Protéine membranaire périphérique
Une protéine membranaire périphérique est une protéine membranaire qui n'adhère que temporairement à la membrane biologique à laquelle elle est associée. Ces protéines peuvent se lier aux protéines membranaires intégrales ou pénétrer les régions périphériques de la bicouche lipidique. Les sous-unités protéiques régulatrices de nombreux canaux ioniques et récepteurs transmembranaires, par exemple, sont des protéines membranaires périphériques. Contrairement aux protéines membranaires intégrales, les protéines membranaires périphériques ont tendance à s'accumuler dans la fraction soluble dans l'eau de toutes les protéines extraites par une procédure de purification des protéines. Les protéines qui ont un ancrage GPI font exception à cette règle et peuvent avoir des propriétés de purification semblables à celles des protéines membranaires intégrales.
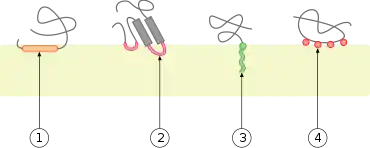
L'attachement réversible des protéines aux membranes biologiques régule la signalisation cellulaire et plusieurs autres phénomènes cellulaires importants en faisant intervenir des mécanismes variés[1]. Par exemple, l'association étroite de nombreuses enzymes avec des membranes biologiques peut les rapprocher de leurs substrats lipidiques[2]. La liaison membranaire peut également favoriser des réarrangements, des dissociations ou des changements conformationnels au sein de nombreux domaines structuraux protéiques, entraînant l'activation de leur activité biologique[3] - [4]. De nombreuses protéines se positionnent de surcroît sur les feuillets interne ou externe de leur membrane d'attache[5], ce qui facilite l'assemblage de complexes protéiques (en) en accroissant la probabilité que s'établissent les interactions protéine-protéine appropriées.
Liaison à la bicouche lipidique
Les protéines membranaires périphériques peuvent interagir directement avec la bicouche lipidique ou par l'intermédiaire d'autres protéines. Dans le premier cas, on parle de protéines amphitropiques[3]. Certaines protéines, telles que les protéines G et certaines protéine kinases, interagissent simultanément avec la bicouche lipidique et des protéines transmembranaires. Certains peptides antimicrobiens, hormones peptidiques et neurotoxines s'accumulent à la surface de la membrane avant de se rassembler et d'interagir avec leurs récepteurs de surface cellulaire cibles, qui peuvent eux-mêmes être des protéines membranaires périphériques.
La bicouche phospholipidique qui forme la membrane plasmique se compose d'une région centrale hydrophobe prise en sandwich entre deux régions hydrophiles, une sur la face intérieure et l'autre sur la face extérieure de la membrane plasmique. On a montré que les surfaces internes et externes, ou régions interfaciales, de modèles de bicouches de phospholipides ont une épaisseur d'environ 0,8 à 1 nm, bien que cette épaisseur puisse être plus importante dans les membranes biologiques qui contiennent de grandes quantités de gangliosides ou de lipopolysaccharides[7]. La région centrale interne hydrophobe de membranes biologiques typiques peut avoir une épaisseur d'environ 2,7 à 3,2 nm, estimée par diffusion des rayons X aux petits angles (SAXS)[8]. La limite entre la région centrale hydrophobe et les régions interfaciales hydrophiles est très étroite, avec une épaisseur d'environ 0,3 nm. Lorsqu'on quitte la région centrale hydrophobe vers les régions hydrophiles interfaciales, la concentration d'eau effective varie brutalement à travers cette couche limite, passant de presque zéro à une concentration d'environ 2 mol·L-1[9] - [10]. Les groupes phosphate dans les bicouches phospholipidiques sont entièrement hydratés ou saturés d'eau et sont situés à environ 0,5 nm en dehors de la limite de la région centrale hydrophobe[11].
Certaines protéines hydrosolubles s'associent irréversiblement aux bicouches lipidiques et peuvent former des canaux transmembranaires en hélices α ou tonneaux β. De telles transformations surviennent avec les toxines formant des pores (en), comme la colicine A et l'α-hémolysine. Elles peuvent également se produire pour la Bcl-2–associated X protein, pour certains peptides antimicrobiens amphiphiles et pour certaines annexines. Ces protéines sont généralement décrites comme périphériques car l'un de leurs états conformationnels est soluble dans l'eau ou n'est associé à une membrane que de façon lâche[12].
