Proteus (bactérie)
Proteus est un genre de bacilles Gram négatifs (BGN) de la famille des Morganellaceae, proche des genres Morganella et Providencia avec lesquels il forme la tribu des Proteeae. Son nom provient de celui du dieu grec Protée, doué de métamorphose, en référence à l'aspect particulièrement polymorphe de ces bactéries[1].
| Règne | Bacteria |
|---|---|
| Embranchement | Pseudomonadota |
| Classe | Gammaproteobacteria |
| Ordre | Enterobacterales |
| Famille | Morganellaceae |
| Tribu | Proteeae |
Plusieurs espèces de Proteus sont des pathogènes humains opportunistes. L'espèce type du genre, Proteus mirabilis, possède de multiples facteurs de virulence et cause le plus souvent des infections urinaires connues pour leur tendance à se compliquer de calculs[2].
Caractéristiques
Habitat
Les espèces P. mirabilis et P. vulgaris sont les plus répandues. Ce sont des commensaux habituels du tube digestif humain et de divers autres mammifères tels que chiens, chats, rats, singes, cochons, moutons, bétail, ratons laveurs etc.[3] Elles disposent également d'un vaste réservoir environnemental puisqu'on les retrouve dans les eaux, les sols, les égouts et les matières fécales.
L'espèce P. penneri semble moins courante. D'après une étude de 1986 c'est la seule des espèces alors connues de la tribu des Proteeae à n'avoir pas été détectée dans l'environnement (litière, lait) de bovins d'élevage[4].
Morphologie et culture

Ce sont des bactéries mobiles par ciliature péritriche c'est-à-dire qu'elles se déplacent à l'aide de flagelles implantés tout autour de la cellule. Comme tous les Enterobacterales leur type respiratoire est anaérobie facultatif. Elle ont la particularité de changer d'aspect en fonction de la consistance du milieu dans lequel elles évoluent. En milieu liquide ce sont de courts bacilles (swimmer cells) de 1,5 à 2 µm de long munies de 4 à 10 flagelles, mais en milieu solide elles adoptent un autre phénotype (swarmer cell) caractérisé par une importante élongation (de 10 à > 80 µm de long) et une augmentation considérable du nombre de flagelles (1 000 à 10 000 par cellule)[3].
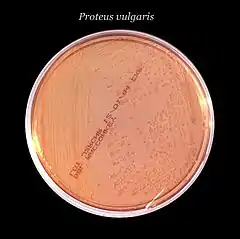
Cette deuxième forme dispose d'une propriété remarquable appelée « essaimage » (en anglais swarming)[5] - [6] - [7]. Il s'agit d'un type particulier de mobilité, cyclique et coordonnée, qui donne aux colonies un aspect typique en « œil de bœuf » avec des cercles concentriques qui correspondent aux vagues de migration successives à partir du point d'inoculation. L'essaimage n'est pas spécifique aux Proteus, il a également été décrit chez plusieurs autres genres bactériens tels que Aeromonas[8], Bacillus[9], Escherichia[10] - [11], Pseudomonas[12] - [13], Salmonella[10], Serratia[14] - [15], Vibrio[16] ou encore Yersinia[17].
Les Proteus sont des bactéries sans exigence nutritionnelle particulière ce qui signifie qu'elles peuvent croître sur la plupart des milieux de base, sans nécessiter ni facteurs de croissance ni conditions de culture spécifiques. En pratique on peut les cultiver sur les géloses non sélectives (GNO, GTS etc.) ou sur les géloses « entériques » utilisées pour l'isolement des BGN (MacConkey, EMB etc). Pour inhiber ou du moins limiter le phénomène d'essaimage, qui peut gêner l'isolement des colonies, il est recommandé de modifier la composition des milieux de culture en augmentant la concentration d'agar jusqu'à 2% (m/v) et ajoutant 5 mL de glycérol par litre de milieu[3].
Caractères biochimiques
Les Proteus sont des bactéries oxydase négatives et catalase positives comme la plupart des Enterobacterales. Elles partagent avec les autres bactéries de la famille des Morganellaceae l'absence d'arginine dihydrolase (ou arginine décarboxylase) et un résultat négatif à la réaction de Voges-Proskauer.
Avec les genres Morganella et Providencia (tribu des Proteeae) elles partagent de plus les caractéristiques suivantes[3] - [18] :
- présence d'une phénylalanine désaminase, d'une tryptophane désaminase et d'une ornithine désaminase ;
- absence de lysine décarboxylase ;
- absence de β-galactosidase (test à l'ONPG négatif) ;
- réduction des nitrates en nitrites ;
- acidification du D-glucose mais pas du mélibiose.
