Acide lipoïque
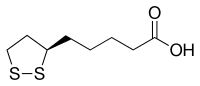
L'acide lipoïque, ou acide alpha-lipoïque[3] (ALA), appelé aussi acide thioctique[4], à ne pas confondre avec l'acide α-linolénique, est un composé organosulfuré dérivant de l'acide octanoïque, un acide carboxylique, et possédant un cycle 1,2-dithiolane. L'atome de carbone en C6 est stéréogène. Il existe par conséquent deux énantiomères de cette molécule : l'acide (R)-(+)-lipoïque et l'acide (S)-(-)-lipoïque, dont le mélange forme le racémique acide (R/S)-lipoïque. L'énantiomère (R) est celui qui est naturellement synthétisé par les animaux. Sa base conjuguée, l'anion lipoate, est un cofacteur intervenant sous forme d'un groupe prosthétique lié par covalence à certaines enzymes clés du métabolisme aérobie[5]. Sa forme oxydée se lie à un groupe acyle par une liaison acylthioester. Elle constitue un système rédox avec l'acide dihydrolipoïque, la forme réduite de l'acide lipoïque. Ce dernier est également distribué dans le commerce comme complément alimentaire antioxydant, ou comme médicament vendu en pharmacie, selon les pays.
| Acide lipoïque | ||
| ||
 | ||
| Structure de l'acide lipoïque. | ||
| Identification | ||
|---|---|---|
| Nom UICPA | acide (R)-5-(1,2-dithiolan-3-yl)pentanoïque | |
| Synonymes |
acide α-lipoïque, acide alpha-lipoïque, acide thioctique, acide 6,8-dithiooctanoïque |
|
| No CAS | D ou R(+) (RS) (RS) |
|
| No ECHA | 100.012.793 | |
| No CE | 214-071-2 (RS) 200-534-6 (RS) |
|
| Code ATC | A16 | |
| DrugBank | DB00166 | |
| PubChem | 6112 | |
| ChEBI | 30314 | |
| SMILES | ||
| InChI | ||
| Propriétés chimiques | ||
| Formule | C8H14O2S2 [Isomères] |
|
| Masse molaire[1] | 206,326 ± 0,018 g/mol C 46,57 %, H 6,84 %, O 15,51 %, S 31,08 %, |
|
| Propriétés physiques | ||
| T° fusion | 48 à 52 °C[2] | |
| Unités du SI et CNTP, sauf indication contraire. | ||
Biochimie
L'acide (R)-(+)-lipoïque est le seul énantiomère naturel de cette coenzyme. L'anion lipoate, sa base conjuguée, est la forme principale de cette molécule aux conditions physiologiques. L'acide lipoïque produit dans les tissus n'est pas libre dans les cellules car son précurseur, l'acide octanoïque, est lié aux complexes enzymatiques avant l'insertion enzymatique des atomes de soufre. Plus précisément, l'acide lipoïque est lié par une liaison amide à l'amine terminale de la chaîne latérale d'un résidu de lysine au niveau du domaine lipoyle de l'enzyme dont il est un cofacteur. L'une des fonctions les plus étudiées de l'acide lipoïque est son rôle de cofacteur du complexe pyruvate déshydrogénase, qui relie la glycolyse au cycle de Krebs, bien que ce ne soit pas le seul complexe enzymatique dont il est un cofacteur.
