Fossa (animal)
Cryptoprocta ferox

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Classe | Mammalia |
| Ordre | Carnivora |
| Sous-ordre | Feliformia |
| Famille | Eupleridae |
| Sous-famille | Euplerinae |
| Genre | Cryptoprocta |
- Cryptoprocta typicus A. Smith, 1834
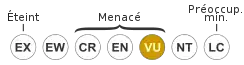
VU A2cd : Vulnérable
[1]
Statut CITES
Le fossa (Cryptoprocta ferox), aussi appelé cryptoprocte féroce et unique représentant actuel du genre Cryptoprocta, est une espèce de mammifères féliformes de la famille des Eupleridae, endémique de Madagascar. Il est le plus gros mammifère carnivore de l'île et est parfois comparé à un petit puma. Les adultes mesurent 70 à 80 cm de long du museau à la base de la queue, et pèsent de 5,5 à 8,6 kg, les mâles étant plus lourds que les femelles. Ses griffes semi-rétractiles permettent au fossa de grimper et de descendre des arbres la tête en avant, et l'animal peut aussi sauter d'arbre en arbre. Le fossa est un représentant très particulier dans sa famille, ses organes génitaux partageant certains caractères avec les félins et les hyènes.
Le fossa a une aire de répartition répandue sur Madagascar, mais les densités de peuplement sont généralement faibles. Il vit uniquement en habitat forestier, et chasse activement la nuit comme le jour. Plus de la moitié de son régime alimentaire se compose de lémuriens, les primates également endémiques de l'île ; les autres proies connues sont représentées par des tenrecs, des rongeurs, des lézards, des oiseaux et divers autres animaux. L'accouplement a habituellement lieu dans des arbres, sur des branches horizontales, et peut durer plusieurs heures. La portée compte un à six petits, qui naissent aveugles et sans dents. Les jeunes sont sevrés au bout de quatre mois et demi, et sont indépendants au bout d'un an. Ils atteignent la maturité sexuelle à l'âge de deux ans, et la longévité mesurée en captivité est de vingt ans.
Sa classification a été sujette à débats, l'animal partageant des traits avec les félins, d'autres suggérant une relation étroite avec les viverridés. Sa position phylogénétique, avec celles des autres carnivores malgaches, a influencé les hypothèses sur le nombre de colonisations de l'île par les mammifères carnivores. Les études génétiques ayant montré que le fossa et tous les autres carnivores de Madagascar étant plus proches entre eux, formant le clade qu'est la famille des Eupleridae, on sait aujourd'hui que les carnivores ont colonisé l'île en une seule arrivée, remontant à 18 ou 20 millions d'années. Le fossa est considéré comme « vulnérable » par l'Union internationale pour la conservation de la nature, ses effectifs sont jugés en déclin et principalement menacés par la destruction de l'habitat. Il est généralement craint par le peuple malgache et est souvent protégé par un tabou (fady en malgache) qui le préserve de la chasse.
Description

Aspect général
Le fossa ressemble à une forme diminuée d'un grand félin, comme un puma[2], mais avec un corps svelte et des membres musclés[3], et une queue presque aussi longue que le reste du corps[2]. Sa tête ressemble à celle d'une mangouste, relativement plus longue que celle d'un chat et avec un museau large et court, de grandes oreilles arrondies[3] - [2]. Il a les yeux bruns relativement écartés aux pupilles fendues. Comme de nombreux carnivores qui chassent la nuit, il possède un tapetum lucidum augmentant son acuité visuelle, et qui reflète une lumière orange[3]. Le Fossa mesure 70 à 80 cm de long du museau à la base de la queue et celle-ci est longue de quelque 65 à 70 centimètres. Il y a un certain dimorphisme sexuel, les mâles adultes étant plus grands que les femelles, avec des fourchettes de poids respectives de 6,2-8,6 kg et de 5,5-6,8 kg[2]. Les individus plus petits se trouvent généralement dans le Nord et l'Est de Madagascar, tandis que les plus grands se répartissent vers le sud et dans l'Ouest[4]. De grands individus pesant exceptionnellement jusqu'à 20 kg ont été signalés, mais il existe un certain doute quant à la fiabilité des mesures[2]. Le fossa a de bonnes facultés olfactives, auditives et visuelles ; il est robuste et les maladies sont rares en captivité[5].
