Histoire de la paléoanthropologie
La paléoanthropologie est la science qui étudie les fossiles humains ou pré-humains, et l'histoire évolutive de la lignée humaine. Elle est née dans la seconde moitié du XIXe siècle, peu de temps après la préhistoire, qui, en tant que discipline, s'intéresse plutôt aux artéfacts laissés par les hommes préhistoriques, notamment leurs industries lithiques.
L'idée d'évolution des espèces émerge au sein de la science au début du XIXe siècle avec Jean-Baptiste de Lamarck, et s'affirme avec le travail de Charles Darwin, publié en 1859 et 1871, qui suit de peu la découverte en 1856 du premier homme fossile, l'Homme de Néandertal[1].
Avant Darwin

Le premier à comparer les espèces et à établir un classement est Aristote, au IVe siècle av. J.-C. Il en déduit une organisation du vivant tendant à s'élever vers l'homme selon une échelle, la Scala naturæ[1]. Hippocrate dans De natura hominis voit une relation entre l'anatomie et l'influence du milieu[1].
Galien (au IIe siècle) note la ressemblance entre l'anatomie des singes et des hommes[1]. Il incite les médecins à s'entraîner à la dissection sur des singes.
Les cabinets de curiosités qui prolifèrent aux XVIIe et XVIIIe siècles voient naitre les premiers inventaires du vivant et leurs premières classifications dans le but de glorifier l’œuvre du créateur[2].
Dans la lignée de Noël-Antoine Pluche et son célèbre Spectacle de la nature, Carl von Linné établit une grande classification de la nature, Systema naturae. Dans l'édition de 1758, l'homme y est décrit sous le nom d’homo sapiens, membre du groupe des Anthropomorphes comprenant les chimpanzés (Homo troglodytes) et les paresseux[2]. Linné considère que les grands singes sont les plus proches parents de l'homme en fonction des similitudes morphologiques et anatomiques.
Le mot Homo est le nom du genre biologique qui regroupe toutes les espèces humaines. Le terme Homo, humain en latin, dérive d'une racine de l'Indo-européen commun *dʰǵʰm̥mō (« [chose / fils] de la terre »)[3]. Il a été choisi par Carl von Linné, dans sa méthode de classification de la nature, Systema naturae (édition de 1758)[2].
Georges-Louis Leclerc de Buffon critique le travail de Linné et ironise : « Dieu a créé, Linné a classé ». Pour sa part, son œuvre est organisée autour de l'homme, le classant en tant qu'espèce unique. Il introduit la notion de « dégradation des espèces », les espèces les plus nobles, sauf l'homme, pouvant subir des altérations au fil du temps[2].
Jean-Baptiste de Lamarck reprend la notion de dégradation en l'inversant : des espèces plus complexes émergent en fonction des changements d'environnement, selon un processus évolutif[4]. Il publie ses théories « positives » de l'évolution des espèces en 1802, dans Système des animaux puis dans Philosophie zoologique, en 1809. Il introduit les notions d'antériorité, de descendance, de généalogie des espèces et de transformisme[4].
Darwin
« Ainsi, l'Homme descendrait du singe. Pourvu que cela ne soit pas vrai. Mais si cela devait être, prions pour que cela ne se sache pas. »
— Phrase attribuée à la femme de l'évêque de Worcester à la suite du débat entre Huxley et Wilberforce en 1861[5].

Le 1er juillet 1858, sont présentés à la Linnean Society of London, un essai de Alfred Russel Wallace, « On the Tendency of Species to form Varieties », ainsi qu'un essai de Charles Darwin[6]. Ces deux essais présentent la théorie de l'évolution par la sélection naturelle[7].
En 1859, Darwin publie son livre, L'Origine des espèces, dans lequel il explique le mécanisme présidant à l'évolution graduelle des espèces vivantes dans la nature. Sa théorie propose que seuls les individus les mieux adaptés aux difficultés survivent et se reproduisent. C'est la sélection naturelle[7]. Le livre de Darwin n'aborde pas la question de l'évolution humaine, mais l'idée de la filiation entre les humains et les anciens singes est devenue évidente[7].