Biosynthèse
Précurseur de l'acide lipoïque, l'acide octanoïque est produit au cours de la biosynthèse des acides gras sous forme d'octanoyl-ACP. Chez les eucaryotes, ce dernier résulte d'une seconde voie de biosynthèse des acides gras, qui se déroule dans les mitochondries[6] - [7]. La biosynthèse des acides gras fournit l'unité octanoate. Celle-ci est transférée par une lipoyl(octanoyl) transférase depuis l'ACP vers l'amine terminale de la chaîne latérale d'un résidu de lysine du domaine lipoyle sous forme de thioester. Une lipoyle synthase catalyse le remplacement de deux atomes d'hydrogène de l'octanoate par des atomes de soufre à partir de SAM à l'aide d'un mécanisme radicalaire[8]. Pour cette raison, l'acide lipoïque est synthétisé en étant déjà lié aux protéines, et non sous forme libre dans les cellules. Il peut être éliminé lorsque les protéines sont dégradées, ainsi que par l'action d'une lipoamidase[9]. Certains êtres vivants peuvent fixer l'acide lipoïque libre sur des enzymes à l'aide d'une lipoate-protéine ligase (EC ) ; l'activité de cette enzyme requiert l'hydrolyse concomitante d'ATP[10].
Activité coenzymatique
L'acide lipoïque est cofacteur d'au moins cinq enzymes. Deux d'entre elles appartiennent au cycle de Krebs, voie métabolique centrale de la respiration cellulaire chez de nombreux êtres vivants convertissant leurs nutriments en énergie métabolique. Ces enzymes lipoylées possèdent un résidu d'acide lipoïque liées à elles par covalence. Le groupe lipoyle transfère d'une part les groupes acyle dans les complexes enzymatiques à α-cétoacide déshydrogénase (ou 2-oxoacide déshydrogénase), et d'autre part les groupes méthylamine dans le système de clivage de la glycine et dans la glycine déshydrogénase.
Les réactions de transfert des complexes à 2-oxoacide déshydrogénase font appel à des mécanismes semblables dans les quatre enzymes suivantes :
- la dihydrolipoamide S-acétyltransférase, enzyme E2 du complexe pyruvate déshydrogénase ;
- la dihydrolipoamide S-succinyltransférase, enzyme E2 du complexe α-cétoglutarate déshydrogénase ;
- la dihydrolipoyl transacylase, enzyme E2 du complexe 3-méthyl-2-oxobutanoate déshydrogénase ;
- la dihydrolipoamide acétyltransférase, enzyme E2 du complexe acétoïne déshydrogénase.
Ces complexes sont chacun composés de trois sous-unités, appelées E1, E2 et E3, qui sont respectivement une décarboxylase, une acétyltransférase de type lipoyl transférase, et une réductase de type dihydrolipoyl déshydrogénase. L'enzyme E2 forme le cœur de ces complexes, autour duquel les autres sous-unités s'organisent. Le domaine lipoyle véhicule les intermédiaires entre les sites actifs des différentes sous-unités enzymatiques de ces complexes[11] - [12]. Le domaine lipoyle lui-même est lié au cœur E2 par un bras moléculaire flexible. Le nombre de domaines lipoyle varie de un à trois selon les organismes considérés ; ce nombre peut être modifié artificiellement, ce qui a montré que l'activité des complexes décroît au-delà de trois domaines lipoyle[13], et que la croissance des organismes est altérée lorsqu'on en ajoute plus de 9.
L'acide lipoïque intervient également comme cofacteur du complexe acétoïne déshydrogénase, qui, chez certaines bactéries, catalyse la conversion de la 3-hydroxybutanone (acétoïne) CH3–CHOH–CO–CH3 en acétaldéhyde CH3CHO et acétyl-CoA, permettant à la 3-hydroxybutanone d'être utilisée comme source de carbone.
Le système de clivage de la glycine diffère des autres complexes, et sa nomenclature est différente. Les composantes de ce système sont libres, de sorte que l'ensemble ne forme pas un complexe. Dans ce système, la protéine H est un domaine lipoyle libre avec des hélices α supplémentaires, la protéine L est une dihydrolipoyl déshydrogénase, la protéine P est une décarboxylase, et la protéine T est une protéine qui transfère la méthylamine depuis le lipoate vers le tétrahydrofolate (THF) pour donner du méthylène-THF et de l'ammoniac NH3. Le méthylène-THF est ensuite utilisé par la sérine hydroxyméthyltransférase pour synthétiser de la sérine à partir de glycine. Ce système fait partie de la photorespiration des plantes[14].
