Enzyme de conversion de l'angiotensine 2
L'enzyme de conversion de l'angiotensine 2, ou ACE2 (de l'anglais Angiotensin-Converting Enzyme 2), est une enzyme liée à la face externe des membranes plasmiques de cellules du poumon, des artères, du cœur, du rein et de l'appareil digestif[3] - [4] - [5]. Elle joue un rôle important dans le système rénine-angiotensine-aldostérone (SRAA), qui joue un rôle physiologique majeur pour la régulation du coeur, du rein et de la pression sanguine[6] - [7] - [8] - [9], qui régule l'homéostasie hydrosodée et la pression artérielle.
| Enzyme de conversion de l'angiotensine 2 | ||
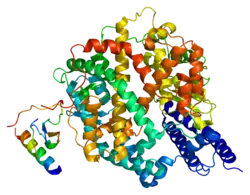 Enzyme de conversion de l'angiotensine 2 humaine (PDB 1R42[1]) | ||
| Caractéristiques générales | ||
|---|---|---|
| Nom approuvé | Angiotensin I Converting Enzyme 2 | |
| Symbole | ACE2 | |
| Synonymes | ACEH | |
| N° EC | 3.4.17.23 | |
| Homo sapiens | ||
| Locus | Xp22.2 | |
| Masse moléculaire | 92 463 Da[2] | |
| Nombre de résidus | 805 acides aminés[2] | |
| Entrez | 59272 | |
| HUGO | 13557 | |
| OMIM | 300335 | |
| UniProt | Q9BYF1 | |
| RefSeq (ARNm) | NM_021804.2 | |
| RefSeq (protéine) | NP_068576.1 | |
| Ensembl | ENSG00000130234 | |
| PDB | 1R42, 1R4L, 1XJP, 2AJF, 3D0G, 3D0H, 3D0I, 3KBH, 3SCI, 3SCJ, 3SCK, 3SCL, 6ACG, 6ACJ, 6ACK, 6CS2 | |
|
GENATLAS • GeneTests • GoPubmed • HCOP • H-InvDB • Treefam • Vega | ||
| Liens accessibles depuis GeneCards et HUGO. | ||
Il s'agit d'une métallocarboxypeptidase (en) transmembranaire qui catalyse le clivage de l'angiotensine I 1–10 en angiotensine 1–9[4] et de l'angiotensine II 1–8, peptide induisant une vasoconstriction, en angiotensine 1–7, peptide induisant une vasodilatation[10] - [11]. Elle s'oppose à l'action de l'enzyme de conversion de l'angiotensine (ACE), qui catalyse la conversion de l'angiotensine I en angiotensine II, de sorte que l'ACE2 réduit la concentration en angiotensine II[12] ; celle-ci agissant comme vasoconstricteur, cela fait de l'ACE2 une cible intéressante pour le traitement des maladies cardiovasculaires[12] - [13].
L'ACE2 est le récepteur de certains coronavirus à la membrane plasmique des cellules. C'est le cas pour le coronavirus humain NL63, le SARS-CoV-1, coronavirus à l'origine du SRAS, et le SARS-CoV-2, à l'origine de la COVID-19[14] - [15] - [16] - [17]. Plusieurs types de cellules humaines sont dotées d'ACE2, de même pour d'autres mammifères, dont – à titre d'exemple – le chat, le chien, le pangolin, le vison, le Hamster de Chine et d'autres rongeurs du groupe des Cricetidae[18] - [19] - [20] - [21].
Le gène
L'enzyme de conversion de l'angiotensine 2 est codée par le gène ACE2 (angiotensin-converting enzyme 2), identifié en 2000[4]. Il est situé sur le chromosome X humain.
La protéine
Il s'agit d'une métalloprotéinase (métalloexopeptidase). Elle est présente principalement dans les cellules endothéliales vasculaires du cœur et intervient dans la fonction cardiaque. Elle est également exprimée dans le rein et dans les testicules[4]. Elle est donc moins ubiquitaire que son homologue, l'enzyme de conversion de l'angiotensine, renommée enzyme de conversion de l'angiotensine 1 (ACE1) depuis la découverte de l'ACE2.
La structure secondaire de l'ACE2 contient 20 hélices α et 9 hélices 310 qui totalisent environ 62 % de la structure. Elle contient également 6 feuillets β qui totalisent 3,5 % supplémentaires. La région extracellulaire de l'ACE2 humaine contient deux domaines : le domaine métallopeptidase à zinc et le domaine d'homologie aux collectines C-terminal, qui est désordonné ; le domaine métallopeptidase à zinc peut lui-même être divisé en deux sous-domaines, notés I et II, qui entourent le site actif, dont le sous-domaine I, du côté N-terminal, contient le cation de zinc Zn2+.