Wallace pense que seule la sélection naturelle préside à la transformation des espèces. Pour Darwin toutefois, elle ne peut tout expliquer. Il complète sa théorie avec celle de la sélection sexuelle quand il publie La Filiation de l'homme et la sélection liée au sexe, en 1871[8] - [7]. Il y considère que les singes sont les animaux les plus proches de l'homme et conjecture que les origines de la lignée humaine se trouvent en Afrique[9] - [10].
Toutefois, l'idée qu'une espèce d'homme distincte de la nôtre ait existé par le passé et ait disparu fut particulièrement difficile à admettre.
Les premiers débats sur la nature de l'évolution humaine ont surgi entre Thomas Henry Huxley et Richard Owen, en particulier, lors du Débat sur l'hippocampe. Huxley argumente pour l'évolution humaine à partir de singes, et aboutit à la déduction que les « différences entre les races humaines les plus élevées et les plus frustes sont du même ordre de grandeur que celles qui séparent le cerveau humain du cerveau simien »[11]. Il reprend ses conclusions dans son livre, publié en 1863, La Place de l'homme dans la nature.
Premiers fossiles

Un problème majeur à cette époque est le manque de fossiles humains. La plus ancienne découverte de restes fossiles est la Dame rouge de Paviland en 1823[12], identifiée lors de sa découverte comme une femme de l'époque romaine[alpha 1].
En 1830, Philippe-Charles Schmerling exhume les restes de trois individus dans les Grottes Schmerling à Engis (Belgique). S'ils attirent l'attention de Charles Lyell, celui-ci n'est pas convaincu de leur ancienneté[12]. D'autres fossiles sont découverts dans la carrière de Forbes à Gibraltar en 1848[12], mais sont assimilés à des restes d'une créature souffrant de maladie[13]. Tous ces fossiles sont identifiés ultérieurement à des néandertaliens.
En 1856, des ossements humains sont découverts dans une carrière de calcaire de la vallée de Néander, près de Düsseldorf (Allemagne), trois ans avant la publication de l’Origine des espèces. Identifiés par Johann Carl Fuhlrott comme des ossements anciens et primitifs, ils font l'objet d'une vaste polémique, l'idée que d'autres types d'hommes aient pu exister n'étant pas admise[12].
La découverte d'une mandibule dans la Grotte de Naulette (Belgique) en 1866, fournit la preuve de l'existence d'autres espèces d'hommes. L'étude comparée de Paul Broca lui permet d'affirmer que cette mandibule « fournit un argument anatomique aux darwinistes. C'est le premier anneau d'une chaîne qui s'étend de l'homme aux singes »[12].
La présentation à la société d’archéologie de Namur de la découverte de fossiles humains en 1886, à la grotte de Spy (Belgique), a un grand retentissement dans la presse du monde entier. Cette découverte est en effet une preuve pour confirmer l'existence et l'ancienneté d'un type humain de morphologie différente de celle de l'homme actuel.
Le premier squelette de Néandertalien mis au jour en France est « le vieillard » de La Chapelle-aux-Saints en 1908[14]. Découvert par Amédée, Jean et Paul Bouyssonie, le corps repose dans une sépulture[14]. Marcellin Boule accentue les traits simiesques des néandertaliens, les décrivant comme une sorte d'homme des cavernes sauvage et brutal, se déplaçant en traînant les pieds et n'arrivant pas à marcher redressé[12].
Le chaînon manquant

À partir des années 1860, Huxley, Broca et Ernst Haeckel modélisent les principes de l'évolution du singe à l'homme. Nommé anthropopithèque par Gabriel de Mortillet (en reprenant le genre Anthropopithecus établi en 1839 par De Blainville[15]) et Pithécanthrope par Haeckel (Pithecanthropus 1868[15]), la recherche du « chaînon manquant », forme hybride entre l'homme moderne et les grands singes, commence[16].
Pour Haeckel, le gibbon est le singe le plus proche de l'homme. Pour lui, le chainon manquant est une forme intermédiaire entre les deux[16].
