Protéine à ancrage lipidique
Les protéines à ancrage lipidique sont des protéines situées sur les deux faces de la membrane plasmique et qui sont liées par covalence à des lipides de cette membrane. La terminaison lipidique de ces protéines s'insère dans la structure de la bicouche lipidique et prend place parmi les chaînes aliphatiques des autres acides gras qui la constituent. Le lipide membranaire sert ainsi à ancrer la protéine à la membrane plasmique[1] - [2]. Il s'agit de protéolipides.
Les groupes lipidiques interviennent dans les interactions de la protéine et peuvent contribuer à la fonction biologique de la protéine auxquels ils sont liés[2]. Le lipide peut également servir de médiateur dans l'association de la membrane ou de déterminant pour certaines interactions protéine-protéine[3]. Par exemple, des groupes lipidiques peuvent jouer un rôle important en augmentant l'hydrophobie d'une molécule, ce qui permet à des domaines protéiques d'interagir avec des membranes biologiques[4]. La lipidation peut également isoler une enzyme de son substrat, ce qui a pour effet d'inactiver l'enzyme.
On connaît trois principaux types de protéines à ancrage lipidique : les protéines prénylées, les protéines acylées et les protéines à ancrage glycosylphosphatidylinositol[2] - [5] (GPI).
Protéines prénylées
Les protéines prénylées sont des protéines ayant des polymères hydrophobes d'isoprène — unités ramifiées à cinq atomes de carbone — liées par covalence à un ou plusieurs résidus de cystéine[2] - [3]. Plus précisément, ces groupes isoprénoïdes, généralement le farnésyle (trois unités isoprène) et le géranylgéranyle (quatre unités isoprène) sont liées à la protéine par des liaisons thioéther au niveau de résidus de cystéine près de l'extrémité C-terminale de la protéine[3] - [4]. La prénylation des protéines facilite l'interaction de ces dernières avec les membranes[1].

Les motifs de type « boîte CaaX » sont les sites de prénylation des protéines les plus courants, les groupes prényle se liant aux résidus de cystéine des boîtes CaaX[2] - [3]. Le motif CaaX est composé de quatre résidus d'acides aminés : C représente une cystéine ; a représente un acide aminé aliphatique quelconque ; et X représente un résidu dont la nature détermine le type de prénylation devant survenir sur la protéine : s'il s'agit d'un résidu d'alanine, de méthionine, de sérine ou de glutamine alors la protéine sera farnésylée par une farnésyltransférase (en), tandis que s'il s'agit d'un résidu de leucine alors la protéine sera géranylgéranylée par une géranylgéranyltransférase de type 1[3] - [4]. Ces deux enzymes sont semblables, chacune contenant deux sous-unités[6].
Fonctions biologiques
Les protéines prénylées sont particulièrement importantes dans la croissance, la différenciation et la morphogenèse des cellules d'eucaryotes[6]. La prénylation des protéines est également une modification post-traductionnelle réversible de certaines protéines membranaires. L'interaction dynamique entre protéines prénylées et membranes plasmiques est un processus important de la signalisation cellulaire souvent déréglé dans des maladies telles que les cancers[7]. Plus précisément, les protéines Ras sont prénylées par des farnésyltransférases (en) et, lorsqu'elles sont activées, elles activent également les gènes impliqués dans la croissance et la différenciation cellulaires, raison pour laquelle la suractivation des protéines Ras peut induire un cancer[8], la compréhension du rôle et du fonctionnement de ces protéines prénylées ayant été un élément important du développement de certains anticancéreux[9].
Outre les protéines Ras, les protéines Rab et Rho (en) sont prénylées, ainsi que les lamines[6].
Protéines acylées
Les protéines acylées sont des protéines ayant subi une modification post-traductionnelle conduisant à la liaison covalente d'un acide gras sur un ou plusieurs résidus d'acides aminés[10] - [11]. Les acides gras les plus couramment liés aux protéines sont l'acide myristique et l'acide palmitique, acides gras saturés en C14 et en C16 respectivement. Les protéines peuvent être liées à l'un ou l'autre ou à ces deux acides gras en même temps[10].
N-Myristoylation
La N-myristoylation, c'est-à-dire la liaison covalente d'acide myristique à une protéine, est une modification irréversible qui se produit généralement pendant la biosynthèse des protéines[10] - [12] au cours de laquelle l'acide myristique est lié au groupe α-aminé d'un résidu de glycine N-terminal à travers une liaison amide[2] - [11]. Cette réaction est facilitée par la glycylpeptide N-tétradécanoyltransférase 1 (en) et concerne des protéines dont la séquence commence généralement par Met–Gly avec un résidu de sérine ou de thréonine en position 5[10]. Les protéines N-myristoylées sont impliquées dans les cascades de transduction de signal, les interactions protéine-protéine et les mécanismes qui régulent le ciblage et la fonction des protéines[12]. C'est par exemple le cas de l'apoptose, ou mort cellulaire programmée . La myristoylation de la protéine BID (en) marque la protéine pour être déplacée vers la membrane mitochondriale externe et libérer le cytochrome c, lequel déclenche à son tour la mort cellulaire[13]. L'actine et la gelsoline interviennent également dans l'apoptose sous l'effet d'une myristoylation.