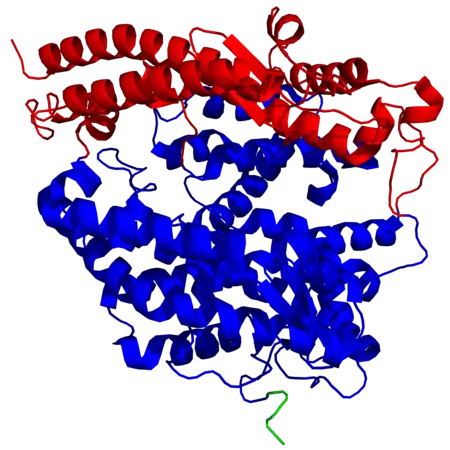 Structure d'une ACE2 humaine montrant les sous-domaines I en rouge et II en bleu du domaine métallopeptidase à zinc ainsi que le début du domaine d'homologie aux collectines C-terminal en vert (PDB 1R42[1]).
Structure d'une ACE2 humaine montrant les sous-domaines I en rouge et II en bleu du domaine métallopeptidase à zinc ainsi que le début du domaine d'homologie aux collectines C-terminal en vert (PDB 1R42[1]).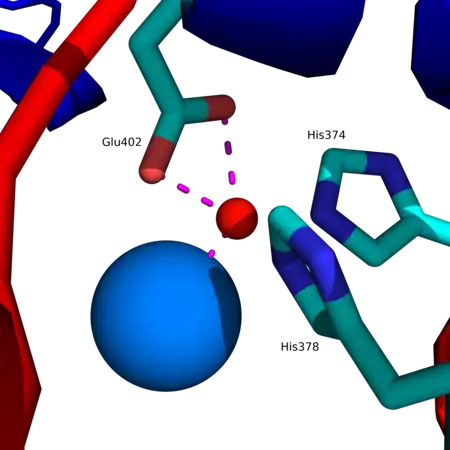 Site actif d'une ACE2 humaine avec le motif HEXXH + E montrant le cation de zinc Zn2+ en rouge coordonné aux résidus His374, His378 et Glu402, ainsi qu'à une molécule d'eau (en bleu).
Site actif d'une ACE2 humaine avec le motif HEXXH + E montrant le cation de zinc Zn2+ en rouge coordonné aux résidus His374, His378 et Glu402, ainsi qu'à une molécule d'eau (en bleu).
Le cation de zinc Zn2+, qui intervient comme cofacteur dans le site actif de l'enzyme, est coordonné aux résidus His374, His378, Glu402 et à une molécule d'eau H2O (à l'état natif). Ces résidus forment la séquence HEXXH + E, où H = histidine, E = glutamate et X = acide aminé indéterminé ; ce motif est conservé dans les métallopeptidases à zinc. L'anion chlorure Cl– est coordonné aux résidus Arg169, Trp477 et Lys481 du côté C-terminal[1].
Rôle
| N° EC | EC |
|---|---|
| Cofacteur(s) | Zn2+[22] |
| IUBMB | Entrée IUBMB |
|---|---|
| IntEnz | Vue IntEnz |
| BRENDA | Entrée BRENDA |
| KEGG | Entrée KEGG |
| MetaCyc | Voie métabolique |
| PRIAM | Profil |
| PDB | RCSB PDB PDBe PDBj PDBsum |
L'ACE1 convertit l'angiotensine I (décapeptide) en angiotensine II (octapeptide) par clivage des deux résidus d'acides aminés C-terminaux :
L'ACE2 convertit l'angiotensine I 1-10 (décapeptide) en angiotensine 1-9 (nonapeptide) et l'angiotensine II 1-8 (octapeptide induisant une vasoconstriction) en angiotensine 1-7 (heptapeptide ayant des propriétés vasodilatatrices)[5] - [23] - [24] :
L'effet de l'ACE2 est antagoniste à celui de l'ACE1, en déplaçant l'équilibre de production depuis l'angiotensine I vers angiotensine 1-7, ce qui correspond fonctionnellement à un mécanisme de clairance de l'angiotensine II[25]. L'angiotensine 1-7 est un vasodilatateur qui antagonise les effets de l'angiotensine II en interagissant avec un récepteur couplé à une protéine G, l'oncogène Mas[26].