En outre les Proteus ont en commun avec les Morganella l'activité uréase ainsi que l'incapacité à acidifier le D-adonitol, le D-arabitol, le D-mannitol et le myo-inositol. Cependant ils se distinguent à la fois des Morganella et des Providencia par l'incapacité à acidifier le D-mannose.
Le tableau suivant indique d'autres caractères, variables au sein du genre, qui peuvent contribuer au diagnostic d'espèce par la méthode phénotypique[18] - [19] :
| production indole | production H2S | activité ornithine décarboxylase | activité lipase | hydrolyse gélatine | hydrolyse esculine | acidification maltose | acidification tréhalose | acidification salicine | |
|---|---|---|---|---|---|---|---|---|---|
| P. mirabilis | ― | + | + | + | + | ― | ― | + | ― |
| P. vulgaris | + | ± | ― | ― | ± | + | + | ― | + |
| P. penneri | ― | ± | ― | ± | ± | ― | + | ± | ― |
| P. hauseri | + | ± | ― | + | + | ― | + | + | ― |
Pathogénicité humaine
Tableaux cliniques
Les infections urinaires à P. mirabilis constituent la forme clinique la plus fréquente des infections humaines à Proteus[19] mais beaucoup d'autres localisations ont été décrites avec des prédominances variables selon les espèces. De rares cas d'endocardite infectieuse infectieuse ont été rapportés[20]. Leur rôle pathogène est discuté dans divers tableaux infectieux digestifs[21].
P. mirabilis est le plus souvent responsable d'infections urinaires hautes (pyélonéphrites) ou basses (cystites) dont il ne représente cependant que 1 à 10% du total, la première cause étant de loin E. coli. Ces infections surviennent volontiers sur un terrain à risque de complications, par exemple en cas de vessie neurologique ou de malformation des voies urinaires, et se compliquent fréquemment de lithiase urinaire[2] - [18]. P. mirabilis est particulièrement impliqué dans les infections urinaires sur sonde à demeure dont il cause 10 à 44% des cas, avec un pic d'incidence chez les personnes âgées[2]. C'est aussi l'agent de colonisations urinaires se traduisant par une bactériurie asymptomatique chez des personnes âgées diabétiques[2]. En dehors de l'appareil urinaire P. mirabilis peut causer des bactériémies[22] - [23] - [24], des pneumonies[25], des infections cutanées, en particulier des surinfections de brûlures[26] et d'autres plaies[27], ainsi que des infections oculaires graves[28] - [29]. Des cas sporadiques de méningoencéphalite néonatale[30] - [31], d'empyème pleural[32], d'ostéomyélite sur drépanocytose[33] et d'abcès rénaux[34] ont été décrits.
Les autres espèces sont plus souvent retrouvées dans les localisations extra-urinaires telles que les infections des parties molles et les surinfections de plaies[19]. P. vulgaris a été ponctuellement impliqué dans des cas d'empyème pleural[35] et d'endophtalmie[36].
Certains travaux suggèrent que les infections urinaires à Proteus (en particulier P. mirabilis) pourraient être impliquées dans la physiopathologie de la polyarthrite rhumatoïde[37] - [38]. D'autres études rapportent une association entre la présence de Proteus dans le tube digestif et la survenue de la maladie de Crohn[39].
Facteurs de virulence
Ils sont particulièrement bien étudiés chez P. mirabilis[40] - [41], en particulier dans le cas des infections urinaires sur sonde à demeure[2] - [42].
Uréase
_-_Str%C3%BCvit_kristalleri_(idrar)_-_01.png.webp)
Les uréases sont des métalloenzymes cytoplasmiques dont le site actif comporte des atomes de nickel[42]. Elles catalysent l'hydrolyse de l'urée en ammoniac et en dioxyde de carbone selon l'équation :
L'ammoniac produit est un nutriment azoté directement assimilable par les bactéries. En cas d'infection urinaire à P. mirabilis l'ammoniac dissous dans l'urine provoque son alcalinisation, c'est-à-dire l'élévation de son pH, car c'est une petite molécule très soluble dans l'eau et une base faible. Ces conditions sont favorables à la précipitation de deux composés dissous dans l'urine : le phosphate d'ammonium et de magnésium (NH3MgPO4) qui cristallise sous la forme d'un minéral appelé struvite, et le phosphate de calcium (Ca3(PO4)2) qui forme des cristaux d'hydroxyapatite[43]. C'est ainsi que les infections urinaires à P. mirabilis favorisent la formation de calculs qui évoluent ensuite pour leur propre compte. L'une des complications les plus graves est le blocage des urines infectées en amont d'un calcul enclavé dans la voie urinaire (pyélonéphrite aiguë obstructive ou « sur obstacle ») car sans traitement rapide, l'évolution se fait spontanément vers un choc septique pouvant engager le pronostic vital. Sur le long terme, l'obstruction urinaire prolongée peut engendrer une néphropathie tubulo-interstitielle qui évolue lentement vers l'insuffisance rénale chronique.