Les mâles et les femelles ont le poil court et droit, formant une fourrure unie et relativement dense. Les deux sexes ont le dos généralement brun roussâtre et le ventre d'un crème sale. En rut, ils peuvent avoir une coloration orange sur leur abdomen à cause d'une substance rougeâtre sécrétée par des glandes exocrines pulmonaires, mais cela n'a pas été observé par tous les chercheurs. La queue est généralement d'une coloration plus claire que les flancs. Les juvéniles sont gris ou presque blancs[4] - [2].
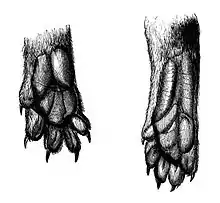
Plusieurs des caractéristiques physiques de l'animal sont des adaptations au milieu arboricole[3]. Sa queue l'aide à s'équilibrer et ses griffes semi-rétractiles lui permettent de grimper aux arbres à la recherche d'une proie[2]. Ses pattes sont semi-plantigrades[4], l'animal alternant entre une démarche arboricole aux allures plantigrades et un déplacement plutôt digitigrade au sol[6]. Le dessous de ses pattes est presque nu et couvert de solides coussinets[4]. Le fossa est pourvu de chevilles très flexibles qui lui permettent de s'agripper facilement aux troncs pour y grimper ou en descendre, tête la première, ou encore de sauter d'arbre en arbre. En captivité, on a observé les juvéniles se balancer tête en bas, accrochés à des cordes à nœuds par leurs pattes arrière[3].
Le fossa dispose de plusieurs glandes odorantes, moins développées chez la femelle que chez le mâle. Comme les mangoustes, il possède une glande périanale à l'intérieur d'un sac anal entourant l'anus comme une poche. Cette dernière débouche à l'extérieur par une fente horizontale au-dessous de la queue. D'autres glandes sont situées près du pénis ou du vagin, les glandes péniennes émettant une forte odeur. Comme les mangoustes, il n'a pas de glandes préscrotales[4].
Organes génitaux
Les organes génitaux externes sont l'une des caractéristiques physiques les plus intéressantes de cette espèce. Le mâle a le pénis (et son baculum) exceptionnellement long, parvenant en érection entre ses pattes avant. Le gland se prolonge jusque vers le milieu du pénis et est épineux, sauf en son bout ; à titre de comparaison, le gland des félidés est court et épineux, tandis que celui des viverridés est lisse et long[4]. La femelle fossa présente une masculinisation transitoire, commençant à un ou deux ans d'âge, et consistant dans le développement d'un clitoris imposant et épineux qui prenant la forme d'un pseudo-pénis. Le clitoris est soutenu par un os clitoridien, qui diminue à mesure que l'animal grandit[2] - [6]. Les femelles n'ont pas un pseudo-scrotum[2], mais sécrètent une substance orange qui colore leurs parties inférieures, très semblablement aux sécrétions des mâles[7]. Les niveaux d'hormones (testostérone, androstènedione, dihydrotestostérone) ne semblent pas jouer un rôle dans cette masculinisation transitoire, ces niveaux étant identiques chez les juvéniles masculinisés et les adultes non masculinisés. La masculinisation transitoire pourrait réduire le harcèlement sexuel des jeunes femelles par les mâles adultes, ou réduire l'agressivité des femelles territoriales[2]. Si les femelles d'autres espèces de mammifères (comme la hyène tachetée) ont également un pseudo-pénis, chez aucune de celles-ci ce clitoris hypertrophié ne décroît en volume quand l'animal grandit[8] - [7].
Comparaison avec les carnivores proches

Dans l'ensemble, le fossa a des caractéristiques en commun avec trois familles de carnivores différentes, et d'éminents scientifiques l'ont placé, ainsi que d'autres membres des Eupleridae, dans les Herpestidae, les Viverridae et les Felidae. Les caractères félins sont principalement ceux associés au système digestif[4], comme la forme des dents et les parties faciales du crâne, la langue et le tube digestif, typiques de son régime alimentaire exclusivement carnivore[3]. Le reste du crâne ressemble plus à celui du genre Viverra, tandis que la structure générale du corps est très semblable à celle des différents membres de la famille des Herpestidae.