Mécanismes de liaison à la membrane
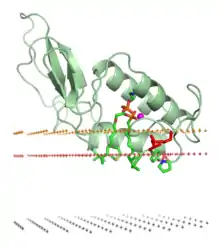
L'association d'une protéine à une bicouche lipidique peut entraîner des changements conformationnels importants affectant la structure tertiaire de la protéine. Il peut s'agir du repliement d'un domaine précédemment déplié ou d'un réarrangement du repliement du domaine de la protéine interagissant avec la membrane. Il peut également s'agir d'une modification de la structure quaternaire, avec le départ ou l'arrivée d'une ou plusieurs sous-unités ou d'oligomères peptidiques, et faire intervenir la liaison spécifique à des ions, des ligands ou des molécules de phosphatidylinositol.
Les protéines amphitropiques doivent généralement interagir fortement avec la bicouche lipidique pour être biologiquement actives. Il peut s'agir du traitement enzymatique de lipides ou d'autres substances hydrophobes, l'ancrage sur la membrane, et la liaison et le transfert de petites molécules apolaire entre différentes membranes cellulaires. Ces protéines peuvent être ancrées à la bicouche par l'intermédiaire d'interactions hydrophobes entre les lipides de la bicouche et les chaînes latérales apolaires des résidus d'acides aminés de la surface de ces protéines, par des liaisons non covalentes particulières avec des lipides régulateurs, ou par ancrage covalent à des lipides membranaires.
La composition lipidique de la membrane conditionne son affinité pour de nombreuses protéines périphériques qui lui sont associées[14].
Association hydrophobe non spécifique
Les protéines amphitropiques s'associent aux bicouches lipidiques via diverses structures d'ancrage hydrophobes. Tels que des hélices α amphiphiles, des boucles apolaires exposées, des résidus d'acides aminés acylés ou lipidés par modification post-traductionnelle, ou des chaînes acyles de lipides régulateurs spécifiquement liés tels que des phosphatidylinositol phosphates. On a montré que les interactions hydrophobes sont importantes même pour des peptides et protéines fortement cationiques, comme le domaine polybasique de la protéine MARCKS (en) ou de l'histactophiline, lorsque leurs ancres hydrophobes naturelles sont présentes[15].
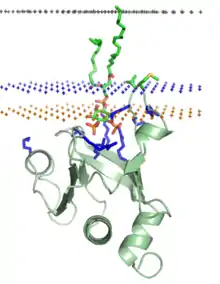
Liaison covalente à des ancres lipidiques
Du côté cytosolique de la membrane plasmique, les protéines à ancrage lipidique sont liées par covalence à différentes chaînes acyle d'acides gras par palmitylation, myristoylation ou prénylation, tandis que, sur la face extérieure de la membrane plasmique, ces protéines sont liées par covalence à des molécules de glycosylphosphatidylinositol et de cholestérol[16] - [17]. L'association des protéines aux membranes biologiques par l'intermédiaire de résidus acylés est un processus réversible car la chaîne acyle peut se retrouver enfouie dans la poche de liaison hydrophobe d'une protéine après sa dissociation de la membrane. Ce processus se produit au sein des sous-unités bêta des protéines G. Les ancres lipidiques sont généralement liées à des segments très flexibles des protéines, qu'il est difficile de réoudre par cristallographie aux rayons X.
Liaison protéine-lipide spécifique
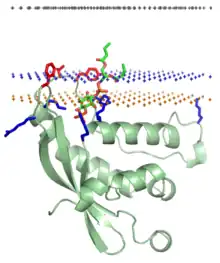
Certaines protéines cytosoliques sont recrutées sur des membranes plasmiques par reconnaissance de certains types de lipides spécifiques à ces membranes[19]. La liaison d'une protéine à un lipide spécifique fait intervenir des domaines structurels particuliers ciblant les membranes plasmiques contenant ce lipide, dont la région hydrophile se lie à des poches de liaison spécifiques sur la protéine. Il s'agit d'une interaction protéine–ligand stabilisée par la formation de liaisons hydrogène, des liaisons de van der Waals et des liaisons hydrophobes entre la protéine et le ligand lipidique. Il s'établit également des liaisons ioniques entre les résidus d'aspartate ou de glutamate des protéines et les groupes phosphate des lipides via des cations de calcium Ca2+. De telles liaisons ioniques s'établissent lorsque les protéines en solution sont déjà liées à des cations Ca2+, avant de se lier aux lipides ; c'est par exemple le cas des domaines C2 (en) et des annexines.
