Anatomie des poissons
L'anatomie des poissons est principalement régie par les caractéristiques physiques de l'eau ; elle est beaucoup plus dense que l'air, relativement plus pauvre en oxygène et absorbe plus la lumière que l'air.
Système digestif

Chez les poissons, la nourriture est ingérée par la bouche et subit un début de trituration dans l'œsophage. Le broyage des aliments se fait essentiellement dans l'estomac et, chez de nombreuses espèces, dans des diverticules en forme de doigt appelés cæca pyloriques. Ces caeca sécrètent des enzymes digestives et commencent l'absorption des nutriments.
Certains poissons comme la carpe ou le koï ne possèdent pas d'estomac anatomique.
Des organes comme le foie et le pancréas apportent de nouvelles enzymes digestives au fur et à mesure que la nourriture progresse dans le tube digestif. Les intestins complètent le processus de digestion et d'absorption des nutriments. Les poissons mangent des plantes et d'autres organismes.
Système respiratoire
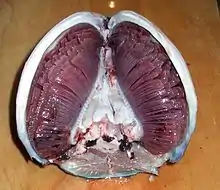

La plupart des poissons pratiquent usuellement la respiration aquatique à travers leurs branchies. Néanmoins, certains poissons sont capables de pratiquer la respiration aérienne en annexe lorsque le milieu aquatique devient hypoxique, tandis que d'autres poissons possèdent de véritables poumons et doivent respirer de l'air périodiquement pour ne pas suffoquer.
Poissons à respiration strictement aquatique
Les poissons respirent généralement grâce à des branchies localisées de part et d'autre du pharynx. Les branchies sont constituées de filaments contenant un réseau de capillaires sanguins, dont la grande surface développée facilite l'échange du dioxygène et du dioxyde de carbone dissous dans l'eau. Sur le côté opposé aux filaments branchiaux, les branchiospines (ou branchicténie) sont des projections osseuses, digitiformes, de l'arc branchial et qui servent à retenir les proies.
Les poissons pompent l'eau (contenant du dioxygène) par la bouche, puis la font circuler sur les branchies. Le sang dans les capillaires sanguins s'écoule dans la direction opposée à celle de l'eau, permettant ainsi un échange à contre-courant ; l'eau appauvrie en dioxygène est ensuite expulsée par les ouvertures situées latéralement par rapport au pharynx.
Chez la plupart des poissons cartilagineux (hormis les chimères), les fentes branchiales sont visibles (latéralement chez les requins et les lamproies, ventralement chez les raies). Ces fentes branchiales sont cachées chez les poissons osseux par un opercule (osseux lui-aussi) s'ouvrant postérieurement par une fente appelée ouïe.
Poissons à respiration aérienne
De nombreux poissons peuvent respirer de l'air, grâce à différents mécanismes.
- Respirer par la peau :
- La peau des anguilles peut être utilisée pour absorber l'oxygène.
- Les périophthalmes respirent en absorbant l'oxygène à travers la peau, de façon similaire aux grenouilles.
- Un « organe de respiration annexe », utilisé pour extraire l'oxygène de l'air :
- La cavité buccale des anguilles électriques peut être utilisée pour respirer de l'air.
- Les gouramis, ainsi que les familles de poissons-chats Channidae, Luciocephalidae et Clariidae ont un « organe labyrinthe » au-dessus des branchies pour cela[1].
- Les Cobitidae, les trahiras et certains poissons-chats des familles Loricariidae, Callichthyidae et Scoloplacidae peuvent absorber l'air à travers leur système digestif[1].
- Certaines espèces, tels les dipneustes, possèdent des poumons plus ou moins rudimentaires qui leur permettent de survivre en respirant hors de l'eau dans des mares appauvries en oxygène ou régulièrement asséchées. Les semionotiformes et le poisson-castor ont une vessie natatoire vascularisée utilisée de la même manière.
La faculté de respirer de l'air est surtout utile aux poissons habitant des eaux peu profondes où la concentration en oxygène peut baisser à certains moments de l'année. Pendant ces périodes, les poissons qui dépendent uniquement de l'oxygène contenu dans l'eau (comme les perches) suffoquent rapidement tandis que les poissons pouvant respirer de l'air survivent mieux, même dans une eau qui ressemble davantage à de la boue humide. Dans des cas extrêmes, certains de ces poissons peuvent survivre plusieurs semaines dans des replis humides, à des endroits où l'eau s'est presque complètement retirée, dans un état d'estivation.