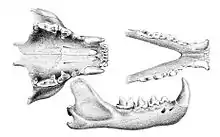
La dentition permanente est 3.1.3-4.13.1.3-4.1 (trois incisives, une canine, trois ou quatre prémolaires et une molaire de chaque côté sur chaque demi-mâchoire), la formule déciduale étant similaire mais dépourvue de la quatrième prémolaire et de la molaire. Le fossa a un grand rhinarium proéminent similaire à celui des viverridés, mais a des oreilles rondes plus grandes, presque aussi grandes que celles des félins de taille similaire. Les vibrisses de sa face sont longues, les plus longues l'étant plus que sa tête. Comme certains genres de mangoustes, en particulier Galidia (qui est aussi de la famille des Eupleridae) et Herpestes (des Herpestidae), il est muni de vibrisses carpiennes. Ses griffes sont rétractiles mais ne sont pas cachées dans une gaine de peau contrairement à celles des félidés. Le fossa a trois paires de mamelons (une inguinale, une ventrale et une pectorale)[2].
Écologie et comportement
Alimentation
Le fossa est l'un des huit carnivores endémiques de Madagascar. Opportuniste, il chasse des proies de taille petite à moyenne. Il est le plus grand mammifère terrestre actuel endémique de l'île et le seul prédateur capable de s'en prendre aux adultes de toutes les espèces de lémuriens existantes[9] - [10], la plus grande pouvant peser jusqu'à 90 % du poids d'un fossa de taille moyenne[3] - [10]. Bien qu'il soit le principal prédateur des lémuriens[10] - [11], les relevés de ses habitudes alimentaires révèlent une grande variété de sélectivité de proies entre les individus et une spécialisation en fonction de l'habitat et de la saison ; le régime alimentaire ne varie pas selon le sexe. Par exemple, alors que le fossa est spécialisé dans la chasse de lémuriens dans le parc national de Ranomafana, son régime alimentaire est plus variable dans les autres habitats des forêts humides[12].

Le régime alimentaire du fossa sauvage a été étudié par l'analyse des excréments caractéristiques de l'espèce, qui ressemblent à des cylindres gris aux extrémités torsadées, et mesurent de 10 à 14 cm de long pour 1,5 à 2,5 cm d'épaisseur[13]. Les fèces collectées et analysées à Andohahela et Andringitra renferment des restes de lémuriens et de rongeurs. Les populations de l'Est du massif d'Andringitra ont la plus grande variété enregistrée de proies, des vertébrés comme des invertébrés. Les vertébrés consommés comprennent des reptiles, une grande variété d'oiseaux (du sous-bois ou terrestres) et des mammifères, avec des insectivores, des rongeurs, et des lémuriens. Les invertébrés consommés par le fossa dans les hautes montagnes de l'Andringitra comptent des insectes et des crabes[14] - [15]. L'étude de ces excréments montre que les vertébrés représenteraient 94 % du régime alimentaire du fossa, avec les lémuriens comptant pour plus de la moitié, suivis des tenrecs (9 %), des lézards (9 %) et des oiseaux (2 %). Les graines, qui représenteraient 5 % de l'alimentation, proviennent peut-être des estomacs des lémuriens ingérés, ou de fruits consommés par le fossa pour leur eau, ces graines étant plus fréquentes dans l'alimentation pendant la saison sèche. La taille moyenne des proies varie géographiquement, avec seulement 40 grammes dans les hautes montagnes de l'Andringitra, mais 480 grammes dans les forêts humides et plus de 1 000 grammes dans les forêts sèches décidues[2]. Selon une étude du régime alimentaire du fossa menée dans la forêt sèche à feuilles caduques de l'ouest de Madagascar, plus de 90 % des proies étaient des vertébrés, et plus de 50 % étaient des lémuriens. La base du régime est constituée d'environ six espèces de lémuriens et de deux ou trois espèces de tenrecs-hérissons, de serpents et de petits mammifères[13]. En règle générale, le fossa cherche à chasser les plus gros lémuriens et rongeurs, et non les plus petits[16].