Interactions protéine-lipide électrostatiques
Toute protéine chargée positivement est attirée par une membrane chargée négativement sous l'effet d'interactions électrostatiques non spécifiques. Cependant, toutes les protéines périphériques ne sont pas chargées positivement et les membranes ne sont chargées négativement que sur certaines surfaces, comme le côté cytosolique des membranes plasmiques et le feuillet externe des membranes bactériennes externes et des membranes mitochondriales externes. Les interactions électrostatiques jouent par conséquent un rôle dans le ciblage membranaire de transporteurs d'électrons comme le cytochrome c, de toxines cationiques comme le charybdotoxine, et de domaines de ciblage membranaire spécifiques comme le domaine PH (en), le domaine C1 (en) et le domaine C2 (en).
Les interactions électrostatiques sont très dépendantes de la force ionique de la solution. Elles sont relativement faibles à la force ionique physiologique (0,14 mol L−1 de NaCl), équivalant à environ 13 à 17 kJ mol−1 pour les petites protéines cationiques comme le cytochrome c, la charybdotoxine ou l'hisactophiline[15] - [20] - [21].
Géométrie de l'insertion dans les membranes
On peut étudier l'orientation et la profondeur d'insertion de nombreux peptides et protéines amphitropiques dans les membranes par site-directed spin labeling (en)[22] (SDSL), marquage chimique, mesure de l'affinité de liaison à la membrane de protéines obtenues par mutagenèse[23], spectroscopie de fluorescence[24], spectroscopie RMN en solution ou à l'état solide[25], spectroscopie infrarouge à transformée de Fourier et réflectance totale atténuée[26] (ATR FTIR), cristallographie aux rayons X ou par diffraction de neutrons[27] et diverses méthodes de modélisation numérique[28] - [29] - [30].
Deux modes d'association protéine-membrane ont été identifiés. Les protéines solubles dans l'eau n'ont généralement pas de résidu d'acide aminé apolaire ni d'ancrage hydrophobe, de sorte qu'elles demeurent entièrement en solution aqueuse et ne pénètrent pas dans la bicouche lipidique. De telles protéines n'interagissent avec les lipides membranaires qu'à travers des interactions électrostatiques, comme l'illustrent les ribonucléases et la polylysine. Cependant, les protéines amphitropiques présentent généralement à leur surface des ancres hydrophobes susceptibles de pénétrer dans le cœur hydrophobe de la bicouche lipidique. Ces protéines « déforment » la bicouche lipidique, ce qui abaisse la température de transition liquide-gel de cette dernière[31]. La formation d'une liaison de ce type est un phénomène généralement fortement exothermique[32]. L'association d'hélices α amphiphiles avec des membranes se produit de la même manière[27] - [33]. Des protéines intrinsèquement désordonnées ou non repliées présentant des résidus non polaires ou des ancres lipidiques peuvent également pénétrer la région interfaciale de la membrane et atteindre le cœur hydrocarboné, notamment lorsque ces peptides sont cationiques et interagissent avec des membranes chargées négativement[34] - [35] - [36].