Biosynthèse et dégradation
L'acide lipoïque est présent en petite quantité dans presque tous les types d'aliments, mais avec une abondance légèrement supérieure dans les reins, le cœur, le foie, les épinards, les brocoli et les extraits de levure[15]. La molécule naturelle est toujours liée par covalence à une protéine et n'est donc pas directement disponible à partir de l'alimentation. La quantité totale d'acide lipoïque présente dans les sources alimentaires est par ailleurs très faible : on estime à 10 tonnes la masse de foie nécessaire à la purification de 30 mg d'acide lipoïque en vue de la détermination de sa structure[16]. En conséquence, tout l'acide lipoïque disponible comme complément alimentaire est synthétisé par voie chimique.
Les niveaux d'acide lipoïque et d'acide dihydrolipoïque dans le plasma sanguin humain sont indétectables en l'absence de supplémentation[17]. Après hydrolyse acide, qui libère l'acide lipoïque lié aux protéines, le taux plasmatique d'acide lipoïque a été mesuré entre 12,3 et 43,1 ng/mL, tandis que l'hydrolyse enzymatique libère jusqu'à 38,2 ng/mL par la subtilisine[18] - [19] - [20]. Les lipoamidases sériques clivent rapidement le lipoamide synthétique et la (R)-lipoyl-L-lysine en libérant de l'acide (R)-lipoïque et de la L-lysine ou de l'ammoniac[21] - [22] - [23].
Pharmacologie
L'acide lipoïque est commercialisé comme complément alimentaire avant tout pour ses propriétés antioxydantes[24], ainsi que pour tout un ensemble de bienfaits potentiels[25]. Néanmoins, son mécanisme d'action dans cet usage demeure controversé. Dans les cellules, l'acide lipoïque semble induire avant tout une réponse au stress oxydant plutôt qu'éliminer directement les radicaux libres. Cet effet est spécifique à l'énantiomère (R)[3]. Bien que le milieu intracellulaire soit fortement réducteur, l'acide lipoïque y a été observé à la fois sous forme oxydée et sous forme réduite[26] (acide dihydrolipoïque). L'acide lipoïque est capable in vitro d'éliminer les espèces réactives de l'oxygène ainsi que de celles de l'azote, mais il n'y a pas d'éléments clairs étayant que cela se produise également in vivo dans les cellules, ni que l'élimination des radicaux libres soit à la base de l'action physiologique de cette molécule[3] - [25]. L'acide lipoïque est également assez efficace pour neutraliser l'acide hypochloreux HOCl, bactéricide produit par les granulocytes neutrophiles susceptible d'induire une inflammation et d'endommager les tissus ; cette activité provient de la conformation tendue du cycle dithiolane, qui est relâchée lors de la réduction en acide dihydrolipoïque. C'est essentiellement sous cette forme que l'acide lipoïque est présent dans les cellules, l'acide dihydrolipoïque étant généralement considérée comme la forme la plus bioactive de l'acide lipoïque, responsable de l'essentiel de son activité antioxydante[27]. Cette conception a cependant été discutée en raison du fait que les deux groupes sulfhydryle sont très réactifs et tendent à être méthylés rapidement, que la concentration intracellulaire de l'acide dihydrolipoïque libre est faible (d'autant qu'il tend à être rapidement éliminé des cellules), et que la chaîne carbonée est facilement rompue par oxydation pour former des composés plus courts. Il est possible que les effets thérapeutiques et anti-âge de l'acide lipoïque proviennent d'une modulation d'une transduction de signal et de la transcription de certains gènes : ceci aurait pour effet d'améliorer l'état rédox de la cellule en activant des mécanismes antioxydants, et expliquerait mieux l'action de l'acide lipoïque que l'élimination des radicaux libres ou la réduction des espèces oxydantes réactives[3] - [25].