S-Palmitoylation
La S-palmitoylation, c'est-à-dire la liaison covalente d'acide palmitique à une protéine, est une modification post-traductionnelle réversible au cours de laquelle une molécule d'acide palmitique est liée à un résidu de cystéine spécifique au moyen d'une liaison thioester[2] - [10]. On parle également de S-acylation (en) lorsque d'autres chaînes d'acides gras sont également liées à des protéines palmitoylées. On n'a pas identifié de séquence consensus pour la palmitoylation des protéines[10]. On trouve les protéines palmitoylées principalement du côté cytosolique de la membrane plasmique, où elles jouent un rôle dans la signalisation cellulaire. Le groupe palmitoyle peut être clivé par les palmitoyl-protéine thioestérases (en). On pense que cette dépalmitoylation pourrait réguler l'interaction de la protéine avec la membrane et agir ainsi sur la signalisation transmembranaire[2].
La palmitoylation intervient de surcroît dans la régulation de la localisation, de la stabilité et de la circulation intracellulaire de la protéine[14]. La palmitoylation intervient par exemple dans le regroupement des protéines dans la synapse, où elle joue un rôle dans la signalisation cellulaire. Ainsi, lorsque la protéine PSD-95 (en) est palmitoylée, elle est confinée sur la membrane et lui permet de se lier à des canaux ioniques et de les accumuler au niveau de la membrane post-synaptique, de sorte que la palmitoylation peut intervenir dans la régulation de la libération des neurotransmetteurs[15].
La palmitoylation module l'affinité d'une protéine pour les radeaux lipidiques et facilite l'agglomération des protéines[16]. L'agglomération des protéines peut favoriser le rapprochement de deux molécules ou au contraire isoler une protéine de son substrat. Par exemple, la palmitoylation de la phospholipase D éloigne cette enzyme de la phosphatidylcholine, qui est son substrat ; lorsque le taux de cholestérol décroît ou que le taux de phosphatidylinositol-4,5-bisphosphate (PIP2) croît, la localisation intracellulaire des protéines palmitoylées est modifiée et l'enzyme se déplace en direction du PIP2, ce qui lui fait rencontrer de la phosphatidylcholine : c'est une activation par présentation du substrat (en)[17] - [18] - [19].
Protéines à ancrage GPI
Les protéins liées à un complexe glycosylphosphatidylinositol (GPI) le sont par une liaison amide sur le groupe carbonyle C-terminal[20]. Ce complexe GPI se compose de plusieurs constituants qui sont tous interconnectés : une phosphoéthanolamine, un tétraholoside linéaire constitué de trois résidus de mannose et d'un résidu de glucosamine, et un phosphatidylinositol[21]. Le groupe phosphatidylinositol est lié à la glucosamine non N-acétylée du tétraholoside par une liaison osidique. Il se forme une liaison phosphodiester entre la phosphoéthanolamine et le mannose de l'extrémité non réductrice du tétraholoside. La phosphoéthanolamine est alors liée au carboxyle C-terminal de la protéine par une liaison amide[2].
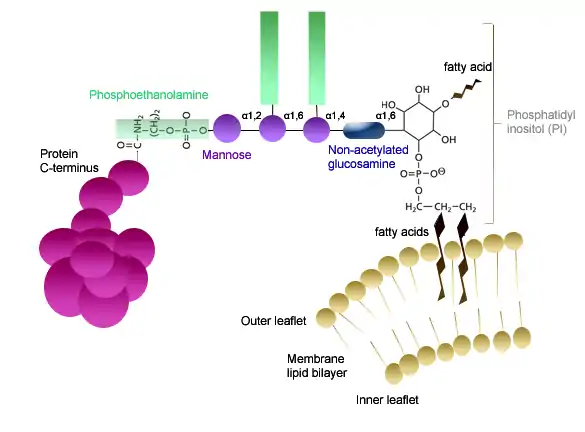 (en) Structure schématique d'une ancre GPI, indiquant l'enchaînement de la protéine, de la phosphoéthanolamine, des quatre résidus osidiques et le groupe phosphatidylinositol, lui-même lié aux acides gras insérés dans le feuillet externe de la bicouche lipidique de la membrane plasmique.