Notes et références
- (en) David S. Cafiso, « Structure and interactions of C2 domains at membrane surfaces », Protein-Lipid Interactions: From Membrane Domains to Cellular Networks, édité par L. K. Tamm, John Wiley & Sons, Chichester, 2005, p. 403–422. (ISBN 3-527-31151-3)
- (en) Moumita Ghosh, Dawn E. Tucker, Scott A. Burchett et Christina C. Leslie, « Properties of the Group IV phospholipase A2 family », Progress in Lipid Research, vol. 45, no 6, , p. 487-510 (PMID 16814865, DOI 10.1016/j.plipres.2006.05.003, lire en ligne)
- (en) Joanne E. Johnson et Rosemary B. Cornell, « Amphitropic proteins: regulation by reversible membrane interactions (Review) », Molecular Membrane Biology, vol. 16, no 3, , p. 217-235 (PMID 10503244, DOI 10.1080/096876899294544, lire en ligne)
- (en) Guruvasuthevan R. Thuduppathy, Jeffrey W. Craig, Victoria Kholodenko, Arne Schon et R. Blake Hill, « Evidence that Membrane Insertion of the Cytosolic Domain of Bcl-xL is Governed by an Electrostatic Mechanism », Journal of Molecular Biology, vol. 359, no 4, , p. 1045-1058 (PMID 16650855, PMCID 1785297, DOI 10.1016/j.jmb.2006.03.052, lire en ligne)
- (en) Satoshi Takida et Philip B. Wedegaertner, « Exocytic pathway‐independent plasma membrane targeting of heterotrimeric G proteins », FEBS Letters, vol. 567, nos 2-3, , p. 209-213 (PMID 15178324, DOI 10.1016/j.febslet.2004.04.062, lire en ligne)
- (en) Kathryn M. Ferguson, Mark A. Lemmon, Joseph Schlessinger et Paul B. Sigler, « Structure of the high affinity complex of inositol trisphosphate with a phospholipase C pleckstrin homology domain », Cell, vol. 83, no 6, , p. 1037-1046 (PMID 8521504, DOI 10.1016/0092-8674(95)90219-8, lire en ligne)
- (en) Thomas J. McIntosh, Adriana Vidal et Sidney A. Simon, « The energetics of peptide-lipid interactions: Modulation by interfacial dipoles and cholesterol », Current Topics in Membranes, vol. 52, , p. 309-338 (DOI 10.1016/S1063-5823(02)52013-5, lire en ligne)
- (en) Kakoli Mitra, Iban Ubarretxena-Belandia, Tomohiko Taguchi, Graham Warren et Donald M. Engelman, « Modulation of the bilayer thickness of exocytic pathway membranes by membrane proteins rather than cholesterol », Proceedings of the National Academy of Sciences of the United States of America, vol. 101, no 12, , p. 4083-4088 (PMID 15016920, PMCID 384699, DOI 10.1073/pnas.0307332101, Bibcode 2004PNAS..101.4083M, lire en ligne)
- (en) Derek Marsh, « Polarity and permeation profiles in lipid membranes », Proceedings of the National Academy of Sciences of the United States of America, vol. 98, no 14, , p. 7777-7782 (PMID 11438731, PMCID 35418, DOI 10.1073/pnas.131023798, Bibcode 2001PNAS...98.7777M, lire en ligne)
- (en) Derek Marsh, « Membrane water-penetration profiles from spin labels », European Biophysics Journal, vol. 31, no 7, , p. 559-562 (PMID 12602343, DOI 10.1007/s00249-002-0245-z, lire en ligne)
- (en) John F. Nagle et Stephanie Tristram-Nagle, « Structure of lipid bilayers », Biochimica et Biophysica Acta (BBA) - Reviews on Biomembranes, vol. 1469, no 3, , p. 159-195 (PMID 11063882, PMCID 2747654, DOI 10.1016/S0304-4157(00)00016-2, lire en ligne)
- (en) Félix M. Goñi, « Non-permanent proteins in membranes: when proteins come as visitors (Review) », Molecular Membrane Biology, vol. 19, no 4, , p. 237-245 (PMID 12512770, DOI 10.1080/0968768021000035078, lire en ligne)
- (en) David L. Scott, Zbyszek Otwinowski, Michael H. Gelb et Paul B. Sigler, « Crystal structure of bee-venom phospholipase A2 in a complex with a transition-state analogue », Science, vol. 250, no 4987, , p. 1563-1566 (PMID 2274788, DOI 10.1126/science.2274788, JSTOR 2878340, Bibcode 1990Sci...250.1563S)
