des « protoplumes », homologues aux plumes ramifiées rencontrées chez les oiseaux et certains théropodes non-aviaires[32] - [33], bien que d'autres hypothèses aient été proposées[34]. Un squelette de Dilong, décrit en 2004, fut le premier exemple de plumes trouvées chez les Tyrannosauroidea. Semblables au duvet des oiseaux modernes, les plumes rencontrées pour Dilong sont ramifiées mais non pennées, et pourraient avoir servi de protection thermique[2] - [35]. Même les grands Tyrannosauroidea ont été trouvés avec des indices de plumes. Yutyrannus huali, également de la formation d'Yixian, est connu à partir de trois spécimens, chacun conservant des traces de plumes sur diverses parties du corps. Bien que toutes les zones du corps ne présentent pas d'empreintes de plumes chez ces trois individus, ces fossiles montrent que, même pour cette espèce de taille importante (9 mètres de long), la majeure partie du corps était couverte de plumes[36].
La présence de plumes, attestée chez certains Tyrannosauroidea primitifs, est également une caractéristique des coelurosauriens, dans d'autres genres basaux comme Sinosauropteryx[32], ainsi que chez tous les groupes plus dérivés[31]. De rares empreintes de peau fossilisées retrouvées chez certains tyrannosauridés sans plume du Crétacé supérieur, cependant, montrent soit une peau nue ou une peau recouverte d'écailles[37]. Il est possible que les plumes étaient présentes sur des zones du corps là où les empreintes de la peau n'étaient pas conservées, car ces zones sont très petites et apparaissent principalement sur les jambes et sur le dessous de la queue. Sinon, la perte secondaire des plumes chez les grands tyrannosauridés pourrait être analogue à la perte des poils chez les plus grands mammifères modernes comme les éléphants, où un faible rapport surface-volume ralentit le transfert thermique, ce qui rend l'isolation par une couche de poils superflue[2].
Crêtes crâniennes
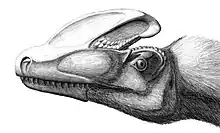
Des crêtes osseuses sont présentes sur les crânes de beaucoup de théropodes, dont de nombreux Tyrannosauroidea. La plus élaborée se trouve chez Guanlong, où les os du nez supportent une unique et grande crête qui longe la ligne médiane du crâne d'avant en arrière. Cette crête est perforée par plusieurs grands foramina (trous) qui réduisent son poids[3]. Une crête moins importante se retrouve chez Dilong, où des petites arêtes parallèles courent de chaque côté du crâne, soutenues par les os du nez et les os lacrymaux. Ces arêtes se courbent vers l'intérieur et se rejoignent juste derrière les narines, donnant à la crête une forme de Y[2]. Les naseaux fusionnés des tyrannosauridés sont souvent très rugueux. Alioramus, genre possible de tyrannosauridé de Mongolie, porte une unique rangée de cinq bosses osseuses importantes sur les os du nez ; une rangée similaire de bosses beaucoup plus petites est présente sur le crâne d'Appalachiosaurus, ainsi que sur quelques spécimens de Daspletosaurus, d'Albertosaurus, et de Tarbosaurus[7]. Chez Albertosaurus, Gorgosaurus et Daspletosaurus, une corne proéminente est présente en face de chaque œil sur l'os lacrymal. Cette corne lacrymale est absente chez Tarbosaurus et Tyrannosaurus, qui ont à la place une crête en forme de croissant derrière chaque œil sur l'os postorbitaire[1].
Ces crêtes crâniennes pourraient avoir été utilisés pour la reconnaissance des individus ou la parade nuptiale[1]. Un exemple du principe de handicap pourrait être le cas pour Guanlong, où la grande crête délicate pourrait avoir été un obstacle à la chasse pour ce probable prédateur actif. Si un individu était en bonne santé et performant à la chasse malgré cette crête fragile, cela pourrait indiquer la supériorité de celui-ci sur les autres avec de petites crêtes. De même que la queue d'un paon mâle ou les bois hors normes d'un Megaloceros, la crête de Guanlong pourrait avoir évolué par sélection sexuelle, en fournissant un avantage lors de la parade nuptiale, l'emportant sur toute diminution de la capacité à chasser[3].
Voir aussi
Notes et références
Notes
Références taxonomiques
- (en) Référence Paleobiology Database : †superfamily Tyrannosauroidea Walker 1964 (coelurosaur) (consulté le )
Références
- (en) Thomas R. Holtz, « Tyrannosauroidea », dans David B. Weishampel, Peter Dodson et Halszka, The Dinosauria (Second ed.), Berkeley, University of California Press, (ISBN 0-520-24209-2), p. 111–136.
- (en) Xing X. Xu, Mark A. Norell, Xuewen Kuang, Xiaolin Wang, Qi Zhao et Chengkai Jia, « Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids », Nature, vol. 431, no 7009, , p. 680–684 (PMID 15470426, DOI 10.1038/nature02855).