Premières découvertes hors d'Europe
Eugène Dubois décrit des fossiles découverts en 1891 à Trinil au bord du fleuve Solo (12 km à l'ouest de Ngawi, en Java oriental) qu'il nomme d'abord (en 1892-1893) Anthropopithecus javanensis : le singe-homme de Java. La découverte d'un fémur, démontrant que l'être devait se tenir debout et marcher, l'incite à considérer qu'il s'agissait du « chainon manquant » entre le singe et l'homme. Il le rebaptise (en 1893-1894) Pithecanthropus erectus, l'homme-singe debout[16].
Dans les années 1920, on découvre ce que l'on croit être au départ les premiers néandertaliens connus hors d'Europe :
- 1921 : l'Homme de Kabwe (Homo rhodesiensis), en Zambie
- 1925 : l'Homme de Galilée (Homo heidelbergensis ou Homo rhodesiensis), en Israël

En 1925, Raymond Dart, anthropologue de l'université du Witwatersrand, chercheur convaincu par la lecture de Darwin de l'unique berceau africain de l'espèce humaine, décrit son « chainon manquant » dans la revue Nature, l’Australopithecus africanus[17]. Le spécimen type ainsi décrit est l'enfant de Taung, un crâne et un endocrâne bien conservés d'un individu juvénile, découverts en 1924 dans une carrière de la Northern Lime Company à Taung, près de Kimberley en Afrique du Sud. Bien que le cerveau soit petit (410 cm3), sa forme arrondie, contrairement à celui des chimpanzés et des gorilles, possède une organisation comparable à celle d'un cerveau humain moderne. En outre, le trou occipital, au centre du crâne et orienté vers le bas, atteste d'une locomotion bipède[18]. Tous ces traits ont convaincu Dart que l'enfant de Taung était « une race éteinte de singes, intermédiaires entre les anthropoïdes actuels et l'homme »[19] - [20].
La fraude de Piltdown
En février 1912, Charles Dawson informe Arthur Smith Woodward, président de la Société de géologie de Londres et conservateur du département d'histoire naturelle au Muséum d'histoire naturelle de Grande-Bretagne, qu'il avait trouvé, entre 1908 et 1911, des fragments d'ossements et des outils. Le crâne, mi-homme, mi singe, correspond au chaînon manquant idéal qu'attendent les anthropologues[21].
Toutefois, cet Homme de Piltdown se révèle être une supercherie composée d'un crâne d'homme et d'une mandibule d'orang-outan. Cette supercherie est définitivement établie en 1953 grâce à des analyses physico-chimiques[21].
Les auteurs de la fraude n'ont jamais été découverts, mais seul un spécialiste en paléontologie pouvait donner aux os une patine comparable à celle des autres fossiles trouvés sur le même site[21].
Cap vers l'Asie
Découverte de fossiles d'Homo erectus à Zhoukoudian (Chine, 1921-1937), Ngandong (Java, 1931-1933), et Sangiran (Java, 1936-1941).
Afrique du Sud
Découverte de 1936 à 1949 de plusieurs sites karstiques au Gauteng, en Afrique du Sud, ayant livré des fossiles d'Australopithecus africanus (2,5 à 2,3 Ma) et de Paranthropus robustus (2 à 1,5 Ma). Après le genre Australopithèque en 1925, c'est le genre Paranthrope qui est créé par Robert Broom en 1938.
La ruée vers l'os (Afrique de l'Est)
_Mary_Douglas_Nicol_Leakey_(1913-1996)_and_her_husband_Louis_Seymour_Bazett_Leakey_(1903-1972)_(2).jpg.webp)
| 1850 |  |
| 1900 |  |
| 1950 | 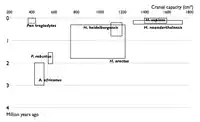 |
| 2002 | 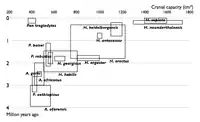 |
Des années 1960 aux années 1980, des centaines de fossiles ont été trouvés en Afrique orientale, dans les régions des gorges d'Olduvai (Tanzanie), du lac Turkana (Kenya), et de la vallée de l'Awash (Éthiopie).