L’homéostasie du muscle cardiaque serait en partie sous la dépendance de l’expression de l’angiotensine II et de l’angiotensine 1-7. Alors que l’inactivation de l’ACE chez la souris induit une hypertension, celle de l’ACE2 entraîne des anomalies cardiaques : dilatation ventriculaire gauche, amincissement des parois et surtout hypocontractilité ventriculaire gauche[27]. Cependant, ceci n'a pas été retrouvé dans d'autres études qui montrent une fonction cardiaque normale de base[28], mais une augmentation de l'hypertrophie des parois de cardiaque en cas de surcharge de pression[29].
Le rôle de l'ACE2 dans le rein semble également essentiel[30]. Elle serait principalement localisée au niveau du tubule rénal. Les effets d'un antagoniste de l'ACE2, le MLN-4760, commencent à être évalués chez l'animal[31].
Régulation
Contrairement à l'ACE1, l'ACE2 est insensible aux inhibiteurs de l'enzyme de conversion[32]. Son expression s'élève en cas d'insuffisance cardiaque[33].
On a constaté que l'exercice physique, s'il est chronique, peut être l'un des stimuli importants de l'ACE2 dans le cerveau et sa périphérie[34] - [35]. On a constaté en 2020 que « cependant, si l'exercice est clairement associé à de meilleurs résultats cardiovasculaires dans des situations chroniques, l'exercice peut contribuer à un plus grand risque d'infection par le SARS-CoV-2 ». Ainsi Keidar et al. avaient déjà rapporté en 2005 que l'antagoniste des minéralocorticoïdes, la spironolactone, avait quadruplé l'activité ACE2 dans les macrophages dérivés de monocytes de patients atteints de CHF[36] ; mais la spironolactone n'augmente pas significativement l'ACE2 cardiaque dans la CHF expérimentale[37]. Des études expérimentales montrent que les statines bloquent le SRAA, mais augmentent aussi l'expression de l'ACE2 selon Tikoo et al. la protéine ACE2 est environ deux fois plus présente dans le cœur et les reins de lapins athérosclérotiques traités à l'atorvastatine, en lien avec des modifications épigénétiques du gène ACE2[38].
Récepteur de certains coronavirus
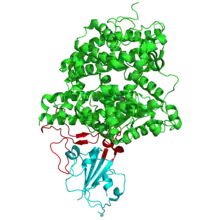
L'ACE2 humaine se lie à l'enveloppe du coronavirus SARS-CoV, responsable du SRAS, à travers la glycoprotéine S du virus[40], qui forme un péplomère à sa surface. Cette protéine est ensuite clivée par une endopeptidase, la cathepsine L, lors du transit vers les endosomes[41] de sorte que le virus pénètre dans la cellule hôte par une endocytose dépendante du pH mais indépendante de la clathrine et des cavéoles[42]. Ce coronavirus dispose également d'une autre possibilité d'entrer dans la cellule par activation de la glycoprotéine S par la protéase transmembranaire à sérine 2 (TMPRSS2) conduisant à la fusion des membranes[43].
La glycoprotéine S du SARS-CoV est composée de deux sous-unités. La sous-unité S1 contient le domaine de liaison au récepteur, noté RBD (pour receptor-binding domain), susceptible de se lier à l'ACE2. Cette liaison provoque des changements conformationnels dans la sous-unité S2 qui facilitent la fusion de l'enveloppe virale avec la membrane plasmique de la cellule hôte[44]. Les résidus d'acides aminés 424 à 494 du domaine de liaison au récepteur forment le motif de liaison au récepteur, noté RBM (pour receptor-binding motif). Parmi les 14 résidus du RBM qui sont en contact direct avec l'ACE2 figurent six résidus de tyrosine qui participent à la reconnaissance spécifique de l'ACE2. Des résidus de cystéine participent également à cette reconnaissance en formant des ponts disulfure.
Les résidus Asn479 et Thr487 du RBM ont une influence sur l'évolution de la maladie du SRAS ainsi que du tropisme du virus SARS-CoV. Le résidu Asn479 est présent dans la plupart des séquences de glycoprotéines S de virus SARS-CoV retrouvés chez l'homme. Toute mutation affectant les résidus 479 et 487 du RBD est susceptible d'induire un changement dans le mode de transmission zoonotique ou interhumaine du SARS-CoV[39] - [45].
Le domaine de liaison au récepteur du virus SARS-CoV de civette masquée présente un résidu de lysine en position 479 qui induit un encombrement stérique et des interférences électrostatiques avec des résidus de l'hélice N-terminale de l'ACE2 humaine, notamment l'His34. Une mutation Lys479 → Asn479 réduit ces blocages et accroît l'affinité entre le RBD de la glycoprotéine S virale et l'ACE2 humaine, ce qui permet une transmission zoonotique. De plus, les ponts salins formé en environnement hydrophobe entre les résidus Lys31 et Glu35 de l'ACE2 humaine ont pour effet de renforcer les interactions de cette dernière avec la glycoprotéine S du virus SARS-CoV. Le résidu Thr487 augmente également l'affinité entre le RBD viral et l'ACE2 humaine. Le groupe méthyle γ de ce résidu de thréonine assure le positionnement du résidu Lys353 de l'ACE2 de telle sorte qu'il forme un pont salin avec le résidu Asp38, ce qui pourrait favoriser la transmission interhumaine du virus[46] - [47].