Toutes les formes disulfure de l'acide lipoïque peuvent être réduites en acide dihydrolipoïque. Il existe des mécanismes de réduction spécifiques à certains tissus, et d'autres qui sont spécifiques à un énantiomère plutôt qu'à un autre. Au moins deux enzymes cytosoliques (la glutathion réductase et la thiorédoxine réductase Trx1) ainsi que deux enzyme mitochondriales (la dihydrolipoyl déshydrogénase et la thiorédoxine réductase Trx2) sont susceptibles de réduire l'acide (R)-lipoïque de manière stéréospécifique. L'acide (R)-(+)-lipoïque est réduit en acide (R)-(-)-dihydrolipoïque, tandis que l'acide (S)-(-)-lipoïque est réduit en acide (S)-(+)-dihydrolipoïque[28] - [29] - [30] - [31] - [32] - [33] - [34].
L'énantiomère (S) est toxique lorsqu'il est administré à des souris présentant une déficience en thiamine[35] - [36]. Plusieurs études ont montré que l'acide (S)-lipoïque présente une activité inférieure à celle d'énantiomère (R), ou interfère avec celui-ci à la façon d'un inhibiteur compétitif[37] - [38] - [39].
En Belgique
En Belgique, le Conseil Supérieur de la Santé émet une opinion d'expert quant à l'innocuité de la commercialisation de compléments alimentaires contenant de l'acide alpha-lipoïque (environ 260 produits enregistrés en Belgique) destinés à la population générale. À ce sujet, le Conseil recommande de considérer l'acide alpha-lipoïque comme un médicament et non comme un complément alimentaire (composant) et donc de retirer ces produits du marché des compléments alimentaires en vente libre au grand public[40].
Références
- Cet article est partiellement ou en totalité issu de l'article intitulé « Acide alpha-lipoïque » (voir la liste des auteurs).
- Masse molaire calculée d’après « Atomic weights of the elements 2007 », sur www.chem.qmul.ac.uk.
- Fiche Sigma-Aldrich du composé (R)-(+)-α-Lipoic acid analytical standard, consultée le 22 mars 2017.
- (en) Kate Petersen Shay, Régis F. Moreau, Eric J. Smith et Tory M. Hagen, « Is α-lipoic acid a scavenger of reactive oxygen species in vivo? Evidence for its initiation of stress signaling pathways that promote endogenous antioxidant capacity », IUBMB Life, vol. 60, no 6, , p. 362-367 (PMID 18409172, DOI 10.1002/iub.40, lire en ligne)
- https://www.vidal.fr/parapharmacie/complements-alimentaires/acide-alpha-lipoique-thioctique.html
- (en) G. Raddatz et H. Bisswanger, « Receptor site and stereospecifity of dihydrolipoamide dehydrogenase for R- and S-lipoamide: a molecular modeling study », Journal of Biotechnology, vol. 58, no 2, , p. 89-100 (PMID 9383983, DOI 10.1016/S0168-1656(97)00135-1, lire en ligne)
- (en) John E. Cronan, Ian L. Fearnley et John E. Walker, « Mammalian mitochondria contain a soluble acyl carrier protein », FEBS Letters, vol. 579, no 21, , p. 4892-4896 (PMID 16109413, DOI 10.1016/j.febslet.2005.07.077, lire en ligne)
- (en) Sean W. Jordan et John E. Cronan Jr., « A new metabolic link. The acyl carrier protein of lipid synthesis donates lipoic acid to the pyruvate dehydrogenase complex in Escherichia coli and mitochondria. », The Journal of Biological Chemistry, vol. 272, no 29, , p. 17903-17906 (PMID 9218413, DOI 10.1074/jbc.272.29.17903, lire en ligne)