Le fossa capture ses proies au sol ou dans les arbres. S'il chasse seul en dehors de la période de reproduction, les partenaires d'un couple, ou plus tard les mères et leurs jeunes peuvent chasser ensemble lors de la saison reproductrice. Un fossa du groupe escalade un arbre et déniche des lémuriens qu'il poursuit d'arbre en arbre, les forçant à descendre vers le sol où les autres fossas peuvent facilement les capturer[2]. Le fossa éviscère les plus gros lémuriens qu'il capture ce qui, avec ses excréments caractéristiques, permet d'identifier ses proies[10]. Les observations sur le long terme des modes de prédation du fossa sur les propithèques de la forêt tropicale suggèrent que l'animal chasse dans une parcelle de son territoire jusqu'à ce que la densité de proies y diminue et qu'il lui faille se déplacer[17]. Le fossa peut parfois s'attaquer aux animaux domestiques, comme les chèvres et les petits veaux, et surtout aux poules. En captivité, le fossa consomme entre 800 et 1 000 grammes de viande par jour, des amphibiens, oiseaux, insectes, reptiles, et petits ou moyens mammifères[4].
Cette grande variété de proies chassées dans des habitats différents de la forêt tropicale est similaire à la composition variée du régime alimentaire constatée dans les forêts sèches de l'ouest de Madagascar[14] - [15]. Le fossa étant le plus grand prédateur endémique à Madagascar, sa flexibilité alimentaire combinée à une activité flexible[9] lui a permis d'exploiter une grande variété de niches disponibles dans toute l'île[14] - [15], et fait potentiellement de lui une espèce clé pour les écosystèmes malgaches[18].
Reproduction
La plupart des détails connus de la reproduction à l'état sauvage proviennent des populations des forêts sèches à feuilles caduques dans l'Ouest de l'île. D'autres recherches de terrain sont nécessaires pour savoir si ces connaissances sont généralisables aux populations orientales[2]. L'accouplement a généralement lieu en septembre et octobre[4], même s'il peut parfois être réalisé jusqu'en décembre[2], et peut être très ostensible[3]. En captivité dans l'hémisphère Nord, les fossas copulent au printemps, de mars à juillet[5]. Dans la nature, le rapport a habituellement lieu dans les arbres, sur des branches horizontales situées à une vingtaine de mètres du sol. Souvent, le même arbre est utilisé année après année, avec une précision remarquable quant à la date du début de la saison. Les arbres utilisés sont souvent situés à proximité d'une source d'eau, et ont de grosses branches assez fortes et assez larges, d'une vingtaine de centimètres de diamètre, pour supporter le couple reproducteur. On a également rapporté certains cas d'accouplement au sol[2].

Il peut y avoir jusqu'à huit mâles sur un même site d'accouplement, restant à proximité immédiate de la femelle réceptive. La femelle semble choisir ses partenaires, et les mâles se disputent son attention par une quantité importante de vocalisations et d'interactions hostiles. La femelle peut choisir de s'accoupler avec plusieurs partenaires, et son choix ne semble pas être lié à l'apparence physique des mâles[2]. Elle produit une série de miaulements pour stimuler le mâle. Celui-ci la monte par derrière, s'appuyant de manière légèrement excentrée[2], une posture exigeant un équilibre délicat ; si la femelle devait être maintenue, le mâle aurait une difficulté significative pour accomplir l'acte. Il pose ses pattes sur les épaules de sa partenaire[3] ou la saisit autour de la taille, et lèche souvent son cou[2]. L'accouplement peut durer près de trois heures. Cet accouplement d'une longueur inhabituelle est dû à la nature physique du pénis en érection, qui a des épines pointant vers l'arrière sur presque toute sa longueur[7]. Celles-ci pourraient également être responsables du lien copulatoire qui attache l'un à l'autre les deux partenaires, et qui leur rend difficile de se séparer même si l'accouplement vient à être interrompu[2] - [7]. La femelle peut copuler plusieurs fois avec un même mâle, avec un temps total d'accouplement pouvant atteindre quatorze heures, et le mâle peut rester avec la femelle jusqu'à une heure après l'accouplement. Une seule femelle peut occuper l'arbre, pour s'accoupler avec plusieurs mâles, jusqu'à une semaine. Il se peut aussi que d'autres femelles prennent sa place pour s'accoupler avec les mêmes mâles ou d'autres[2]. Cette stratégie d'accouplement, où les femelles monopolisent un site et maximisent le nombre disponible de partenaires, semble être unique parmi les carnivores. Une recherche publiée en 2009 suggère que ce système aide le fossa à surmonter des facteurs qui seraient normalement bloquants pour trouver des partenaires, comme les faibles densités de population et la non-utilisation de tanières[19].