Un mécanisme semblable a été observé pour le coronavirus SARS-CoV-2[48], responsable du COVID-19. La protéine ACE2 interagit avec la protéine B0AT1/SLC6A19 qui la stabilise sur la membrane plasmique en formant un dimère d'hétérodimères[49], le domaine d'homologie aux collectines de l'ACE2 favorisant la dimérisation. ACE2 joue en outre le rôle de protéine chaperon pour B0AT[50].
Notes et références
- (en) Paul Towler, Bart Staker, Sridhar G. Prasad, Saurabh Menon, Jin Tang, Thomas Parsons, Dominic Ryan, Martin Fisher, David Williams, Natalie A. Dales, Michael A. Patane et Michael W. Pantoliano, « ACE2 X-Ray Structures Reveal a Large Hinge-bending Motion Important for Inhibitor Binding and Catalysis », Journal of Biological Chemistry, vol. 279, no 17, , p. 17996-18007 (PMID 14754895, DOI 10.1074/jbc.M311191200, lire en ligne).
- Les valeurs de la masse et du nombre de résidus indiquées ici sont celles du précurseur protéique issu de la traduction du gène, avant modifications post-traductionnelles, et peuvent différer significativement des valeurs correspondantes pour la protéine fonctionnelle.
- (en) I. Hamming, W. Timens, M. L. C. Bulthuis, A. T. Lely, G. J. Navis et H. van Goor, « Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis », The Journal of Pathology, vol. 203, no 2, , p. 631-637 (PMID 15141377, DOI 10.1002/path.1570, lire en ligne).
- (en) Mary Donoghue, Frank Hsieh, Elizabeth Baronas, Kevin Godbout, Michael Gosselin, Nancy Stagliano, Michael Donovan, Betty Woolf, Keith Robison, Raju Jeyaseelan, Roger E. Breitbart et Susan Acton, « A Novel Angiotensin-Converting Enzyme–Related Carboxypeptidase (ACE2) Converts Angiotensin I to Angiotensin 1-9 », Circulation Research, vol. 87, no 5, , E1-E9 (PMID 10969042, DOI 10.1161/01.res.87.5.e1, lire en ligne).
- (en) Mamfred Boehm et Elizabeth G. Nabel, « Angiotensin-converting enzyme 2 — a new cardiac regulator. », The New England Journal of Medicine, vol. 347, no 22, , p. 1795-1797 (PMID 12456857, DOI 10.1056/NEJMcibr022472, lire en ligne).
- (en) L. Anguiano, M. Riera, J. Pascual et M. J. Soler, « Circulating ACE2 in Cardiovascular and Kidney Diseases », Current Medicinal Chemistry, vol. 24, no 30, (DOI 10.2174/0929867324666170414162841, lire en ligne, consulté le ).
- (en) Gavin Y. Oudit, Michael A. Crackower, Peter H. Backx et Josef M. Penninger, « The Role of ACE2 in Cardiovascular Physiology », Trends in Cardiovascular Medicine, vol. 13, no 3, , p. 93-101 (PMID 12691672, DOI 10.1016/S1050-1738(02)00233-5, lire en ligne).
- (en) Louise M. Burrell, Colin I. Johnston, Christos Tikellis et Mark E. Cooper, « ACE2, a new regulator of the renin–angiotensin system », Trends in Endocrinology and Metabolism, vol. 15, no 4, , p. 166-169 (PMID 15109615, DOI 10.1016/j.tem.2004.03.001, lire en ligne).
- (en) Katalin Úri, Miklós Fagyas, Ivetta Mányiné Siket, Attila Kertész, Zoltán Csanádi, Gábor Sándorfi, Marcell Clemens, Roland Fedor, Zoltán Papp, István Édes, Attila Tóth et Erzsébet Lizanecz, « New Perspectives in the Renin-Angiotensin-Aldosterone System (RAAS) IV: Circulating ACE2 as a Biomarker of Systolic Dysfunction in Human Hypertension and Heart Failure », PLoS One, vol. 9, no 4, , article no e87845 (PMID 24691269, PMCID 3972189, DOI 10.1371/journal.pone.0087845, Bibcode 2014PLoSO...987845U, lire en ligne).