- (en) Thomas J. McIntosh et Sidney A. Simon, « Roles of bilayer material properties in function and distribution of membrane proteins », Annual Review of Biophysics and Biomolecular Structure, vol. 35, , p. 177-198 (PMID 16689633, DOI 10.1146/annurev.biophys.35.040405.102022, lire en ligne)
- (en) Frank Hanakam, Günther Gerisch, Sandra Lotz, Thomas Alt et Anna Seelig, « Binding of Hisactophilin I and II to Lipid Membranes Is Controlled by a pH-Dependent Myristoyl−Histidine Switch », Biochemistry, vol. 35, no 34, , p. 11036-11044 (PMID 8780505, DOI 10.1021/bi960789j, lire en ligne)
- (en) John R. Silvius, « Lipidated peptides as tools for understanding the membrane interactions of lipid-modified proteins », Current Topics in Membranes, vol. 52, , p. 371-395 (DOI 10.1016/S1063-5823(02)52015-9, lire en ligne)
- (en) Nikola A. Baumann et Anant K. Menon, « Chapter 2 Lipid modifications of proteins », New Comprehensive Biochemistry, vol. 36, , p. 37-54 (DOI 10.1016/S0167-7306(02)36004-6, lire en ligne)
- (en) Jerónimo Bravo, Dimitrios Karathanassis, Christine M. Pacold, Michael E. Pacold, Chris D. Ellson, Karen E. Anderson, P. Jonathan G. Butler, Isabelle Lavenir, lga Perisic, Phillip T. Hawkins, Len Stephens et Roger L. Williams, « The crystal structure of the PX domain from p40phox bound to phosphatidylinositol 3-phosphate », Molecular Cell, vol. 8, no 4, , p. 829-839 (PMID 11684018, DOI 10.1016/S1097-2765(01)00372-0, lire en ligne)
- (en) Wonhwa Cho et Robert V. Stahelin, « Membrane-Protein Interactions in Cell Signaling and Membrane Trafficking », Annual Review of Biophysics and Biomolecular Structure, vol. 34, , p. 119-151 (PMID 15869386, DOI 10.1146/annurev.biophys.33.110502.133337, lire en ligne)
- (en) N. Ben-Tal, B. Honig, C. Miller et S. McLaughlin, « Electrostatic binding of proteins to membranes. Theoretical predictions and experimental results with charybdotoxin and phospholipid vesicles », Biophysical Journal, vol. 73, no 4, , p. 1717-1727 (PMID 9336168, PMCID 1181073, DOI 10.1016/S0006-3495(97)78203-1, Bibcode 1997BpJ....73.1717B, lire en ligne)
- (en) Mantripragada B. Sankaram et Derek Marsh, « Chapter 6 Protein-lipid interactions with peripheral membrane proteins », New Comprehensive Biochemistry, vol. 25, , p. 127-162 (DOI 10.1016/S0167-7306(08)60235-5, lire en ligne)
- (en) Nathan J. Malmberg et Joseph J. Falke, « Use of EPR Power Saturation to Analyze the Membrane-Docking Geometries of Peripheral Proteins: Applications to C2 Domains », Annual Review of Biophysics and Biomolecular Structure, vol. 34, , p. 71-90 (PMID 15869384, PMCID 3637887, DOI 10.1146/annurev.biophys.34.040204.144534, lire en ligne)
- (en) Andrew G. Spencer, Elizabeth Thuresson, James C. Otto, Inseok Song‡, Tim Smith, David L. DeWitt, R. Michael Garavito et William L. Smith, « The membrane binding domains of prostaglandin endoperoxide H synthases 1 and 2. Peptide mapping and mutational analysis. », Journal of Biological Chemistry, vol. 274, no 46, , p. 32936-32942 (PMID 10551860, DOI 10.1074/jbc.274.46.32936, lire en ligne)
- (en) Brian Lathrop, Martha Gadd, Rodney L. Biltonen et Gordon S. Rule, « Changes in Ca2+ Affinity upon Activation of Agkistrodon piscivorus piscivorus Phospholipase A2 », Biochemistry, vol. 40, no 11, , p. 3264-3272 (PMID 11258945, DOI 10.1021/bi001901n, lire en ligne)
- (en) Tatiana Kutateladze et Michael Overduin, « Structural Mechanism of Endosome Docking by the FYVE Domain », Science, vol. 291, no 5509, , p. 1793-1796 (PMID 11230696, DOI 10.1126/science.291.5509.1793, Bibcode 2001Sci...291.1793K, lire en ligne)