Les membres de la famille Leakey, Louis Leakey et son épouse Mary Leakey, et plus tard leur fils Richard et leur belle-fille Meave ont été parmi les plus actifs paléoanthropologues dans les recherches en Afrique orientale. Après plus de trente ans à fouiller et à découvrir des milliers d'outils préhistoriques, Mary et Louis découvrent, le 17 juillet 1959, leur « dear boy », appelé également Mister Zinj, un représentant de l'espèce Paranthropus boisei. Appliquant une technique de datation absolue, nouvelle pour l'époque, ils repoussent à 1,8 million d'années la naissance de la lignée humaine, estimée jusqu'alors à quelques centaines de milliers d'années[22]. Homo habilis est découvert l'année suivante, en 1960, et publié en 1964.
Dans les années 1970, l'Éthiopie apparait comme le lieu phare de la paléoanthropologie avec la découverte de Lucy, le 30 novembre 1974. C'est un fossile complet à 40 % de l'espèce Australopithecus afarensis, démontrant que l’acquisition de la marche bipède date d'au moins 3,2 millions d’années[23].
En 1976, Mary Leakey fouille la région de Laetoli en Tanzanie, qui livre des empreintes de pas d'hominines bipèdes, datées de 3,7 millions d'années, exceptionnellement conservées dans de la cendre volcanique indurée[24].
En 1984, Kamoya Kimeu, membre de l'équipe de Richard Leakey, découvre à l'ouest du lac Turkana, au Kenya, le squelette exceptionnellement complet du garçon de Turkana, daté de 1,55 million d'années, représentant juvénile de l'espèce Homo ergaster. Le squelette post-crânien apparait très proche de celui de l'Homme moderne, contrairement aux espèces précédentes Homo habilis et Homo rudolfensis. Le crâne conserve en revanche des caractères archaïques.
Ces découvertes et bien d'autres ont permis d'affirmer le rôle de l'Afrique comme berceau du genre Homo.
Les diagrammes ci-contre montrent l'augmentation du nombre d'espèces connues d'Hominina depuis 1850. Chaque rectangle représente la fourchette de dates et de volumes crâniens estimés pour chaque espèce connue à la date considérée. Le rythme de découverte de nouveaux taxons continue de s'accélérer au XXIe siècle.
East Side Story
La découverte de ces nombreux fossiles permet d'appuyer la théorie d'une origine est-africaine de la lignée humaine. Yves Coppens propose en 1982 le modèle de l'East Side Story lors d'une réunion scientifique organisée par l'Académie pontificale des sciences de Rome[25]. La formation de la vallée du Grand Rift, depuis plus de 20 millions d'années, aurait fini par créer une différenciation climatique et environnementale entre l'Afrique de l'Est et le reste du continent. La savane à l'est du grand Rift aurait favorisé la naissance de la bipédie. À partir d'une souche commune, la lignée des hominines s'est séparée de celle des chimpanzés il y a plus de 7 millions d'années, les hominines se développant à l'Est et les pré-chimpanzés à l'Ouest.
Toutefois, la locomotion encore partiellement arboricole des Australopithèques, ainsi que les découvertes au Tchad d’Australopithecus bahrelghazali en 1995 et de Sahelanthropus tchadensis en 2001 ont fragilisé ce modèle[26]. De plus, les découvertes d’Ardipithecus ramidus en 1994 et d’Orrorin tugenensis en 2000 suggèrent que la bipédie a pu naitre dans des environnements boisés au sein de paysages mosaïques, et pas nécessairement dans des paysages de savanes[27].
La première révolution génétique
L'étude de l'évolution humaine connait une révolution quand Vincent Sarich (en) et Allan Wilson comparent les réactions immunologiques de l'albumine sérique des humains à celles de singes africains (chimpanzés et gorilles)[28]. En 1967 dans un article fondamental publié par la revue Science, grâce à la biologie moléculaire, ils montrent que la lignée humaine et celle des grands singes se sont séparées il y a quatre à cinq millions d'années[28] et que le matériel génétique de ces deux lignées ne diffère pas de plus de 2 %[29].
Les progrès du séquençage de l'ADN, notamment du génome mitochondrial (ADNmt) et de l'ADN du chromosome Y (en) (Y-ADN), ont éclairé la compréhension des origines de l'homme[30] - [31] - [32].