- (en) Shlomo Keidar, Marielle Kaplan et Aviva Gamliel-Lazarovich, « ACE2 of the heart: From angiotensin I to angiotensin (1–7) », Cardiovascular Research, vol. 73, no 3, , p. 463-469 (PMID 17049503, DOI 10.1016/j.cardiores.2006.09.006, lire en ligne).
- (en) Wang Wang, Shaun M.K. McKinnie, Maikel Farhan, Manish Paul, Tyler McDonald, Brent McLean, Catherine Llorens-Cortes, Saugata Hazra, Allan G. Murray, John C. Vederas et Gavin Y. Oudit, « Angiotensin-Converting Enzyme 2 Metabolizes and Partially Inactivates Pyr-Apelin-13 and Apelin-17 », Hypertension, vol. 68, no 2, , p. 365-377 (PMID 27217402, DOI 10.1161/HYPERTENSIONAHA.115.06892, lire en ligne).
- (en) Mohammed A. R. Chamsi-Pasha, Zhili Shao et W. H. Wilson Tang, « Angiotensin-Converting Enzyme 2 as a Therapeutic Target for Heart Failure », Current Heart Failure Reports, vol. 11, no 1, , p. 58-63 (PMID 24293035, PMCID 3944399, DOI 10.1007/s11897-013-0178-0, lire en ligne).
- (en) Annamaria Mascolo, Konrad Urbanek, Antonella De Angelis, Maurizio Sessa, Cristina Scavone, Liberato Berrino, Giuseppe Massimo Claudio Rosano, Annalisa Capuano et Francesco Rossi, « Angiotensin II and angiotensin 1–7: which is their role in atrial fibrillation? », Heart Failure Reviews, vol. 25, no 2, , p. 367-380 (PMID 31375968, DOI 10.1007/s10741-019-09837-7, lire en ligne).
- (en) Peng Zhou, Xing-Lou Yang, Xian-Guang Wang, Ben Hu, Lei Zhang, Wei Zhang, Hao-Rui Si, Yan Zhu, Bei Li, Chao-Lin Huang, Hui-Dong Chen, Jing Chen, Yun Luo, Hua Guo, Ren-Di Jiang, Mei-Qin Liu, Ying Chen, Xu-Rui Shen, Xi Wang, Xiao-Shuang Zheng, Kai Zhao, Quan-Jiao Chen, Fei Deng, Lin-Lin Liu, Bing Yan, Fa-Xian Zhan, Yan-Yi Wang, Geng-Fu Xiao et Zheng-Li Shi, « A pneumonia outbreak associated with a new coronavirus of probable bat origin », Nature, vol. 579, no 7798, , p. 270-273 (PMID 32015507, PMCID 7095418, DOI 10.1038/s41586-020-2012-7, lire en ligne).
- (en) Xintian Xu, Ping Chen, Jingfang Wang, Jiannan Feng, Hui Zhou, Xuan Li, Wu Zhong et Pei Hao, « Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission », Science China Life Sciences, vol. 63, no 3, , p. 457-460 (PMID 32009228, PMCID 7089049, DOI 10.1007/s11427-020-1637-5, lire en ligne).
- Ricki Lewis, « COVID-19 Vaccine Will Close in on the Spikes », sur DNA Science Blog, Public Library of Science (consulté le ).
- Alexandra Walls et al., « Structure, function and antigenicity of the SARS-CoV-2 spike glycoprotein », bioRxiv, (DOI 10.1101/2020.02.19.956581, lire en ligne, consulté le ).
- (en) Junwen Luan, Yue Lu, Xiaolu Jin et Leiliang Zhang, « Spike protein recognition of mammalian ACE2 predicts the host range and an optimized ACE2 for SARS-CoV-2 infection », Biochemical and Biophysical Research Communications, vol. 526, no 1, , p. 165–169 (PMID 32201080, PMCID PMC7102515, DOI 10.1016/j.bbrc.2020.03.047, lire en ligne, consulté le ).
- (en) Anthony R. Fehr et Stanley Perlman, « Coronaviruses: An Overview of Their Replication and Pathogenesis », Coronaviruses, vol. 1282, , p. 1-23 (PMID 25720466, PMCID 4369385, DOI 10.1007/978-1-4939-2438-7_1, lire en ligne).