- (en) Xing X. Xu, James M. Clark, Catherine A. Forster, Mark A. Norell, Gregory M. Erickson, David A. Eberth, Chengkai Jia et Qi Zhao, « A basal tyrannosauroid dinosaur from the Late Jurassic of China », Nature, vol. 439, no 7077, , p. 715–718 (PMID 16467836, DOI 10.1038/nature04511).
- (en) « Tyrannosaurus ancestor's teeth found in Hyogo », sur The Japan Times,
- (en) S.L. Brusatte, Mark A. Norell, Thomas D. Carr, Gregory M. Erickson, John R. Hutchinson, Amy M. Balanoff, Gabe S. Bever, Jonah N. Choiniere, Peter J. Makovicky et Xing Xu, « Tyrannosaur paleobiology: new research on ancient exemplar organisms », Science, vol. 329, no 5998, , p. 1481−1485 (PMID 20847260, DOI 10.1126/science.1193304, Bibcode 2010Sci...329.1481B).
- (en) Stephen Hutt, Darren Naish, David M. Martill, Michael J. Barker et Penny Newberry, « A preliminary account of a new tyrannosauroid theropod from the Wessex Formation (Early Cretaceous) of southern England », Cretaceous Research, vol. 22, no 2, , p. 227–242 (DOI 10.1006/cres.2001.0252).
- (en) Thomas D. Carr, Thomas E. Williamson et David R. Schwimmer, « A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama », Journal of Vertebrate Paleontology, vol. 25, no 1, , p. 119–143 (DOI 10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.)
- (en) Elizibeth D. Quinlan, Kraig Derstler et Mercedes M. Miller Mercedes M. « Anatomy and function of digit III of the Tyrannosaurus rex manus » () (lire en ligne, consulté le )
—Geological Society of America Annual Meeting - (en) Oliver W. M. Rauhut, « A tyrannosauroid dinosaur from the Upper Jurassic of Portugal », Palaeontology, vol. 46, no 5, , p. 903–910 (DOI 10.1111/1475-4983.00325).
- (en) Henry F. Osborn, « Tyrannosaurus and other Cretaceous carnivorous dinosaurs », Bulletin of the American Museum of Natural History, vol. 21, no 3, , p. 259–265 (DOI 10.1111/j.1468-5965.2007.00735_17.x)
- (en) Alick D. Walker, « Triassic reptiles from the Elgin area: Ornithosuchus and the origin of carnosaurs », Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, vol. 248, no 744, , p. 53–134 (Bibcode 1964RSPTB.248...53W).
- (en) José F. Bonaparte, Fernando E. Novas et Rodolfo A. Coria, « Carnotaurus sastrei Bonaparte, the horned, lightly built carnosaur from the Middle Cretaceous of Patagonia », Contributions in Science (Natural History Museum of Los Angeles County), vol. 416, , p. 1–42.
- (en) Paul C. Sereno, « A rationale for phylogenetic definitions, with application to the higher-level taxonomy of Dinosauria », Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, vol. 210, no 1, , p. 41–83.
- (en) Paul C. Sereno, « Archosauria: Avemetatarsalia to Neornithes » [archive du ], sur http://www.taxonsearch.org/, (consulté le ).
- (en) F. E. Novas, F. L. Agnolín, M. D. Ezcurra, J. I. Canale et J. D. Porfiri, « Megaraptorans as members of an unexpected evolutionary radiation of tyrant-reptiles in Gondwana », Ameghiniana, vol. 49, no Suppl., , R33 (lire en ligne).
- (en) Juan D. Porfiri, Fernando E. Novas, Jorge O. Calvo, Federico L. Agnolín, Martín D. Ezcurra et Ignacio A. Cerda, « Juvenile specimen of Megaraptor (Dinosauria, Theropoda) sheds light about tyrannosauroid radiation », Cretaceous Research, vol. 51, , p. 35–55 (DOI 10.1016/j.cretres.2014.04.007).
- (en) C. Yun, « Comments on the juvenile Megaraptor specimen and systematic positions of megaraptoran theropods », PeerJ PrePrints, vol. 3, , e1051 (DOI 10.7287/peerj.preprints.851v1).
- (en) P. R. Bell, A. Cau, F. Fanti et E. Smith, « A large-clawed theropod (Dinosauria: Tetanurae) from the Lower Cretaceous of Australia and the Gondwanan origin of megaraptorid theropods », Gondwana Research, (DOI 10.1016/j.gr.2015.08.004).
- (en) M.A. Loewen, R.B. Irmis, J.J.W. Sertich, P. J. Currie et S. D. Sampson, « Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous Oceans », PLoS ONE, vol. 8, no 11, , e79420 (DOI 10.1371/journal.pone.0079420).