Lorsqu'une sonde urinaire est placée dans l'urètre, les microcristaux de struvite et d'apatite participent à la formation d'un biofilm qui se dépose et adhère à sa surface interne.
Essaimage
Sensibilité aux antibiotiques
P. mirabilis est intrinsèquement résistant à la nitrofurantoïne et aux tétracyclines[18] ainsi qu'aux polymyxines (colistine et apparentés)[44]. Les souches sauvages sont habituellement sensibles aux amino- et uréidopénicillines, à des céphalosporines de diverses générations (céfazoline, céfoxitine, céfuroxime, céfotaxime, ceftriaxone, ceftizoxime, ceftazidime, céfépime), à l'imipénème, à plusieurs aminosides (amikacine, gentamicine, tobramycine), à la ciprofloxacine et au cotrimoxazole[18].
Résistance naturelle
Souches sauvages sensibles à toutes les bêta-lactamines (pas de céphalosporinase chromosomique de classe C, exemple : Proteus mirabilis 1761). P. vulgaris possède quant à lui une céfuroximase à l'état sauvage.
Résistance acquise
Mécanismes identiques à ceux décrits pour E. coli. Résistances constatées :
- 1. beta-lactamase de classe A haut niveau (pénicillinase) : des carbénicillinases (type PSE-4) ont été décrites.
- 2. résistance aux inhibiteurs des beta-lactamases : un mutant de type TEM, IRT-2 (Arg244Ser), a été décrit en association avec TEM-1 dans une souche insensible aux inhibiteurs des beta-lactamases de classe A.
- 3. beta-lactamases de classe A à spectre étendu (BLSE) : une souche produisant TEM-10 a été décrite.
- 4. résistance à l'imipénème : chez P. mirabilis, elle n'est pas d'origine enzymatique. La résistance à l'imipénème semble associée à une altération des PLP1A et 2 (protéines liant la pénicilline).
Liste d'espèces
Selon la LPSN[1], en juin 2021, les espèces suivantes sont recensées :
- Proteus alimentorum Dai et al. 2018
- Proteus cibi Dai et al. 2019
- Proteus columbae Dai et al. 2018
- Proteus faecis Dai et al. 2019
- Proteus hauseri O'Hara et al. 2000
- Proteus mirabilis Hauser 1885 – espèce type
- Proteus penneri Hickman et al. 1983
- Proteus terrae Behrendt et al. 2016
- Proteus terrae subsp. terrae (Behrendt et al. 2016) Behrendt et al. 2021
- Proteus terrae subsp. cibarius (Hyun et al. 2016) Behrendt et al. 2021 : reclassement de P. cibarius
- Proteus vulgaris Hauser 1885
Certaines espèces ont été reclassées dans d'autres genres :
- Cosenzaea myxofaciens (Cosenza & Podgwaite 1966) Giammanco et al. 2011 : reclassement de P. myxofaciens
- Morganella morganii (Winslow et al. 1919) Fulton 1943 : reclassement de P. morganii
- Providencia alcalifaciens (de Salles Gomes 1944) Ewing 1962 : reclassement de P. inconstans
- Providencia rettgeri (Hadley et al. 1918) Brenner et al. 1978 : reclassement de P. rettgeri
Références
- https://lpsn.dsmz.de/genus/proteus, consulté le 11/10/22.
- Schaffer JN & Pearson MM « Proteus mirabilis and urinary tract infections » Microbiol Spectr. 2015;3(5):10.1128/microbiolspec.UTI-0017-2013. Accès libre.
- Manos J & Belas R (2006) The Genera Proteus, Providencia, and Morganella. In: Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackebrandt E (eds) The Prokaryotes. Springer, New York, NY. https://doi.org/10.1007/0-387-30746-X_12
- Hawkey PM et al. « Speciation, serotyping, antimicrobial sensitivity and plasmid content of Proteeae from the environment of calf-rearing units in South West England » J Hyg (Lond). 1986;97(3):405-417. Accès libre.