- (en) Keiji Kuba, Yumiko Imai, Shuan Rao, Hong Gao, Feng Guo, Bin Guan, Yi Huan, Peng Yang, Yanli Zhang, Wei Deng, Linlin Bao, Binlin Zhang, Guang Liu, Zhong Wang, Mark Chappell, Yanxin Liu, Dexian Zheng, Andreas Leibbrandt, Teiji Wada, Arthur S Slutsky, Depei Liu, Chuan Qin, Chengyu Jiang et Josef M Penninger, « A crucial role of angiotensin converting enzyme 2 (ACE2) in SARS coronavirus–induced lung injury », Nature Medicine, vol. 11, no 8, , p. 875-879 (PMID 16007097, PMCID 7095783, DOI 10.1038/nm1267, lire en ligne).
- (en) Anthony J. Turner, Sarah R. Tipnis, Jodie L. Guy, Gillian I. Rice et Nigel M. Hoope, « ACEH/ACE2 is a novel mammalian metallocarboxypeptidase and a homologue of angiotensin-converting enzyme insensitive to ACE inhibitors », Revue canadienne de physiologie et pharmacologie, vol. 80, no 4, , p. 346-353 (PMID 12025971, DOI 10.1139/y02-021, lire en ligne)
- (en) Chad Vickers, Paul Hales, Virendar Kaushik, Larry Dick, James Gavin, Jin Tang, Kevin Godbout, Thomas Parsons, Elizabeth Baronas, Frank Hsieh, Susan Acton, Michael Patane, Andrew Nichols et Peter Tummino, « Hydrolysis of Biological Peptides by Human Angiotensin-converting Enzyme-related Carboxypeptidase », Journal of Biological Chemistry, vol. 277, no 17, , p. 14838-14843 (PMID 11815627, DOI 10.1074/jbc.M200581200, lire en ligne)
- (en) C. M. Ferrario et M. C. Chappell, « Novel angiotensin peptides », Cellular and Molecular Life Sciences, vol. 61, no 21, , p. 2720-2727 (PMID 15549173, DOI 10.1007/s00018-004-4243-4, lire en ligne)
- (en) G.Y. Oudit, A.M. Herzenberg, Z. Kassiri, D. Wong, H. Reich, R. Khokha, M.A. Crackower, P.H. Backx, J.M. Penninger et J.W. Scholey, « Angiotensin-Converting Enzyme-2 (ACE2)—A New Player in the Genesis of Glomerular Injury? », Journal of the American Society of Nephrology, vol. 17, no 10, , p. 2637-2643 (DOI 10.1681/ASN.2006080888, lire en ligne)
- (en) Robson A. S. Santos, Ana C. Simoes e Silva, Christine Maric, Denise M. R. Silva, Raquel Pillar Machado, Insa de Buhr, Silvia Heringer-Walther, Sergio Veloso B. Pinheiro, Myriam Teresa Lopes, Michael Bader, Elizabeth P. Mendes, Virgina Soares Lemos, Maria Jose Campagnole-Santos, Heinz-Peter Schultheiss, Robert Speth et Thomas Walther, « Angiotensin-(1–7) is an endogenous ligand for the G protein-coupled receptor Mas », Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no 14, , p. 8258-8263 (PMID 12829792, PMCID 166216, DOI 10.1073/pnas.1432869100, JSTOR 3139914, Bibcode 2003PNAS..100.8258S, lire en ligne)
- (en) Michael A. Crackower, Renu Sarao, Gavin Y. Oudit, Chana Yagil, Ivona Kozieradzki, Sam E. Scanga, Antonio J. Oliveira-dos-Santos, Joan da Costa, Liyong Zhang, York Pei, James Scholey, Carlos M. Ferrario, Armen S. Manoukian, Mark C. Chappell, Peter H. Backx, Yoram Yagil et Josef M. Penninger, « Angiotensin-converting enzyme 2 is an essential regulator of heart function », Nature, vol. 417, no 6891, , p. 822-828 (PMID 12075344, DOI 10.1038/nature00786, Bibcode 2002Natur.417..822C/abstract, lire en ligne)
- (en) Susan B. Gurley, Alicia Allred, Thu H. Le, Robert Griffiths, Lan Mao, Nisha Philip, Timothy A. Haystead, Mary Donoghue, Roger E. Breitbart, Susan L. Acton, Howard A. Rockman et Thomas M. Coffman, « Altered blood pressure responses and normal cardiac phenotype in ACE2-null mice », The Journal of Clinical Investigations, vol. 116, no 8, , p. 2218-2225 (PMID 16878172, PMCID 1518789, DOI 10.1172/JCI16980, lire en ligne)