- (en) Robert M. Cicchillo et Squire J. Booker, « Mechanistic Investigations of Lipoic Acid Biosynthesis in Escherichia coli: Both Sulfur Atoms in Lipoic Acid are Contributed by the Same Lipoyl Synthase Polypeptide », Journal of the American Chemical Society, vol. 127, no 9, , p. 2860-2861 (PMID 15740115, DOI 10.1021/ja042428u, lire en ligne)
- (en) Yanfang Jiang et John E. Cronan, « Expression cloning and demonstration of Enterococcus faecalis lipoamidase (pyruvate dehydrogenase inactivase) as a Ser-Ser-Lys triad amidohydrolase », The Journal of Biological Chemistry, vol. 280, no 3, , p. 2244-2256 (PMID 15528186, DOI 10.1074/jbc.M408612200, lire en ligne)
- (en) John E. Cronan, Xin Zhao et Yanfang Jiang, « Function, Attachment and Synthesis of Lipoic Acid in Escherichia coli », Advances in Microbial Physiology, vol. 50, , p. 103-146 (PMID 16221579, DOI 10.1016/S0065-2911(05)50003-1, lire en ligne)
- (en) Jacqueline L. S. Milne, Xiongwu Wu, Mario J. Borgnia, Jeffrey S. Lengyel, Bernard R. Brooks, Dan Shi, Richard N. Perham et Sriram Subramaniam, « Molecular Structure of a 9-MDa Icosahedral Pyruvate Dehydrogenase Subcomplex Containing the E2 and E3 Enzymes Using Cryoelectron Microscopy », The Journal of Biological Chemistry, vol. 281, no 7, , p. 4364-4370 (PMID 16308322, PMCID 1647297, DOI 10.1074/jbc.M504363200, lire en ligne)
- (en) Gavin E. Murphy et Grant J. Jensen, « Electron Cryotomography of the E. coli Pyruvate and 2-Oxoglutarate Dehydrogenase Complexes », Structure, vol. 13, no 12, , p. 1765-1773 (PMID 16338405, DOI 10.1016/j.str.2005.08.016, lire en ligne)
- (en) Rosane S. Machado, David P. Clark et John R. Guest, « Construction and properties of pyruvate dehydrogenase complexes with up to nine lipoyl domains per lipoate acetyltransferase chain », FEMS Microbiology Letters, vol. 100, nos 1-3, , p. 243-248 (PMID 1478460, DOI 10.1111/j.1574-6968.1992.tb14047.x, lire en ligne)
- (en) Roland Douce, Jacques Bourguignon, Michel Neuburger et Fabrice Rébeillé, « The glycine decarboxylase system: a fascinating complex », Trends in Plant Science, vol. 6, no 4, , p. 167-176 (PMID 11286922, DOI 10.1016/S1360-1385(01)01892-1, lire en ligne)
- (en) Arjumand I. Durrani, Heidi Schwartz, Michael Nagl et Gerhard Sontag, « Determination of free α-lipoic acid in foodstuffs by HPLC coupled with CEAD and ESI-MS », Food Chemistry, vol. 120, no 4, , p. 1143-1148 (DOI 10.1016/j.foodchem.2009.11.045, lire en ligne)
- (en) Lester J. Reed, « A trail of research from lipoic acid to alpha-keto acid dehydrogenase complexes », The Journal of Biological Chemistry, vol. 276, no 42, , p. 38329-38336 (PMID 11477096, DOI 10.1074/jbc.R100026200, lire en ligne)
- (en) R. Hermann, G. Niebch, H. O. Borbe, H. Fieger-Büschges, P. Ruus, H. Nowak, H. Riethmüller-Winzen, M. Peukert et H. Blume, « Enantioselective pharmacokinetics and bioavailability of different racemic α-lipoic acid formulations in healthy volunteers », European Journal of Pharmaceutical Sciences, vol. 4, no 3, , p. 167-174 (DOI 10.1016/0928-0987(95)00045-3, lire en ligne)
- (en) Jens Teichert et Rainer Preiss, « [17] High-performance liquid chromatography methods for determination of lipoic and dihydrolipoic acid in human plasma », Methods in Enzymology, vol. 279, , p. 159-166 (PMID 9211267, DOI 10.1016/S0076-6879(97)79019-0, lire en ligne)