La portée compte un à six petits, généralement deux à quatre[6] - [4]. La mise bas a lieu dans un endroit caché, comme une tanière souterraine, une termitière, une anfractuosité de rochers ou dans le creux d'un grand arbre[2] (en particulier ceux du genre Commiphora[3]). Contrairement à ce que rapportaient d'anciennes recherches, les portées sont de sexes mixtes[4] - [2]. Les jeunes naissent en décembre ou janvier, après une période de gestation de 90 jours[4] ou de six à sept semaines pour les accouplements les plus tardifs[2]. En juin 2018, le zoo de Paris a vu naître trois petits fossas, après une gestation de moins de 50 jours selon le zoo[20]. Les nouveau-nés sont aveugles et dépourvus de dents et ne pèsent pas plus de 100 grammes[4] - [2]. Ils sont recouverts d'une fine fourrure gris-brun[5] ou presque blanche[2]. Ils ouvrent les yeux au bout de deux semaines environ[4] - [5] ; ils deviennent plus actifs et leur fourrure tend vers le gris perle[2]. Les petits ne consomment pas de nourriture solide avant leurs trois mois, ne quittent la tanière que vers l'âge de 4,5 mois et sont sevrés peu de temps après[4] - [2]. Au bout de leur première année, les juvéniles sont indépendants de leur mère[2]. La dentition définitive apparait vers l'âge de 18 à 20 mois[4] - [2]. Les jeunes atteignent leur maturité physique aux environs de leurs deux ans[7] et leur maturité sexuelle au bout d'une ou deux années supplémentaires[4] - [2], et ils peuvent rester avec leur mère jusqu'à ce qu'ils soient complètement mûrs. La durée de vie en captivité est proche de la vingtaine d'années ou plus, probablement en raison de la lenteur du développement des jeunes[6].
Comportement

Le fossa est actif tant en journée que la nuit et est considéré comme cathéméral ; les pics d'activité se situent tôt le matin, tard dans l'après-midi et tard dans la nuit. L'animal ne réutilise généralement pas les sites de couchage, mais les mères accompagnées de leurs jeunes retournent à la même tanière[2]. Le domaine vital d'un mâle dans la forêt de Kirindy couvre jusqu'à 26 km2, contre 13 km2 pour une femelle. Ces territoires se chevauchent, d'environ 30 % selon des données provenant des forêts orientales, mais les femelles ont généralement des répartitions séparées. Ces aires croissent à la saison sèche, peut-être à cause d'une nourriture moins abondante et d'une disponibilité en eau plus faible. Des fossas suivis par colliers émetteurs parcouraient entre 2 et 5 km quotidiennement[9] mais le cas d'un individu ayant fait 7 km en 16 heures a également été rapporté[2]. La densité de population du fossa semble faible : dans la forêt de Kirindy, où il est dit commun, on estime qu'il y a un animal par 4 km2 en 1998[3]. Une autre étude dans la même forêt menée entre 1994 et 1996 et fondée sur une technique de capture-marquage-recapture et de reprise dénombrait un individu par 3,8 km2 et un adulte pour chaque 5,6 km2[21].
À part les mères accompagnées de leurs jeunes et des observations occasionnelles de couples de mâles, les individus vivent généralement seuls, et l'espèce est considérée comme solitaire[4] - [3] - [21]. Une publication de 2009 reporte cependant une observation détaillée d'une chasse coopérative de 45 minutes entre trois mâles, qui capturent et se partagent un sifaka de Verreaux (Propithecus verreauxi) de trois kilogrammes. Ce comportement est peut-être un vestige d'une chasse coopérative nécessaire pour la chasse des lémuriens géants récemment éteints[22].