Squelette
Le squelette du poisson est constitué du neurocrâne et de la colonne vertébrale. Les vertèbres, peu spécialisées, ont des développements latéraux qui portent les côtes ou arêtes.
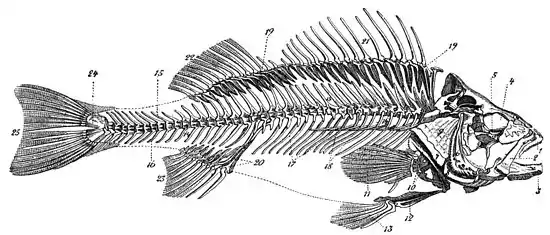
1/Mâchoire supérieure, 2/Arc hyoïde 3/Mâchoire inférieure, 4/orbite oculaire , 5/Suborbital, 6/ Préopercule branchial, 7/ Interoperculaire, 8/Suboperculaire , 9/Opercule branchial, 10/Rayons de ceinture scapulaire, 11/Nageoire pectorale, 12/Ceinture pelvienne 13/ Rayons de nageoire pelvienne, 14/Colonne vertébrale, 15/Épine neurale (apophyse vertébrale supérieure), 16/Épine hémale (apophyse vertébrale inférieure), 17/Côte ou arête ventrale, 18/Fausse côte, 19/Ptérygophore, 20/Cartilage radial, 21/Rayons de la nageoire dorsale antérieure, 22/Rayons de la nageoire dorsale postérieure, 23/Rayons de la nageoire anale, 24/Hypural, 25/Rayons de nageoire caudale.
Système circulatoire
Comme tous les Vertébrés, les poissons ont un système circulatoire clos comportant une pompe cardiaque. Le circuit sanguin forme une boucle unique : le sang va du cœur aux branchies en passant par les arcs aortiques, puis part irriguer le reste du corps avant de revenir au cœur, à l'intérieur duquel ne circule que du sang désoxygéné.
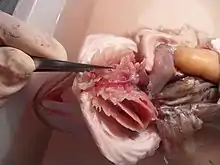
Chez la plupart des poissons, l'oxygénation du cœur par les coronaires est limité à l'épicarde[2], la couche externe du cœur, et l'irrigation des tissus cardiaques se fait principalement par le sang contenu au sein de la cavité cardiaque. Le cœur est donc irrigué uniquement par du sang désoxygéné, ce qui peut conduire à une hypoxie cardiaque lorsque les tissus de l'organisme présentent une trop forte demande en oxygène (comme lors d'un intense effort physique).
Le cœur des poissons est généralement divisé en quatre parties :
- le sinus veineux qui collecte le sang veineux et le dirige vers l'atrium ;
- l'atrium, poche musculaire à sens unique qui dirige le sang vers le ventricule ;
- le ventricule cardiaque, poche musculaire qui effectue le travail de pompage cardiaque ;
- le bulbe cardiaque qui, relié à l'artère ventrale aorte, permet l'éjection du sang vers les branchies en passant par des arcs aortiques (4 paires irriguant les 4 paires de branchies).
Même s'il comporte quatre parties, le cœur d'un poisson comporte deux compartiments[3]
Chez les poissons pulmonés, le cloisonnement du cœur est imparfait, seul l'atrium est cloisonné. L'absence de cloisonnement au niveau du ventricule (comme c'est le cas chez les mammifères), est compensé par la présence d'une lame spirale qui évite le mélange du sang oxygéné (poumons) et du sang désoxygéné (organes).
Système excréteur
Certains déchets métaboliques peuvent diffuser, directement à travers la peau et les branchies, du sang vers l'eau environnante. Les autres sont éliminés par les reins, organes excréteurs qui filtrent les déchets pour les éliminer du sang. Comme de nombreux animaux aquatiques, les poissons rejettent leurs déchets azotés sous forme d'ions ammonium (NH4+). L'anse du néphron est absente chez les poissons. Certains poissons marins sont aglomérulaires, c'est-à-dire qu'ils ne possèdent aucune capsule glomérulaire (ou capsule de Bowman) où la filtration des ions se fait car leur milieu aquatique est hypertonique. La filtration d'ions est alors moins importante dans ce milieu.
Les poissons d'eau de mer ont tendance à perdre de l'eau à cause des phénomènes d'osmose et les reins concentrent les déchets afin de réabsorber l'eau au maximum, puisqu'ils sont hypotoniques par rapport à leur milieu hypertonique. Il se passe l'inverse chez les poissons d'eau douce, qui ont tendance à absorber de l'eau par osmose. Les reins produisent alors de grandes quantités d'urine très diluée, puisqu'ils sont hypertoniques par rapport à leur environnement hypotonique.