L'anthropologie moléculaire permet de définir les parentés entre espèces modernes. Ainsi, l'orang-outan est le dernier représentant d'un groupe de grands singes asiatiques, tandis que le gorille, le chimpanzé et l'Homme appartiennent au groupe des grands singes africains[33]. Les assertions de Linné, Darwin et Huxley s'en trouvent confirmées : les singes sont les animaux les plus proches de l'homme et les origines de la lignée humaine se trouvent en Afrique[33].
« L'hypothèse de l'horloge moléculaire », basée sur la fréquence des mutations passées, a révolutionné l'étude de l'évolution moléculaire. La vitesse de l'horloge moléculaire est cependant inconnue ; son calibrage repose sur les découvertes de fossiles. Un premier calendrier de l'évolution des primates basé sur les modèles de biologie moléculaire a été publié en 1967[34], d'autres l'ont été en 2012, surtout concernant Homo sapiens. Ainsi, le plus faible taux de mutation proposé met l'ancêtre commun de l'homme et de l'orang-outan à 40 millions d'années[35] - [36] - [37].
Une évolution linéaire
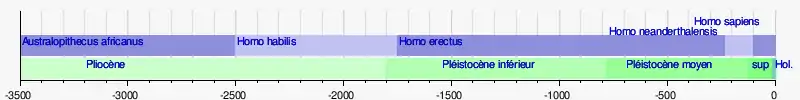
Jusqu'au début des années 1990, l'état de la recherche permettait de penser que l'arbre évolutif des genres Australopithèque et Homo était linéaire et que les espèces se succédaient dans un processus continu et régulier, chaque espèce étant l'ancêtre de l'autre. Cette hypothèse a connu son apogée dans le dernier quart du XXe siècle, époque de forte influence de la Théorie synthétique de l'évolution dans différentes disciplines de la paléoanthropologie. La théorie de l'espèce unique envisageait également qu'à une époque donnée ne pouvait exister qu'une seule espèce d'Hominine, puis d'Homo. L'arbre évolutif de l'Homme était alors perçu comme « un gros tronc avec très peu de branches ».
Dans ce cadre, les recherches sur les origines de l'Homme moderne ont conduit certains chercheurs dans les années 1980 et 1990 à considérer l'Homme de Néandertal comme une variété d'Homo sapiens, tant sur le plan de son anatomie que de son comportement (Homo sapiens neanderthalensis)[38].
La quête du premier hominine
Dans les années 1990, plusieurs équipes de paléoanthropologues ont travaillé en Afrique à la recherche du point de divergence entre la lignée des hominines et celle des chimpanzés.
En 1992, Timothy White, Gen Suwa et Berhane Asfaw découvrent à Aramis, en Éthiopie, l’Ardipithecus ramidus (publié en 1994)[39] - [40], daté de 4,4 millions d'années[41]. En 1994, Meave Leakey et Alan Walker découvrent l’Australopithecus anamensis (publié en 1995), daté de 3,9 à 4,2 millions d'années[42].
En 1997, Yohannes Haile-Selassie découvre dans la vallée de l'Awash, en Éthiopie, les fossiles de l'Ardipithecus kadabba, datés de 5,5 à 5,8 millions d'années. Très fragmentaires, ils ne sont publiés qu'en 2004 par Yohannes Haile-Selassie et Timothy White[43].
En 2000, Martin Pickford et Brigitte Senut découvrent dans les collines de Tugen, au Kenya, un fossile âgé de près de 6 millions d'années, Orrorin tugenensis, dont ils montrent la bipédie et qu'ils attribuent aux hominines (publié en 2001)[44].
En 2001, la Mission paléoanthropologique franco-tchadienne, dirigée par Michel Brunet, découvre au Tchad un fossile daté de 7 millions d'années (publié en 2002). Baptisé Sahelanthropus tchadensis, et surnommé Toumaï, le crâne présente les caractères faciaux et dentaires généralement associés aux hominines, et un trou occipital relativement avancé comme on en trouve chez des espèces bipèdes. Quelques scientifiques, comme Brigitte Senut et Martin Pickford, soulignent que la bipédie de Toumaï ne parait pas certaine[45] - [46].