- (en) Stephen Brusatte et Thomas D. Carr, « The phylogeny and evolutionary history of tyrannosauroid dinosaurs », Scientific Reports, no 20252, (DOI 10.1038/srep20252)
- (en) Rafael Delcourt et Orlando Nelson Grillo, « Tyrannosauroids from the Southern Hemisphere: Implications for biogeography, evolution, and taxonomy », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 511, , p. 379–387 (DOI https://doi.org/10.1016/j.palaeo.2018.09.003, lire en ligne)
- (en) O. W. M. Rauhut, A.C. Milner et S. Moore-Fay, « "Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England », Zoological Journal of the Linnean Society, vol. 158, no 1, , p. 155-195 (DOI 10.1111/j.1096-3642.2009.00591.x)
- (en) Alexander Olegovich Averianov, Sergei A. Krasnolutskii et S.V. Ivantsov, « A new basal coelurosaur (Dinosauria: Theropoda) from the Middle Jurassic of Siberia », Proceedings of the Zoological Institute, vol. 314, no 1, , p. 42–57 (lire en ligne [PDF]).
- (en) James I. Kirkland, Brooks Britt, Donald L. Burge, Kenneth Carpenter, Richard Cifelli, Frank DeCourten, Jeffrey Eaton, Steven Hasiotis et Timothy Lawton, « Lower to Middle Cretaceous Dinosaur faunas of the central Colorado Plateau: a key to understanding 35 million years of tectonics, sedimentology, evolution, and biogeography », Brigham Young University Geology Studies, vol. 42, no II, , p. 69–103.
- (en) Makoto Manabe, « The early evolution of the Tyrannosauridae in Asia », Journal of Paleontology, vol. 73, no 6, , p. 1176–1178.
- (en) Lev A. Nesov (trad. Tatayana Platonova), Dinosaurs of Northern Eurasia: new data about assemblages, ecology and paleobiogeography, St. Petersburg, Scientific Research Institute of the Earth's Crust, St. Petersburg State University, , 156 p.
- (en) Anthony R. Fiorillo et Roland A. Gangloff, « Theropod teeth from the Prince Creek Formation (Cretaceous) of northern Alaska, with speculations on Arctic dinosaur paleoecology », Journal of Vertebrate Paleontology, vol. 20, no 4, , p. 675–682 (DOI 10.1671/0272-4634(2000)020[0675:TTFTPC]2.0.CO;2).
- (en) R. B. J. Benson, P. M. Barrett, T. H. Rich et P. Vickers-Rich, « A southern tyrant reptile », Science, vol. 327, no 5973, , p. 1613 (PMID 20339066, DOI 10.1126/science.1187456, Bibcode 2010Sci...327.1613B).
- (en) Matthew C. Herne, Jay P. Nair et Steven W. Salisbury, « Comment on "A southern tyrant reptile" », Science, vol. 329, no 5995, , p. 1013 (DOI 10.1126/science.1190100).
- (en) Scott A. Hocknull, White, Matt A., Travis R. Tischler, Alex G. Cook, Naomi D. Calleja, Trish Sloan et David A Elliott, « New Mid-Cretaceous (Latest Albian) Dinosaurs from Winton, Queensland, Australia », PLoS ONE, vol. 4, no 7, , e6190 (DOI 10.1371/journal.pone.0006190).
- (en) Zhonghe Z. Zhou, Paul M. Barrett et Jason Hilton, « An exceptionally preserved Lower Cretaceous ecosystem », Nature, vol. 421, no 6925, , p. 807–814 (PMID 12594504, DOI 10.1038/nature01420).
- (en) Pei-ji Chen, Zhiming Dong et Shuonan Zhen, « An exceptionally well-preserved theropod dinosaur from the Yixian Formation of China », Nature, vol. 391, no 6663, , p. 147–152 (DOI 10.1038/34356, Bibcode 1998Natur.391..147C).
- (en) Xing X. Xu, Zhonghe Zhou et Richard A. Prum, « Branched integumental structures in Sinornithosaurus and the origin of feathers », Nature, vol. 410, no 6825, , p. 200–204 (PMID 11242078, DOI 10.1038/35065589).
- (en) Theagarten Lingham-Soliar, Alan Feduccia et Xiaolin Wang, « A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres », Proceedings of the Royal Society of London. Series B, Biological Sciences, vol. 274, no 1620, , p. 1823–1829 (PMID 17521978, PMCID 2270928, DOI 10.1098/rspb.2007.0352).
- Cécile Dumas, « Dragon à plumes », sur Sciences et avenir, (consulté le ).
- (en) X. Xu, K. Wang, K. Zhang, Q. Ma, L. Xing, C. Sullivan, D. Hu, S. Cheng et S. Wang, « A gigantic feathered dinosaur from the Lower Cretaceous of China », Nature, vol. 484, no 7392, , p. 92–95 (PMID 22481363, DOI 10.1038/nature10906, Bibcode 2012Natur.484...92X).
- (en) Larry D. Martin et Stephan A. Czerkas, « The fossil record of feather evolution in the Mesozoic », American Zoologist, vol. 40, no 4, , p. 687–694 (DOI 10.1668/0003-1569(2000)040[0687:TFROFE]2.0.CO;2).