- Rather PN « Swarmer cell differentiation in Proteus mirabilis » Environ Microbiol. 2005;7(8):1065-1073. https://doi.org/10.1111/j.1462-2920.2005.00806.x
- Morgenstein RM, Szostek B & Rather PN « Regulation of gene expression during swarmer cell differentiation in Proteus mirabilis » FEMS Microbiol Rev. 2010;34(5):753-763. https://doi.org/10.1111/j.1574-6976.2010.00229.x
- Gmiter D & Kaca W « Into the understanding the multicellular lifestyle of Proteus mirabilis on solid surfaces » Front Cell Infect Microbiol.12:864305. https://doi.org/10.3389/fcimb.2022.864305
- Kirov SM et al. « Lateral flagella and swarming motility in Aeromonas species » J Bacteriol. 2002;184(2):547-555. Accès libre.
- Kearns DB & Losick R « Swarming motility in undomesticated Bacillus subtilis » Molec Microbiol. 2003;49(3):581-590. Accès libre
- Harshey RM & Matsuyama T « Dimorphic transition in Escherichia coli and Salmonella typhimurium: surface-induced differentiation into hyperflagellate swarmer cells » PNAS 1994;91(18):8631–8635. Accès libre.
- Burkart M, Toguchi A & Harshey RM « The chemotaxis system, but not chemotaxis, is essential for swarming motility in Escherichia coli » PNAS 1998;95(5):2568–2573. Accès libre.
- Köhler T et al. « Swarming of Pseudomonas aeruginosa is dependent on cell-to-cell signaling and requires flagella and pili » J Bacteriol. 2000;182(21):5990–5996. Accès libre
- Tremblay J et al. « Self-produced extracellular stimuli modulate the Pseudomonas aeruginosa swarming motility behaviour » Environ Microbiol. 2007;9(10):2622-2630. https://doi.org/10.1111/j.1462-2920.2007.01396.x
- Alberti L & Harshey RM « Differentiation of Serratia marcescens 274 into swimmer and swarmer cells » J Bacteriol. 1990;172(8):4322–4328. Accès libre.
- Eberl L, Molin S & Givskov M « Surface motility of Serratia liquefaciens MG1 » 1999;181(6):1703–1712. Accès libre.
- McCarter L & Silverman M « Surface-induced swarmer cell differentiation of Vibrio parahaemoiyticus » Molec Microbiol. 1990;4(7):1057-1062. https://doi.org/10.1111/j.1365-2958.1990.tb00678.x
- Young GM et al. « The Yersinia enterocolitica motility master regulatory operon, flhDC, is required for flagellin production, swimming motility, and swarming motility » J Bacteriol. 1999;181(9): 2823–2833. Accès libre.
- O'Hara CM, Brenner FW & Miller JM « Classification, Identification, and Clinical Significance of Proteus, Providencia, and Morganella » Clin Microbiol Rev. 2000;13(4):534-546. Accès libre.
- Forsythe SJ, Abbott SL & J Pitout J (2015) Klebsiella, Enterobacter, Citrobacter, Cronobacter, Serratia, Plesiomonas, and Other Enterobacteriaceae. In: Jorgensen JH, Pfaller MA (eds) Manual of clinical microbiology. 11th edition. ASM Press, Washington, DC. https://doi.org/10.1128/9781555817381
- Ioannou P & Vougiouklakis G « Infective endocarditis by Proteus species: a systematic review » Germs 2020;10(3):229-239. Accès libre.
- Hamilton AL et al. « Proteus spp. as putative gastrointestinal pathogens » Clin Microbiol Rev. 2018;31(3):e00085-17. Accès libre.
- Adler JL et al. « Proteus bacteremia in a general hospital » Ann Intern Med. 1971;75(4):531-536. https://doi.org/10.7326/0003-4819-75-4-531
- Watanakunakorn C & Perni SC « Proteus mirabilis bacteremia: a review of 176 cases during 1980-1992 » Scandinav J Infect Dis. 1994;26(4):361-367. https://doi.org/10.3109/00365549409008605
- Chen CY et al. « Proteus mirabilis urinary tract infection and bacteremia: risk factors, clinical presentation, and outcomes » J Microbiol Immunol Infect. 2012;45(3):228-36. Accès libre.