Les fluctuations climatiques faisaient périodiquement varier l'environnement est-africain et laissaient des mosaïques de paysages (rivières, lacs, marécages, zones boisées, ilots forestiers, savane arborée, prairies herbeuses et zones désertiques) dans lesquels sont apparus nombre d'hominines bipèdes[47]. Cette diversité de formes « rappelle une règle trop souvent oubliée dans l'évolution des lignées : les grandes adaptations ne sont pas apparues en une seule fois et, chez des lignées proches, se font en mosaïque », avec des cas d'homoplasies (convergences, parallélismes)[48].
La seconde révolution génétique
En 2010, on commence à publier des éléments du patrimoine génétique néandertalien[49]. Pour mieux comprendre la période préhistorique récente, les généticiens recherchent aussi les restes de groupes de gènes (haplotypes) néandertaliens persistant dans le génome humain moderne[50] - [51]. Ils en ont notamment détecté un peu plus chez les Eurasiens de l'Est[52] - [53]. Ils recherchent aussi des groupes de gènes provenant d'hybridations entre Néandertaliens et Sapiens pré-modernes[54] - [55] ou Dénisoviens[56] - [57]).
En 2016 ont été publiés les premiers résultats d'une étude internationale fondée sur une nouvelle méthode de recherche de l'ADN hérité de plusieurs ancêtres humains archaïques. Cette méthode a été appliquée à des séquences du génome entier de 1523 personnes d'origine géographique diversifiée, incluant 35 nouveaux génomes identifiés dans les Îles mélanésiennes. Les auteurs de cette étude estiment avoir retrouvé 1,34 Gb de gènes d'origine néandertalienne et 303 Mb de gènes provenant de l'Homme de Denisova. Ils concluent de la cartographie de ces séquences « archaïques » que des populations néandertaliennes se sont sexuellement mélangées à celles de nos ancêtres directs, à plusieurs reprises et dans plusieurs populations non-africaines. On avait déjà trouvé des similarités entre génome contemporain et néandertalien[58], mais c'est la première fois qu'on trouve des gènes provenant a priori de l'Homme de Dénisova[59].
Un travail de caractérisation des régions génomiques contenant le plus ou le moins de séquences archaïques est en cours, qui permettra éventuellement de détecter les signatures d'introgressions adaptatives[60].
Un arbre buissonnant

Jusqu'à la fin du XXe siècle, l'arbre phylogénétique de la lignée humaine était souvent présenté de manière plus ou moins linéaire, avec un groupe préhumain précédant l'émergence du genre Homo. L’enrichissement du registre fossile et l'amélioration des techniques de datation ont remis en cause ce schéma[61]. Depuis les années 1980, les découvertes de fossiles se sont multipliées en Afrique et dans tout l'Ancien Monde, et, avec elles, le nombre d'espèces de la sous-tribu des hominines et du genre Homo. Du même coup, l'histoire évolutive de la lignée humaine est passée d'un schéma linéaire à un arbre buissonnant, et des espèces que nous pensions être nos ancêtres il y a encore peu sont devenues nos défunts cousins.
Les recherches récentes, par exemple celles menées par Lee Rogers Berger à l'université du Witwatersrand dans le Transvaal, sont bien plus compatibles avec une modélisation de structures buissonnantes et interpénétrées ou encore de rivières méandreuses dans une large vallée.
Chaque mise au jour de fossiles anciens, qu'ils appartiennent au genre homo ou australopithèque, accroit fortement le nombre d'informations anatomiques, techniques ou culturelles susceptibles d'être incompatibles avec le modèle linéaire des années 1970 et 1980.
Notes et références
Notes
- Il s'agit du squelette d'un homme datant de 33 000 ans.
Références
- Pascal Picq, [2005], p. 17-19
- Pascal Picq, [2005], p. 22-24
- American Heritage Dictionaries (editors) 2006
- Pascal Picq, [2005], p. 30-31
- Pascal Picq, Laurent Lemire, À la recherche de l'homme, Robert Laffont, , p. 33.