- (en) Jens Teichert et Rainer Preiß, « Determination of lipoic acid in human plasma by high-performance liquid chromatography with electrochemical detection », Journal of Chromatography B: Biomedical Sciences and Applications, vol. 672, no 2, , p. 277-281 (PMID 8581134, DOI 10.1016/0378-4347(95)00225-8, lire en ligne)
- (en) J. Teichert et R. Preiss, « HPLC-methods for determination of lipoic acid and its reduced form in human plasma », International Journal of Clinical Pharmacology, Therapy, and Toxicology, vol. 30, no 11, , p. 511-512 (PMID 1490813)
- (en) Jun Oizumi et Kou Hayakawa, « Liberation of lipoate by human serum lipoamidase from bovine heart pyruvate dehydrogenase », Biochemical and Biophysical Research Communications, vol. 162, no 2, , p. 658-663 (PMID 2502979, DOI 10.1016/0006-291X(89)92361-9, lire en ligne)
- (en) Birgit Backman-Gullers, Ulf Hannestad, Lennart Nilsson et Bo Sörbo, « Studies on lipoamidase: Characterization of the enzyme in human serum and breast milk », Clinica Chimica Acta, vol. 191, nos 1-2, , p. 49-60 (PMID 2127386, DOI 10.1016/0009-8981(90)90057-Y, lire en ligne)
- (en) Cheryl L. Garganta et Barry Wolf, « Lipoamidase activity in human serum is due to biotinidase », Clinica Chimica Acta, vol. 189, no 3, , p. 313-325 (PMID 2225462, DOI 10.1016/0009-8981(90)90313-H, lire en ligne)
- (en) Anna Gorąca, Halina Huk-Kolega, Aleksandra Piechota, Paulina Kleniewska, Elżbieta Ciejka et Beata Skibska, « Lipoic acid – biological activity and therapeutic potential », Pharmacological Reports, vol. 63, no 4, , p. 849-858 (PMID 22001972, DOI 10.1016/S1734-1140(11)70600-4, lire en ligne)
- (en) Kate Petersen Shay, Régis F. Moreau, Eric J. Smith, Anthony R. Smith et Tory M. Hagen, « Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential », Biochimica et Biophysica Acta (BBA) - General Subjects, vol. 1790, no 10, , p. 1149-1160 (PMID 19664690, PMCID 2756298, DOI 10.1016/j.bbagen.2009.07.026, lire en ligne)
- (en) Lester Packer, Eric H. Witt et Hans Jürgen Tritschler, « Alpha-lipoic acid as a biological antioxidant », Free Radical Biology and Medicine, vol. 19, no 2, , p. 227-250 (PMID 7649494, DOI 10.1016/0891-5849(95)00017-R, lire en ligne)
- (en) Guido R. M. M. Haenen et Aalt Bast, « Scavenging of hypochlorous acid by lipoic acid », Biochemical Pharmacology, vol. 42, no 11, , p. 2244-2246 (PMID 1659823, DOI 10.1016/0006-2952(91)90363-A, lire en ligne)
- (en) Elias S. J. Arnér, Jonas Nordberg et Arne Holmgren, « Efficient Reduction of Lipoamide and Lipoic Acid by Mammalian Thioredoxin Reductase », Biochemical and Biophysical Research Communications, vol. 225, no 1, , p. 268-274 (PMID 8769129, DOI 10.1006/bbrc.1996.1165, lire en ligne)
- (en) John E. Biaglow, Iraimoudi S. Ayene, Cameron J. Koch, Jeremiah Donahue, Thomas D. Stamato, John J. Mieyal et Stephen W. Tuttle, « Radiation Response of Cells during Altered Protein Thiol Redox », Radiation Research, vol. 159, no 4, , p. 484-494 (PMID 12643793, DOI 10.1667/0033-7587(2003)159[0484:RROCDA]2.0.CO;2, lire en ligne)