(en) Structure schématique d'une ancre GPI, indiquant l'enchaînement de la protéine, de la phosphoéthanolamine, des quatre résidus osidiques et le groupe phosphatidylinositol, lui-même lié aux acides gras insérés dans le feuillet externe de la bicouche lipidique de la membrane plasmique.
La liaison de la protéine au GPI est réalisée par un complexe GPI-transamidase[21]. Les chaînes d'acide gras du phosphatidylinositol s'insèrent dans le feuillet externe de la bicouche lipidique de la membrane plasmique[2], ce qui assure l'ancrage de la protéine à cette dernière[20].
Fonctions biologiques
Les résidus osidiques du tétraholoside et les résidus d'acides gras du groupe phosphatidylinositol varient en fonction de la protéine[2]. C'est cette diversité structurelle qui offre aux protéines à ancrage GPI une grande variété de fonctions biologiques, notamment en agissant comme hydrolases, protéines d'adhésion cellulaire, récepteurs, inhibiteurs de protéases et protéines régulatrices du complément (en)[22].
Les protéines à ancrage GPI jouent également un rôle important dans l'embryogenèse, la neurogenèse, le développement, le système immunitaire et la fécondation[20]. Plus précisément, l'ancrage GPI de la protéine Junon (en) de l'ovule joue un rôle déterminant dans la fusion du spermatozoïde avec l'ovule. La libération de cette protéine de son ancrage GPI à la membrane plasmique de l'ovule bloque la fusion de ce dernier avec des spermatozoïdes, ce qui pourrait contribuer au blocage de la polyspermie[23].
Notes et références
- (en) Gerald Karp, Cell and Molecular Biology: Concepts and Experiments, 2009, John Wiley and Sons, p. 128. (ISBN 978-0-470-48337-4)
- (en) D. Voet, J. G. Voet et C. W. Pratt, Fundamentals of Biochemistry: Life at the Molecular Level, 4e éd., 2013, John Wiley & Sons, p. 263. (ISBN 978-0470-54784-7).
- (en) Patrick J. Casey et Miguel C. Seabra, « Protein Prenyltransferases », Journal of Biological Chemistry, vol. 271, no 10, , p. 5289-5292 (PMID 8621375, DOI 10.1074/jbc.271.10.5289, lire en ligne)
- (en) Giuseppe Novelli et Maria Rosaria D'Apice, « Protein farnesylation and disease », Journal of Inherited Metabolic Diseases, vol. 35, no 5, , p. 917-926 (PMID 22307208, DOI 10.1007/s10545-011-9445-y, lire en ligne)
- (en) Michael A. J. Ferguson, « Lipid anchors on membrane proteins », Current Opinion in Structural Biology, vol. 1, no 4, , p. 522-529 (DOI 10.1016/S0959-440X(05)80072-7, lire en ligne)
- (en) Kimberly T. Lane et Lorena S. Beese, « Thematic review series: Lipid Posttranslational Modifications. Structural biology of protein farnesyltransferase and geranylgeranyltransferase type I », Journal of Lipid Research, vol. 47, no 4, , p. 681-699 (PMID 16477080, DOI 10.1194/jlr.R600002-JLR200, lire en ligne)
- (en) Viktor Stein, Marta H. Kubala, Jason Steen, Sean M. Grimmond et Kirill Alexandrov, « Towards the Systematic Mapping and Engineering of the Protein Prenylation Machinery in Saccharomyces cerevisiae », PLoS One, vol. 10, no 3, , article no e0120716 (PMID 25768003, PMCID 4358939, DOI 10.1371/journal.pone.0120716, Bibcode 2015PLoSO..1020716S, lire en ligne)
- (en) David S. Goodsell, « The Molecular Perspective: The ras Oncogene », The Oncologist, vol. 4, no 3, , p. 263-264 (PMID 10394594, lire en ligne)
- (en) Christoph W. M. Reuter, Michael A. Morgan et Lothar Bergmann, « Targeting the Ras signaling pathway: a rational, mechanism-based treatment for hematologic malignancies? », Blood, vol. 96, no 5, , p. 1655-1669 (PMID 10961860, DOI 10.1182/blood.V96.5.1655, lire en ligne)
- (en) Marilyn D. Resh, « Trafficking and signaling by fatty-acylated and prenylated proteins », Nature Chemical Biology, vol. 2, no 11, , p. 584-590 (PMID 17051234, DOI 10.1038/nchembio834, lire en ligne)