Certains poissons qui migrent du milieu salé ou milieu dulçaquicole (anadromes, catadromes) ont des reins capables de s'adapter au milieu et de changer leur mode de fonctionnement.
Système nerveux et sensoriel
Les poissons ont un système nerveux bien développé qui contient un cerveau divisé en plusieurs parties :
- le bulbe olfactif, dans la partie la plus antérieure, est impliqué dans le sens de l'odorat ;
- le télencéphale qui, contrairement à celui des autres vertébrés, s'occupe presque uniquement du traitement des informations concernant le sens de l'odorat;
- les lobes optiques qui constituent la partie la plus développée traitent des informations sensorielles et enclenchent la commande motrice ;
- le cervelet coordonne les mouvements du corps ;
- le bulbe rachidien contrôle le fonctionnement des organes internes.
La moelle épinière s'insinue dans la colonne vertébrale dans le prolongement du bulbe rachidien.

La plupart des poissons possèdent des organes sensoriels très développés. La majorité des poissons diurnes ont des yeux capables d'une vision des couleurs au moins aussi bonne que celle des humains. Les poissons disposent aussi de cellules rétiniennes en cônes spécialisées dans la perception de l'ultraviolet ou du proche-UV et de la polarisation de la lumière subaquatique[4]; ils ont une vision polarisée[4].
De nombreux poissons possèdent aussi des cellules spécialisées appelées chémorécepteurs responsables d'un extraordinaire sens du goût et de l'odorat.
Dotés d'oreilles internes et capables d'émettre des sons, entre autres grâce à leur vessie natatoire, les poissons peuvent en percevoir entre 20 et 3 000 hertz[5].
La plupart des poissons possèdent des récepteurs sensitifs formant la ligne latérale, qui leur permet de détecter les courants faibles et les vibrations (par exemple le mouvement de leur proie)[6]. Les poissons-chats et les requins possèdent de plus des organes qui détectent des courants électriques de faible intensité. Certaines espèces — telles les mormyridés, notamment Gnathonemus petersii — peuvent être utilisées pour détecter la pollution de l'eau grâce à la variation de leurs émissions électriques. Des chercheurs de l'université d'Édimbourg (Écosse) travaillant sur la truite arc-en-ciel ont découvert en 2003 que les poissons pouvaient ressentir la douleur[7]. Cette étude a été critiquée par James D. Rose la même année[8], au motif qu'il manque aux poissons un néocortex pour ressentir la douleur (à la manière de l'humain). Un article datant de la même année avançait que les poissons ne ressentaient pas la douleur, mais réagissaient simplement aux stimuli[9].
Système immunitaire
Les types d'organes immunitaires varient selon les types de poissons[10]:
- Chez les Agnathes (myxines et lamproies), il n'existe pas de véritables organes immunitaires. En revanche, ces poissons possèdent des tissus lymphoïdes, capables de produire des cellules immunitaires, dans leurs organes. Par exemple, les érythrocytes, macrophages et plasmocytes sont produits dans le rein antérieur (ou pronéphros), et certaines zones de l'intestin (où les granulocytes font leur maturation) chez la myxine ressemble à une moelle osseuse primitive.
- Les poissons cartilagineux (requins et raies) ont un système immunitaire plus évolué que les Agnathes. Ils possèdent trois organes spécialisés qui sont typiques des Chondrichtyens :
- les organes épigonaux, entourant les gonades, qui ressemblent à la moelle osseuse des mammifères ;
- l'organe de Leydig, à l'intérieur des parois de l'œsophage ;
- une valve spirale dans l'intestin.
Tous ces organes contiennent des cellules immunitaires typiques (granulocytes, lymphocytes et plasmocytes). Les Chondrichtyens possèdent aussi un thymus identifiable et une rate bien développée (qui est leur organe immunitaire le plus important) où divers lymphocytes, plasmocytes et macrophages se développent et sont stockés.