- (en) Koichi Yamamoto, Mitsuru Ohishi, Tomohiro Katsuya, Norihisa Ito, Masashi Ikushima, Masaharu Kaibe, Yuji Tatara, Atsushi Shiota, Sumio Sugano, Satoshi Takeda, Hiromi Rakugi et Toshio Ogihara, « Deletion of Angiotensin-Converting Enzyme 2 Accelerates Pressure Overload-Induced Cardiac Dysfunction by Increasing Local Angiotensin II », Hypertension, vol. 47, no 4, , p. 718-726 (PMID 16505206, DOI 10.1161/01.HYP.0000205833.89478.5b, lire en ligne)
- (en) Julie R. Ingelfinger, « ACE2: A New Target for Prevention of Diabetic Nephropathy? », Journal of the American Society of Nephrology, vol. 17, no 11, , p. 2957-2959 (PMID 17021263, DOI 10.1681/ASN.2006090986, lire en ligne)
- (en) Minghao Ye, Jan Wysocki, Josette William, Maria José Soler, Ivan Cokic et Daniel Batlle, « Glomerular Localization and Expression of Angiotensin-Converting Enzyme 2 and Angiotensin-Converting Enzyme: Implications for Albuminuria in Diabetes », Journal of the American Society of Nephrology, vol. 17, no 11, , p. 3067-3075 (PMID 17021266, DOI 10.1681/ASN.2006050423, lire en ligne)
- (en) Sarah R. Tipnis, Nigel M. Hooper, Ralph Hyde, Eric Karran et Gary Christie, « A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase », Journal of Biological Chemistry, vol. 275, no 43, , p. 33238-33243 (PMID 10924499, DOI 10.1074/jbc.M002615200, lire en ligne)
- (en) Slava Epelman, W.H. Wilson Tang, Stephen Y. Chen, Frederick Van Lente, Gary S. Francis et Subha Sen, « Detection of Soluble Angiotensin-Converting Enzyme 2 in Heart Failure: Insights Into the Endogenous Counter-Regulatory Pathway of the Renin-Angiotensin-Aldosterone System », Journal of the American College of Cardiology, vol. 52, no 9, , p. 750-754 (PMID 18718423, PMCID 2856943, DOI 10.1016/j.jacc.2008.02.088, lire en ligne)
- (en) Albena Nunes-Silva, Guilherme Carvalho Rocha, Daniel Massote Magalhaes et Lucas Neves Vaz, « Physical Exercise and ACE2-Angiotensin-(1-7)-Mas Receptor Axis of the Renin Angiotensin System », Protein & Peptide Letters, vol. 24, no 9, (DOI 10.2174/0929866524666170728151401, lire en ligne, consulté le )
- (en) Sumit Kar, Lie Gao et Irving H. Zucker, « Exercise training normalizes ACE and ACE2 in the brain of rabbits with pacing-induced heart failure », Journal of Applied Physiology, vol. 108, no 4, , p. 923–932 (ISSN 8750-7587 et 1522-1601, DOI 10.1152/japplphysiol.00840.2009, lire en ligne, consulté le )
- (en) Shlomo Keidar, Aviva Gamliel-Lazarovich, Marielle Kaplan et Elsa Pavlotzky, « Mineralocorticoid Receptor Blocker Increases Angiotensin-Converting Enzyme 2 Activity in Congestive Heart Failure Patients », Circulation Research, vol. 97, no 9, , p. 946–953 (ISSN 0009-7330 et 1524-4571, DOI 10.1161/01.RES.0000187500.24964.7A, lire en ligne, consulté le )
- (en) Tony Karram, Anan Abbasi, Shlomo Keidar et Eliahu Golomb, « Effects of spironolactone and eprosartan on cardiac remodeling and angiotensin-converting enzyme isoforms in rats with experimental heart failure », American Journal of Physiology-Heart and Circulatory Physiology, vol. 289, no 4, , H1351–H1358 (ISSN 0363-6135 et 1522-1539, DOI 10.1152/ajpheart.01186.2004, lire en ligne, consulté le )
- (en) Kulbhushan Tikoo, Gaurang Patel, Sandeep Kumar et Pinakin Arun Karpe, « Tissue specific up regulation of ACE2 in rabbit model of atherosclerosis by atorvastatin: Role of epigenetic histone modifications », Biochemical Pharmacology, vol. 93, no 3, , p. 343–351 (DOI 10.1016/j.bcp.2014.11.013, lire en ligne, consulté le )
- (en) Fang Li, Wenhui Li, Michael Farzan et Stephen C. Harrison, « Structure of SARS Coronavirus Spike Receptor-Binding Domain Complexed with Receptor », Science, vol. 309, no 5742, , p. 1864-1868 (PMID 16166518, DOI 10.1126/science.1116480, JSTOR 3843779, lire en ligne)