- Okimoto N et al. « Clinical features of Proteus mirabilis pneumonia » J Infect Chemother. 2010;16(5):364-366. https://doi.org/10.1007/s10156-010-0059-3
- Davis B, Lilly HA & Lowbury EJ « Gram-negative bacilli in burns » J Clin Pathol. 1969;22(6):634-641. Accès libre.
- Bessa LJ et al. « Bacterial isolates from infected wounds and their antibiotic susceptibility pattern : some remarks about wound infection » Int Wound J. 2015;12(1): 47–52. Accès libre.
- Mo S et al. « Proteus mirabilis keratitis : risk factors, clinical features, treatment outcomes, and microbiological characteristics » Cornea 2021;40(6):704-709. https://doi.org/10.1097/ico.0000000000002469
- Leng T et al. « Endophthalmitis caused by proteus species : antibiotic sensitivities and visual acuity outcomes » Retina 2009;29(7):1019-1024. https://doi.org/10.1097/01.iae.0000357756.22868.2d
- Grahnquist L, Lundberg B & Tullus K « Neonatal Proteus meningoencephalitis. Case report » APMIS 1992;100(8):734-736.
- Coskun Y et al. « A newborn with hemorrhagic meningoencephalitis due to Proteus mirabilis » Drug Discov Ther. 2016;10(6):334-337. https://doi.org/10.5582/ddt.2016.01068
- Pine JR & Hollman JL « Elevated pleural fluid pH in Proteus mirabilis empyema » Chest 1983;84(1):109-111. https://doi.org/10.1378/chest.84.1.109
- Marx AC et al. « Case report 496: Intraosseous gas in Proteus mirabilis osteomyelitis complicating bone infarcts in sickle cell disease » Skeletal Radiol. 1988;17(7):510-3. https://doi.org/10.1007/bf00364047
- Kakuchi Y « Multiple renal abscesses due to Proteus mirabilis » Clin Expl Nephrol. 2021;25(3):322-323. https://doi.org/10.1007/s10157-020-01983-9
- Isenstein D & Honig E « Proteus vulgaris empyema and increased pleural fluid pH » Chest 1990;97(2):511. https://doi.org/10.1378/chest.97.2.511b
- Al-Kharashi AS, Al-Kharshi & Al-Faky YH « Endophthalmitis due to Proteus vulgaris after pars plana vitrectomy with devastating outcome » Middle East Afr J Ophtalmol. 2017;24(2):116-118. https://doi.org/10.4103/meajo.meajo_134_15
- Senior BW et al. « Evidence that patients with rheumatoid arthritis have asymptomatic 'non-significant' Proteus mirabilis bacteriuria more frequently than healthy controls » J Infect. 1999;38(2):99-106. https://doi.org/10.1016/s0163-4453(99)90076-2
- Ebringer A & Rashid T « Rheumatoid arthritis is caused by a Proteus urinary tract infection » APMIS 2014;122(5):363-368. https://doi.org/10.1111/apm.12154
- Zhang J et al. « Elucidation of Proteus mirabilis as a key bacterium in Crohn's disease inflammation » Gastroentrology 2021;160(1):317-330. Accès libre.
- Armbruster CE & Mobley HLT « Merging mythology and morphology : the multifaceted lifestyle of Proteus mirabilis » Nat Rev Microbiol. 2012;10(11):743-754. Accès libre.
- Armbruster CE et al. « Pathogenesis of Proteus mirabilis infection » EcoSal Plus 2018;8(1):10.1128/ecosalplus.ESP-0009-2017. Accès libre.
- Norsworthy AN & Pearson MM « From catheter to kidney stone : the uropathogenic lifestyle of Proteus mirabilis » Trends Microbiol. 2017;25(4):304–315. https://doi.org/10.1016%2Fj.tim.2016.11.015
- Griffith DP, Muscher DM & Itin C « Urease. The primary cause of infection-induced urinary stones » Invest Urol. 1976;13(5):346-50 (pas de DOI).
- Moffatt JH, Harper M & Boyce JD « Mechanisms of polymyxin resistance » Adv Exp Med Biol. 2019;1145:55-71. https://doi.org/10.1007/978-3-030-16373-0_5
Liens externes
- (en) Référence Catalogue of Life : Proteus Hauser, 1885 (consulté le )
- (en) Référence LPSN : Proteus Hauser 1885 (consulté le )
- (en) Référence NCBI : Proteus (taxons inclus) (consulté le )
- (en) Référence World Register of Marine Species : taxon Proteus Hauser, 1885 (+ liste espèces) (consulté le )