- Les Chondrostéens (esturgeons et Polyodontidae) et les polyptères possèdent un site principal de production de granulocytes dans une masse cellulaire associée aux méninges, membranes entourant le système nerveux central), et leur cœur est souvent couvert d'un tissu qui contient des lymphocytes, des fibroblastes et quelques macrophages. Le rein des Chondrostéens est un organe hématopoïétique important où les érythrocytes, granulocytes lymphocytes et macrophages se développent ;
- Les Téléostéens, ou poissons osseux ont, comme les Chondrostéens, leurs principaux tissus immunitaires dans les reins (surtout dans le rein antérieur, où l'on trouve de nombreuses cellules immunitaires de type varié)[11]. Les Téléostéens possèdent de plus un thymus, une rate et des zones immunitaires éparses intégrées dans des tissus muqueux (par exemple dans la peau, les branchies, l'intestin et les gonades). On pense que comme chez les mammifères, leurs érythrocytes et granulocytes (notamment les neutrophiles) résident dans la rate, alors que les lymphocytes sont majoritaires dans le thymus[12] - [13]. D'après une découverte récente non encore confirmée, un système lymphatique similaire à celui des mammifères a été décrit chez un Téléostéen, le poisson zèbre (Brachydanio rerio). Ce système serait l'endroit où les lymphocytes T s'accumuleraient en attendant d'être mis en contact avec un antigène[14].
Température corporelle

La plupart des poissons sont des animaux dits à sang froid (poïkilothermes). Plus exactement, leur température interne varie avec celle du milieu (ectothermes).
Des études récentes montrent que certaines espèces prédatrices sont capables de maintenir leur température interne au-dessus de celle de l'eau. Cette aptitude a été démontrée chez certains téléostéens — Scombroïdés (marlin, espadon, voilier), gastéro (Gasterochisma melampus) — et chondrostéens — Lamnidés (requin mako, requin blanc, requins taupes). Leur capacité à réguler leur température varie de l'espadon, qui ne réchauffe que ses yeux et son cerveau, au thon bleu et aux requins taupes capables d'élever leur température corporelle de 20 °C par rapport à celle de l'eau. Des retes mirabiles constituent une sorte d'échangeur thermique et permettent de récupérer les calories du sang chaud et pauvre en oxygène. Le sang qui arrive aux organes est donc relativement chaud tout en diminuant le coût énergétique. Cette méthode accroît la force contractile des muscles, augmente le rendement de la digestion et du système nerveux central, et permet par exemple aux thonidés de chasser en eau profonde durant la nuit et de capturer facilement sardines et maquereaux engourdis par le froid.
Système reproducteur

Il existe différents modes de reproduction : l'oviparité, l'ovoviviparité et la viviparité.
Chez les poissons ovipares, après une fécondation généralement externe, la femelle pond ses œufs, et les embryons se développent et éclosent en dehors de son corps. Les embryons sont nourris grâce aux réserves contenues dans l'œuf. Plus de 97 % des espèces connues de poissons sont ovipares[15], dont par exemple les saumons, les poissons rouges, les thons, les anguilles, etc. Il existe différentes techniques de ponte :
- abandon des œufs au hasard ;
- dépôt des œufs dans un nid construit ou un abri naturel ;
- suspension des œufs à un support ;
- dépôt puis mise à l'abri des œufs dans la bouche d'un des parents ;
- enfouissage des œufs dans les sédiments.
La fécondation est le plus souvent externe, les gamètes se mélangeant à proximité des deux poissons. Chez quelques poissons, la fécondation est interne, le mâle utilisant un organe intromittant pour déposer le sperme dans le cloaque de la femelle. Il s'agit surtout des requins ovipares comme le requin dormeur cornu et des raies ovipares (comme Rajidae). Dans ce cas, le mâle possède deux ptérygopodes, mutations de la nageoire pelvienne.
Les jeunes poissons sortant de l'œuf sont appelés larves. Elles sont généralement peu formées, transportent une vésicule vitelline (ou sac vitellin) contenant leurs réserves nutritives et ont une apparence très différente des adultes. La période larvaire des poissons ovipares est généralement courte (quelques semaines), les larves grandissant rapidement tandis que leur structure et leur apparence change (la métamorphose) pour ressembler aux jeunes de leur espèce. Pendant cette période, les larves se nourrissent d'abord de leurs réserves puis du zooplancton ; ce processus dépend de la concentration en zooplancton et peut causer une forte mortalité parmi les larves.
Chez les poissons ovovivipares, les œufs restent dans le corps de la mère après fécondation interne. Chaque embryon se développe dans son œuf, sans utiliser de réserves nutritives de la mère, puis sort du corps de sa mère lors de l'éclosion. Parmi les poissons ovovivipares, on peut noter les guppys, les platys, les xiphos, les mollys, les squatiniformes et les cœlacanthes.