- (en) John P. Wilson, Anuradha S. Raghavan, Yu-Ying Yang, Guillaume Charron et Howard C. Hang, « Proteomic Analysis of Fatty-acylated Proteins in Mammalian Cells with Chemical Reporters Reveals S-Acylation of Histone H3 Variants », Molecular & Cellular Proteomics, vol. 10, no 3, , article no M110.001198 (PMID 21076176, PMCID 3047146, DOI 10.1074/mcp.M110.001198, lire en ligne)
- (en) Thalia A. Farazi, Gabriel Waksman et Jeffrey I. Gordon, « The Biology and Enzymology of Protein N-Myristoylation », Journal of Biological Chemistry, vol. 276, no 43, , p. 39501-39504 (PMID 11527981, DOI 10.1074/jbc.R100042200, lire en ligne)
- (en) Dale D. O. Martin, Erwan Beauchamp et Luc G. Berthiaume, « Post-translational myristoylation: Fat matters in cellular life and death », Biochimie, vol. 93, no 1, , p. 18-31 (PMID 21056615, DOI 10.1016/j.biochi.2010.10.018, lire en ligne)
- (en) Clara Aicart-Ramos, Ruth Ana et Valero Ignacio Rodriguez-Crespo, « Protein palmitoylation and subcellular trafficking », Biochimica et Biophysica Acta (BBA) - Biomembranes, vol. 1808, no 12, , p. 2981-2994 (PMID 21819967, DOI 10.1016/j.bbamem.2011.07.009, lire en ligne)
- (en) Rochelle M. Hines et Alaa El-Husseini, « Mechanisms That Regulate Neuronal Protein Clustering at the Synapse », Alexander Dityatev et Alaa El-Husseini, Molecular Mechanisms of Synaptogenesis, p. 72-75, Springer, Boston, États-Unis, 2006. (ISBN 978-0-387-32560-6)
- (en) Ilya Levental, Daniel Lingwood, Michal Grzybek, Ünal Coskun et Kai Simons, « Palmitoylation regulates raft affinity for the majority of integral raft proteins », Proceedings of the National Academy of Sciences of the United States of America, vol. 107, no 51, , p. 22050-22054 (PMID 21131568, PMCID 3009825, DOI 10.1073/pnas.1016184107, Bibcode 2010PNAS..10722050L, lire en ligne)
- (en) E. Nicholas Petersen, Hae-Won Chung, Arman Nayebosadri et Scott B. Hansen, « Kinetic disruption of lipid rafts is a mechanosensor for phospholipase D », Nature Communications, vol. 7, , article no 13873 (PMID 27976674, PMCID 5171650, DOI 10.1038/ncomms13873, Bibcode 2016NatCo...713873P, lire en ligne)
- (en) Carol V. Robinson, Tibor Rohacs et Scott B. Hansen, « Tools for Understanding Nanoscale Lipid Regulation of Ion Channels », Trends in Biochemical Sciences, vol. 44, no 9, , p. 795-806 (PMID 31060927, PMCID 6729126, DOI 10.1016/j.tibs.2019.04.001, lire en ligne)
- (en) E. Nicholas Petersen, Mahmud Arif Pavel, Hao Wang et Scott B. Hansen, « Disruption of palmitate-mediated localization; a shared pathway of force and anesthetic activation of TREK-1 channels », Biochimica et Biophysica Acta (BBA) - Biomembranes, vol. 1862, no 1, , article no 183091 (PMID 31672538, PMCID 6907892, DOI 10.1016/j.bbamem.2019.183091, lire en ligne)
- (en) Taroh Kinoshita et Morihisa Fujita, « Biosynthesis of GPI-anchored proteins: special emphasis on GPI lipid remodeling », Journal of Lipid Research, vol. 57, no 1, , p. 6-24 (PMID 26563290, PMCID 4689344, DOI 10.1194/jlr.R063313, lire en ligne)
- (en) Hiroh Ikezawa, « Glycosylphosphatidylinositol (GPI)-Anchored Proteins », Biological and Pharmaceutical Bulletin, vol. 25, no 4, , p. 409-417 (PMID 11995915, DOI 10.1248/bpb.25.409, lire en ligne)
- (en) Taroh KINOSHITA, « Biosynthesis and deficiencies of glycosylphosphatidylinositol », Proceedings of the Japan Academy, Series B, vol. 90, no 4, , p. 130-143 (PMID 24727937, PMCID 4055706, DOI 10.2183/pjab.90.130, Bibcode 2014PJAB...90..130K, lire en ligne)
- (en) Scott A. Coonrod, Soren Naaby-Hansen, Jagathpala Shetty, Hiroaki Shibahara, Michellee Chen, Judith M. White et John C. Herr, « Treatment of Mouse Oocytes with PI-PLC Releases 70-kDa (pI 5) and 35- to 45-kDa (pI 5.5) Protein Clusters from the Egg Surface and Inhibits Sperm–Oolemma Binding and Fusion », Developmental Biology, vol. 207, no 2, , p. 334-349 (PMID 10068467, DOI 10.1006/dbio.1998.9161, lire en ligne)