- Browne, p. 33-42
- Pascal Picq, [2005], p. 32-37
- Darwin 1871
- Pascal Picq, [2003]
- Pascal Picq, [2005], p. 48
- Rupke 2009, p. 195–196
- Pascal Picq, [2005], p. 50-52
- Montgomery 1988, p. 95–96
- L'homme de la Chapelle-aux-Saints, 1998
- Bernard Wood et alii, Wiley-Blackwell Encyclopedia of Human Evolution, June 2013 (single-volume paperback version of the original 2011 2-volume edition), 1056 pp.; (ISBN 978-1-1186-5099-8) (en)
- Pascal Picq, [2005], p. 53-55
- Raymond Dart, « Australopithecus africanus: The Man-Ape of South Africa », Nature, London, Nature Publishing Group, vol. 115, no 2884, , p. 195–199 (ISSN 0028-0836, DOI 10.1038/115195a0, lire en ligne [PDF], consulté le )
- Pascal Picq, [2005], p. 58-59
- Roger Lewin, Bones of Contention: Controversies in the Search for Human Origins, Chicago: The University of Chicago Press, (ISBN 0-226-47651-0), 1987
- Pascal Picq, [2005], p. 76-791
- Pascal Picq, [2005], p. 56-57
- Pascal Picq, [2005], p. 60-61
- Maurice Taieb, « L’Afrique, terre d’origine de l’humanité », Echosciences, janvier 2007 [lire en ligne]
- Academia Le Chemin de l' Humanité par René Puech
- Pascal Picq, [2005], p. 67-70
- Brunet, M. (1997), « Origine des hominidés : East Side Story... West Side Story... », Géobios, M.S. n ° 20, 79-83.
- Michel Brunet, Jean-Jacques Jaeger, « De l’origine des anthropoïdes à l’émergence de la famille humaine », Comptes Rendus Palevol, vol. 16, no 2, mars–avril 2017, p. 189-195 (DOI 10.1016/j.crpv.2016.04.007).
- V. M. Sarich et A. C. Wilson, « Immunological time scale for hominid evolution », Science, vol. 158, no 3805, , p. 1200–1203 (PMID 4964406, DOI 10.1126/science.158.3805.1200)
- (en) Sarich, V. M.; Wilson, A. C., « Immunological time scale for hominid evolution », Science, vol. 158, no 3805, , p. 1200–1203
- M'charek 2005, p. 96
- DeSalle et Tattersall 2008, p. 146
- Trent 2005, p. 6
- Pascal Picq, [2005], p. 65
- V. Sarich et A. Wilson, «Immunological Time Scale for Hominid Evolution», Science, vol. 158, 1967.
- Aylwyn Scally, Richard Durbin: Revising the human mutation rate: implications for understanding human evolution. In: Nature Reviews Genetics. Band 13, 2012, S. 745-753, doi:10.1038/nrg3295
- Studies slow the human DNA clock. In: Nature. Band 489, 2012, S. 343–344, doi:10.1038/489343a
- Kevin E. Langergraber et al.: Generation times in wild chimpanzees and gorillas suggest earlier divergence times in great ape and human evolution. In: PNAS. Band 109, Nr. 39, 2012, S. 15716–15721, doi:10.1073/pnas.1211740109
- Patrick Plumet, Peuples du Grand Nord. Des mythes à la Préhistoire, Errance, , p. 161.
- (en) Timothy White, Gen Suwa et Berhane Asfaw, « Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia », Nature (1994), 371, p. 306-312
- Timothy White, Gen Suwa et Berhane Asfaw, « Ardipithecus ramidus, a new species of early hominid from Aramis, Ethiopia », Nature (1995), 375, p. 88.
- Bernard A. Wood « The oldest hominid yet », Nature, (1994), 371, p. 280-1.
- (en) Warda C.V., « Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya », Journal of Human Evolution, vol. 41, , p. 255–368 (DOI 10.1006/jhev.2001.0507, lire en ligne)
- Jamie Shreeve, « The Evolutionary Road », National Geographic, Washington, D.C., National Geographic Society, (ISSN 0027-9358, lire en ligne, consulté le )
- B. Senut, M. Pickford, D. Gommery, P. Mein, K. Cheboi et Y. Coppens, « First hominid from the Miocene (Lukeino Formation, Kenya) », Comptes Rendus de l'Académie de Sciences, vol. 332, p. 137-144, 2001.
