Haplotype
Un haplotype est un groupe d'allèles de différents loci situés sur un même chromosome et habituellement transmis ensemble. Haplotype est un mot-valise formé par la contraction de la locution anglaise haploid genotype, ou génotype haploïde.
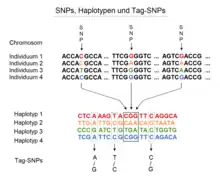
Définition
L'ensemble des gènes situés sur un même chromosome et dont les allèles ségrègent ensemble lors de la méiose constituent un haplotype. Ces gènes sont dits « génétiquement liés ». Deux gènes situés sur un même chromosome mais dont la transmission simultanée des allèles à la descendance est aléatoire sont dits syntènes. L'étude familiale de ces transmissions avait permis (avant la biologie moléculaire) d'établir la distance qui séparait deux gènes sur un même chromosome. Cette distance était exprimée en pourcentage de recombinaison, dont l'unité était le centimorgan, en hommage au généticien Morgan. Deux allèles qui ne se séparaient que dans 1 % des cas lors de leur transmission aux enfants étaient distants de 1 centimorgan. Si leur transmission simultanée n'avait lieu qu'une fois sur deux, il s'agissait d'une transmission « au hasard », ces gènes auxquels appartenaient ces allèles étaient dits indépendants.
Le terme est utilisé en immunologie pour désigner un ensemble de gènes liés situés sur un même chromosome et codant divers antigènes sanguins ou tissulaires. Les premiers systèmes de groupes sanguins définis par des haplotypes ont été les systèmes Rhésus en 1947, et MNS dont le lien entre les antigènes S ou s (MNS3 ou MNS4) et le système de groupe sanguin MN (antigènes M et N, MNS1 et MNS2) a été suspecté en 1947 et la description en haplotypes confirmée en 1951. Actuellement, le système le plus connu défini par des haplotypes est le complexe majeur d'histocompatibilité (système HLA) dont le rôle est primordial dans la mise en œuvre des réactions de défense immunologique de l'organisme. De nombreuses maladies génétiques trouvent leur origine dans une « anomalie » des gènes du système HLA.
Supposons deux gènes liés sur un même chromosome, c'est-à-dire très proches l'un de l'autre et ne se séparant pas par enjambement lors de la méiose. Par exemple dans le système de groupe sanguin MNS, sans considérer les nombreux variants mineurs de ce système, le gène codant une protéine, la glycophorine A (GPA), possède deux allèles M et N, et le gène codant une seconde protéine, la glycophorine B (GPB), possède deux allèles S et s. Il y a donc 4 haplotypes possibles dans ce système : MS, Ms, NS, Ns. Supposons un père ayant reçu de ses parents les haplotypes MS et Ns donc de génotype MS/Ns et de phénotype MSNs, et une mère homozygote MS/MS de phénotype MMSS. Leurs enfants seront nécessairement MS de par leur mère, et seront MS ou Ns de par leur père, mais ne pourront pas être Ms ou NS de par leur père. À moins d'une exceptionnelle recombinaison, dont la fréquence de survenue dans les familles donne une idée de la distance entre les gènes exprimée en centimorgans, les enfants MM seront nécessairement SS, et les enfants MN seront nécessairement Ss.
| Haplotypes maternels | |||
|---|---|---|---|
| MS | MS | ||
| Haplotypes paternels | MS | MS /MS | MS /MS |
| Ns | MS / Ns | MS / Ns | |
Ainsi, dans l'exemple précédent, sur quatre enfants, deux sont MMSS, et deux MNSs. Si nous connaissons à l'origine les haplotypes paternels, nous concluons qu'il n'y a aucun recombinant. Si nous observions, dans cette famille un cinquième enfant qui serait MNSS (l'ensemble des autres marqueurs confirmant la paternité), ce serait un recombinant, et nous dirions que le pourcentage de recombinaison dans cette dernière famille est d'un enfant sur cinq, soit 20 %. Si nous ne connaissons pas les haplotypes parentaux, la probabilité de recombinaison calculée n'est pas exactement la même du fait que les haplotypes parentaux sont déterminés à partir des enfants. Il fallait donc un grand nombre de familles (nombreuses de préférence) d'au moins deux enfants et/ou des familles étudiées sur trois générations, pour estimer, avant la biologie moléculaire, une distance exprimée en centimorgans.