- (en) Yuuki Inoue, Nobuyuki Tanaka, Yoshinori Tanaka, Shingo Inoue, Kouichi Morita, Min Zhuang, Toshio Hattori et Kazuo Sugamura, « Clathrin-Dependent Entry of Severe Acute Respiratory Syndrome Coronavirus into Target Cells Expressing ACE2 with the Cytoplasmic Tail Deleted », Journal of Virology, vol. 81, no 16, , p. 8722-8729 (PMID 17522231, PMCID 1951348, DOI 10.1128/JVI.00253-07, lire en ligne)
- (en) Graham Simmons, Dhaval N. Gosalia, Andrew J. Rennekamp, Jacqueline D. Reeves, Scott L. Diamond et Paul Bates, « Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry », Proceedings of the National Academy of Sciences of the United States of America, vol. 102, no 33, , p. 11876-11881 (PMID 16081529, PMCID 1188015, DOI 10.1073/pnas.0505577102, JSTOR 3376363, Bibcode 2005PNAS..10211876S, lire en ligne)
- (en) Hongliang Wang, Peng Yang, Kangtai Liu, Feng Guo, Yanli Zhang, Gongyi Zhang et Chengyu Jiang, « SARS coronavirus entry into host cells through a novel clathrin- and caveolae-independent endocytic pathway », Cell Research, vol. 18, no 2, , p. 290-301 (PMID 18227861, DOI 10.1038/cr.2008.15, lire en ligne)
- (en) Shutoku Matsuyama, Noriyo Nagata, Kazuya Shirato, Miyuki Kawase, Makoto Takeda et Fumihiro Taguchi, « Efficient Activation of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein by the Transmembrane Protease TMPRSS2 », Journal of Virology, vol. 84, no 24, , p. 12658-12664 (PMID 20926566, PMCID 3004351, DOI 10.1128/JVI.01542-10, lire en ligne)
- (en) Swee Kee Wong, Wenhui Li, Michael J. Moore, Hyeryun Choe et Michael Farzan, « A 193-Amino Acid Fragment of the SARS Coronavirus S Protein Efficiently Binds Angiotensin-converting Enzyme 2 », Journal of Biological Chemistry, vol. 279, no 5, , p. 3197-3201 (PMID 14670965, DOI 10.1074/jbc.C300520200, lire en ligne)
- (en) Lanying Du, Yuxian He, Yusen Zhou, Shuwen Liu, Bo-Jian Zheng et Shibo Jiang, « The spike protein of SARS-CoV — a target for vaccine and therapeutic development », Nature Reviews Microbiology, vol. 7, no 3, , p. 226-236 (PMID 19198616, PMCID 2750777, DOI 10.1038/nrmicro2090, lire en ligne)
- (en) Fang Li, « Structural Analysis of Major Species Barriers between Humans and Palm Civets for Severe Acute Respiratory Syndrome Coronavirus Infections », Journal of Virology, vol. 82, no 14, , p. 6984-6991 (PMID 18448527, PMCID 2446986, DOI 10.1128/JVI.00442-08, lire en ligne)
- (en) Fang Li, Wenhui Li, Michael Farzan et Stephen C. Harrison, « Interactions between SARS coronavirus and its receptor », Advances in Experimental Medicine and Biology, vol. 581, , p. 229-234 (PMID 17037534, DOI 10.1007/978-0-387-33012-9_38, lire en ligne)
- (en) Yushun Wan, Jian Shang, Rachel Graham, Ralph S. Baric et Fang Li, « Receptor Recognition by the Novel Coronavirus from Wuhan: an Analysis Based on Decade-Long Structural Studies of SARS Coronavirus », Journal of Virology, vol. 94, no 7, , article no e00127-20 (PMID 31996437, DOI 10.1128/JVI.00127-20, lire en ligne)
- (en) Renhong Yan, Yuanyuan Zhang, Yaning Li, Lu Xia, Yingying Guo et Qiang Zhou, « Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2 », Science, vol. 367, no 6485, , p. 1444-1448 (PMID 32132184, PMCID 7164635, DOI 10.1126/science.abb2762, lire en ligne)
- (en) Mahmoud Gheblawi, Kaiming Wang, Anissa Viveiros, Quynh Nguyen, Jiu-Chang Zhong, Anthony J. Turner, Mohan K. Raizada, Maria B. Grant et Gavin Y. Oudit, « Angiotensin-Converting Enzyme 2: SARS-CoV-2 Receptor and Regulator of the Renin-Angiotensin System », Circulation Research, vol. 126, no 10, , p. 1456-1474 (PMID 32264791, PMCID 7188049, DOI 10.1161/CIRCRESAHA.120.317015, lire en ligne)