- (en) Nobuya Haramaki, Derick Han, Garry J. Handelman, Hans J. Tritschler et Lester Packer, « Cytosolic and Mitochondrial Systems for NADH- and NADPH-dependent Reduction of α-Lipoic Acid », Free Radical Biology and Medicine, vol. 22, no 3, , p. 535-542 (PMID 8981046, DOI 10.1016/S0891-5849(96)00400-5, lire en ligne)
- (en) Anastasia Constantinescu, Uri Pick, Nobuya Haramaki, Derick Han, Maurizio Podda, Hans J. Tritschler et Lester Packer, « Reduction and transport of lipoic acid by human erythrocytes », Biochemical Pharmacology, vol. 50, no 2, , p. 253-261 (PMID 7632170, DOI 10.1016/0006-2952(95)00084-D, lire en ligne)
- (en) James M. May, Zhi-chao Qu et Deanna J. Nelson, « Cellular disulfide-reducing capacity: An integrated measure of cell redox capacity », Biochemical and Biophysical Research Communications, vol. 344, no 4, , p. 1352-1359 (PMID 16650819, DOI 10.1016/j.bbrc.2006.04.065, lire en ligne)
- (en) Wright Jones, Xia Li, Zhi-chao Qu, Laureta Perriott, Richard R. Whitesell et James M. May, « Uptake, recycling, and antioxidant actions of α-lipoic acid in endothelial cells », Free Radical Biology and Medicine, vol. 33, no 1, , p. 83-93 (PMID 12086686, DOI 10.1016/S0891-5849(02)00862-6, lire en ligne)
- (en) H. Schempp, H. Ulrich et E. F. Elstner, « Stereospecific reduction of R(+)-thioctic acid by porcine heart lipoamide dehydrogenase/diaphorase », Zeitschrift für Naturforschung C, vol. 49, nos 9-10, , p. 691-692 (PMID 7945680, DOI 10.1515/znc-1994-9-1023, lire en ligne)
- (en) Emery M. Gal et Dzidra E. Razevska, « Studies on the in vivo metabolism of lipoic acid. I. The fate of dl-lipoic acid -S35 in normal and thiamine-deficient rats », Archives of Biochemistry and Biophysics, vol. 89, no 2, , p. 253-261 (PMID 13825981, DOI 10.1016/0003-9861(60)90051-5, lire en ligne)
- (en) E. Martin Gal, « Reversal of Selective Toxicity of (−)-α-Lipoic Acid by Thiamine in Thiamine-deficient Rats », Nature, vol. 207, no 996, , p. 535 (PMID 5328673, DOI 10.1038/207535a0, lire en ligne)
- (en) F. Kilic, G. J. Handelman, E. Serbinova, L. Packer et J. R. Trevithick, « Modelling cortical cataractogenesis 17: in vitro effect of a-lipoic acid on glucose-induced lens membrane damage, a model of diabetic cataractogenesis », Biochemistry and Molecular Biology International Journal, vol. 37, no 2, , p. 361-370 (PMID 8673020)
- (en) R. S. Streeper, E. J. Henriksen, S. Jacob, J. Y. Hokama, D. L. Fogt et H. J. Tritschler, « Differential effects of lipoic acid stereoisomers on glucose metabolism in insulin-resistant skeletal muscle », The American Journal of Physiology, vol. 273, no 1 Pt 1, , E185-E191 (PMID 9252495, lire en ligne)
- (en) L. Frölich, M. E. Götz, M. Weinmüller, M. B. H. Youdim, N. Barth, A. Dirr, W. Gsell, K. Jellinger, H. Beckmann, P. Riederer, « (r)-, but not (s)-alpha lipoic acid stimulates deficient brain pyruvate dehydrogenase complex in vascular dementia, but not in Alzheimer dementia », Journal of Neural Transmission, vol. 111, no 3, , p. 295-310 (PMID 14991456, DOI 10.1007/s00702-003-0043-5, lire en ligne)
- « Innocuité de l’acide alpha-lipoïque dans les compléments alimentaires », AVIS DU CONSEIL SUPERIEUR DE LA SANTE N° 9274, Conseil Supérieur de la Santé, (consulté le )