- Wolpoff Milford H., Senut B., Pickford M. et Hawks J., 2002, Sahelanthropus or "Sahelpithecus" ? Nature, 419 (6907): 581-582.
- Cela-Conde C. J. et Ayala F. J., 2003. Genera of the human lineage. PNAS, 100 (13): 7684-7689.
- Michel Brunet, Jean-Jacques Jaeger, « De l’origine des anthropoïdes à l’émergence de la famille humaine », Comptes Rendus Palevol, vol. 16, no 2, , p. 189-195 (DOI 10.1016/j.crpv.2016.04.007).
- Pascal Picq, Premiers hommes, Flammarion, , p. 87.
- Richard Green, Johannes Krause, David Reich, Svante Pääbo et al. (2010) A draft sequence of the Neandertal genome, Science 328, p. 710–722, doi:10.1126/science.1188021
- N. Patterson, P. Moorjani, Y. Luo, S. Mallick, N. Rohland, Y. Zhan, T. Genschoreck, T. Webster, D. Reich (2012) Ancient admixture in human history, Genetics 192, p. 1065–1093, doi:10.1534/genetics.112.145037
- Kay Prüfer, Svante Pääbo et al. (2014) The complete genome sequence of a Neanderthal from the Altai Mountains, Nature 505, p. 43–49, doi:10.1038/nature12886
- J. D. Wall, M. A. Yang, F. Jay, S. K. Kim, E. Y. Durand, L. S. Stevison, C. Gignoux, A. Woerner, M. F. Hammer, M. Slatkin (2011-2013) Higher levels of Neanderthal ancestry in East Asians than in Europeans, Genetics 194, p. 199–209, doi:10.1534/genetics.112.148213
- P. Skoglund, M. Jakobsson (2011) Archaic human ancestry in East Asia, Proceedings of the National Academy of Sciences U.S.A. 108, p. 18301–18306, doi:10.1073/pnas.1108181108
- S. Sankararaman, N. Patterson, H. Li, S. Pääbo, D. Reich (2012) The date of interbreeding between Neandertals and modern humans, PLOS Genetics 8, e1002947, doi:10.1371/journal.pgen.1002947
- B. Vernot & J. M. Akey (2015) Complex history of admixture between modern humans and Neandertals, American Journal of Human Genetics 96, p. 448–453, doi:10.1016/j.ajhg.2015.01.006
- David Reich, Richard Green, Svante Pääbo et al. (2010) Genetic history of an archaic hominin group from Denisova Cave in Siberia, Nature 468, p. 1053–1060, doi:10.1038/nature09710
- Matthias Meyer, M. Kircher, Svante Pääbo et al. (2012) A high-coverage genome sequence from an archaic Denisovan individual, Science 338, p. 222–226,
- S. Vattathil, J. M. Akey (2015) Small amounts of archaic admixture provide big insights into human history, Cell 163, p. 281–284, doi:10.1016/j.cell.2015.09.042
- Voir schéma (Fig. 1C and fig. S3) de l'article déjà cité
- Benjamin Vernot & al. (2016)) Excavating Neandertal and Denisovan DNA from the genomes of Melanesian individuals (résumé)
- Bernard Wood, « Le buissonnant rameau humain », Pour la Science, no 445, , p. 35.
Bibliographie
- Charles Darwin (trad. de l'anglais), La Filiation de l'homme et la sélection liée au sexe, Londres, John Murray (dir.), , 820 p. (ISBN 978-2-7453-2685-0, OCLC 7197127, lire en ligne)Traduction nouvelle réalisée sous la direction de Patrick Tort, Institut Charles Darwin International, Champion Classiques
- (en) Janet Browne, Charles Darwin : The Power of Place, Princeton University Press, , 591 p. (ISBN 978-0-691-11439-2, lire en ligne)
- (en) Nicolaas Adrianus Rupke, Richard Owen : Biology without Darwin, Chicago, University Of Chicago Press, , 344 p. (ISBN 978-0-226-73177-3), p. 182–253