- (en) Suren A. Tatulian, Shan Qin, Abhay H. Pande et Xiaomei He, « Positioning Membrane Proteins by Novel Protein Engineering and Biophysical Approaches », Journal of Molecular Biology, vol. 351, no 5, , p. 939-947 (PMID 16055150, DOI 10.1016/j.jmb.2005.06.080, lire en ligne)
- (en) Kalina Hristova, William C. Wimley, Vinod K. Mishra, G. M. Anantharamiah, Jere P. Segrest et Stephen H. White, « An amphipathic α-helix at a membrane interface: a structural study using a novel X-ray diffraction method », Journal of Molecular Biology, vol. 290, no 1, , p. 99-117 (PMID 10388560, DOI 10.1006/jmbi.1999.2840, lire en ligne)
- (en) Diana Murray et Barry Honig, « Electrostatic Control of the Membrane Targeting of C2 Domains », Molecular Cell, vol. 9, no 1, , p. 145-154 (PMID 11804593, DOI 10.1016/S1097-2765(01)00426-9, lire en ligne)
- (en) R. G. Efremov, D. E. Nolde, A. G. Konshina, N. P. Syrtcev et A. S. Arseniev., « Peptides and Proteins in Membranes: What Can We Learn via Computer Simulations? », Current Medicinal Chemistry, vol. 11, no 18, , p. 2421-2442 (PMID 15379706, DOI 10.2174/0929867043364496, lire en ligne)
- (en) Andrei L. Lomize, Irina D. Pogozheva, Mikhail A. Lomize et Henry I. Mosberg, « Positioning of proteins in membranes: A computational approach », Protein Science, vol. 15, no 6, , p. 1318-1333 (PMID 16731967, PMCID 2242528, DOI 10.1110/ps.062126106, lire en ligne)
- (en) D. Papahadjopoulos, M. Moscarello, E. H. Eylar et T. Isac, « Effects of proteins on the thermotropic phase transitions of phospholipid membranes », Biochimica et Biophysica Acta (BBA) - Biomembranes, vol. 401, no 3, , p. 317-335 (PMID 52374, DOI 10.1016/0005-2736(75)90233-3, lire en ligne)
- (en) Joachim Seelig, « Thermodynamics of lipid–peptide interactions », Biochimica et Biophysica Acta (BBA) - Biomembranes, vol. 1666, nos 1-2, , p. 40-50 (PMID 15519307, DOI 10.1016/j.bbamem.2004.08.004, lire en ligne)
- (en) Malcolm J. M. Darkes, Sarah M. A. Davies et Jeremy P. Bradshaw, « Interaction of tachykinins with phospholipid membranes: A neutron diffraction study », Physica B: Condensed Matter, vol. 241-243, , p. 1144-1147 (DOI 10.1016/S0921-4526(97)00811-9, Bibcode 1997PhyB..241.1144D, lire en ligne)
- (en) Jeffrey F. Ellena, Jason Moulthrop, Jing Wu, Michelle Rauch, Sajith Jaysinghne, J. David Castle et David S. Cafiso, « Membrane Position of a Basic Aromatic Peptide that Sequesters Phosphatidylinositol 4,5 Bisphosphate Determined by Site-Directed Spin Labeling and High-Resolution NMR », Biophysical Journal, vol. 87, no 5, , p. 3221-3233 (PMID 15315949, PMCID 1304792, DOI 10.1529/biophysj.104.046748, Bibcode 2004BpJ....87.3221E, lire en ligne)
- (en) Isabelle Marcotte, Erick J. Dufourc, Marise Ouellet et Michèle Auger, « Interaction of the Neuropeptide Met-Enkephalin with Zwitterionic and Negatively Charged Bicelles as Viewed by 31P and 2H Solid-State NMR », Biophysical Journal, vol. 85, no 1, , p. 328-339 (PMID 12829487, PMCID 1303088, DOI 10.1016/S0006-3495(03)74477-4, Bibcode 2003BpJ....85..328M, lire en ligne)
- (en) Wenyi Zhang, Evan Crocker, Stuart McLaughlin et Steven O. Smith, « Binding of peptides with basic and aromatic residues to bilayer membranes: phenylalanine in the myristoylated alanine-rich C kinase substrate effector domain penetrates into the hydrophobic core of the bilayer », Journal of Biological Chemistry, vol. 278, no 24, , p. 21459-21466 (PMID 12670959, DOI 10.1074/jbc.M301652200, lire en ligne)