Chez les poissons vivipares, la fécondation est aussi interne mais chaque embryon reçoit ses nutriments du corps de sa mère, et non des réserves de l'œuf. Les petits sont mis au monde par accouchement. Les poissons vivipares ont une structure similaire au placenta des mammifères, reliant le sang de la mère et celui de l'embryon. Parmi les poissons vivipares, on peut noter les Embiotocidae, les Goodeidae et les requins citrons. Les embryons de certains poissons vivipares ont un comportement appelé oophagie, lorsque l'embryon en développement mange les œufs produits par la mère. Ce comportement a été observé chez certains requins comme le requin mako ou le maraîche mais aussi chez d'autres poissons comme le Nomorhamphus ebrardtii[16]. Le cannibalisme in utero est un comportement encore plus inhabituel, lorsque l'embryon le plus grand mange ses frères plus petits et plus faibles. Ce comportement a été observé chez des requins comme Carcharias taurus mais aussi avec des espèces comme Nomorhamphus ebrardtii[16].
Système de flottaison
Puisque les tissus corporels sont plus denses que l'eau, les poissons doivent compenser les effets de la gravité pour éviter de couler. De nombreux téléostéens ont un organe interne appelé vessie natatoire qui ajuste leur flottabilité en réduisant ou augmentant le volume de gaz qu'elle contient, selon le même système que les ballasts des sous-marins.
Annexes
Bibliographie
- Edward C. Migdalski et George S. Fichter (trad. Charles Roux, ill. Norman Weaver), Les Poissons du monde, Paris, Odège, , 316 p. (ISBN 2-245-01149-X et 9782245011492)
- Karel Pivnicka et Karel Cerny, Poissons, Paris, Nathan, , 304 p. (ISBN 2-7000-1518-5)
- John R. Paxton, William N. Eschmeyer et J.-J. Vayne (trad. Marc Baudoux), Les poissons, Bordas, coll. « Encyclopédie des animaux », (ISBN 2-04-027019-1)
- (en) J. S. Nelson, Fishes of the World, John Wiley & Sons, (ISBN 0-471-25031-7)
- (en) G. Helfman, Collette B. et D. Facey, The Diversity of Fishes, Malden, Mass. etc., Blackwell Publishing, , 528 p. (ISBN 0-86542-256-7)
Liens externes
Références
- Cet article est partiellement ou en totalité issu de l'article intitulé « Poisson » (voir la liste des auteurs).
- Jonathan W. Armbruster, « Modifications of the Digestive Tract for Holding Air in Loricariid and Scoloplacid Catfishes », dans Copeia, Vol. 3, 1998, pp. 663 - 675 [lire en ligne].
- Roger Eckert, David Randall, Warren Burggren, Kathleen French, François Math, Alain Propper, Louis Henquell. Physiologie animale: Mécanismes et adaptations. De Boeck Université, 1999, 467-516 pp. (ISBN 2744500534)
- John F. Setaro, Circulatory System, Microsoft Encarta 99.
- Hawryshyn C.W (1992) Polarization vision in fish. American Scientist, 164-175.
- Science & Vie n° 1134 (mars 2012), page 89.
- James Orr, Fish, Microsoft Encarta 99.
- Alex Kirby, Fish do feel pain, scientists say, BBC News, 30 avril 2003 [lire en ligne].
- James D; Rose, A critique of the paper: “Do fish have nociceptors: Evidence for the evolution of a vertebrate sensory system”, 2003 [lire en ligne].
- Michel Tarragnat, Les poissons souffrent-il ?, 14 février 2003, [lire en ligne].
- A.G. Zapata, A. Chiba et A. Vara, « Cells and tissues of the immune system of fish » dans The Fish Immune System: Organism, Pathogen and Environment, Fish Immunology Series (éds. G. Iwama et T.Nakanishi), New York, Academic Press, 1996, pages 1-55.
- D.P. Anderson, Fish Immunology (éds. S.F. Snieszko et H.R. Axelrod), Hong Kong: TFH Publications, 1977.
- S. Chilmonczyk, The thymus in fish: development and possible function in the immune response, dans Annual Review of Fish Diseases, Volume 2, 1992, pages 181-200.
- J.D. Hansen et A.G. Zapata, Lymphocyte development in fish and amphibians dans Immunological Reviews, Volume 166, 1998, pages 199-220.
- Kucher et al., Development of the zebrafish lymphatic system requires VegFc signalling, dans Current Biology, Volume 16, 2006, pages 1244-1248.
- Peter Scott, Livebearing Fishes, p. 13. Tetra Press, 1997 (ISBN 1-5646-5193-2).
- A. Meisner et J. Burns, « Viviparity in the Halfbeak Genera Dermogenys and Nomorhamphus (Teleostei: Hemiramphidae) » dans Journal of Morphology 234, pp 295-317, 1997.