Les fossas communiquent en utilisant des sons, des odeurs et des signaux visuels. Les vocalisations comprennent des ronronnements, un cri de menace[4] et un cri de peur composé d'inhalations grasses, fortes et répétées et de halètements[3]. Un long cri aigu pourrait servir à attirer d'autres individus. Le mâle émet un soupir lorsqu'il a trouvé une femelle, et celle-ci miaule pendant l'accouplement[4] - [3]. Les fossas déposent leurs odeurs tout au long de l'année, sur les rochers, les arbres et le sol, en utilisant des glandes de la région anale et de la poitrine[4] - [3] - [2]. Ils communiquent également en utilisant des expressions faciales et corporelles mais la signification de ces signaux est incertaine. L'animal n'est agressif que lors de l'accouplement, et les mâles notamment se livrent à des luttes voyantes. Après un court combat, le perdant fuit, poursuivi sur une courte distance par le gagnant[4]. En captivité, les fossas ne sont généralement pas agressifs et se laissent parfois caresser par les gardiens de zoo, mais les mâles adultes, en particulier, peuvent essayer de mordre[5].
Répartition
_(9578487052).jpg.webp)
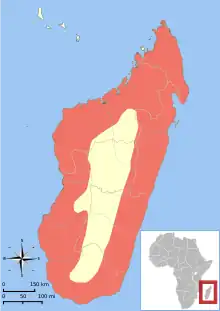
Le fossa a la répartition géographique la plus étendue des carnivores malgaches ; il vit généralement en petit nombre dans toutes les étendues de forêts restantes sur l'île, préférant la forêt vierge et tranquille. Il est également rencontré dans certaines forêts dégradées, mais en moins grand nombre. Bien que le fossa se trouve dans tous les habitats forestiers connus sur Madagascar, y compris les forêts sèches décidues occidentales, les forêts humides de l'Est, et les forêts épineuses du Sud[18], on l'observe plus souvent dans des lieux humides que dans les forêts sèches, peut-être parce que la canopée réduite dans les forêts sèches fournit moins d'ombre, et aussi parce que l'animal semble se déplacer plus facilement dans les forêts humides[3]. Il est absent des zones à l'habitat fortement perturbé et, comme la plupart de la faune malgache, du haut plateau central du pays[18].
La répartition du fossa suit différents gradients altitudinaux dans les parties intactes des zones protégées de Madagascar. Dans la Réserve Naturelle Intégrale d'Andringitra, des signes de présence du fossa ont été signalés dans quatre sites différents, allant de 810 à 1 625 mètres d'altitude[14]. Une occurrence record à 2 000 mètres d'altitude est également connue[24]. La forte présence de l'animal sur le massif d'Andringitra a été confirmée en 1996[14]. Dans le parc national d'Andohahela, le fossa vit entre 440 et 1 875 mètres d'altitude[15]. Sa présence dans ces endroits indique sa capacité à s'adapter à différentes altitudes, lui offrant une distribution couvrant tous les types de forêts sur Madagascar[18].
Dénominations et systématique
Étymologie
Le nom de genre, Cryptoprocta, est formé du grec ancien κρυπτός, (kruptos), « caché » et πρωκτός (proktόs), « anus », l'animal étant muni d'une poche anale[4]. La dénomination spécifique, ferox, vient du latin et signifie « fougueux, féroce », dérivé de ferus (« sauvage »)[25]. Son nom vulgaire de « fossa » vient de l'appellation locale malgache, « fosa »[4] - [26] prononcé /fusa̤/. Le mot est proche de « posa », signifiant « chat » en Iban, une autre langue austronésienne, de Bornéo, et les deux termes pourraient découler de langues véhiculaires commerciales des années 1600. Il est également possible que le mot dérive du malais pusa, se référant à la belette malaise (Mustela nudipes), et qui aurait donné posa pour les chats sur Bornéo, alors qu'à Madagascar le mot aurait pu devenir fosa pour désigner le fossa. Une dernière théorie, défendue par les guides du parc zoologique et botanique du Tsimbazaza d'Antananarivo, indique que le terme "fossa" est en fait lié à la découverte de l'animal qui ressemblait à un chat. Lorsque cet animal a ensuite été identifié comme étant une mangouste et non un félin, la population malgache le surnomma "faux chat" (à prononcer "fossa" puisque le "ch" se prononce "se" en malgache)[26].
Taxinomie et phylogénie

Le fossa a été initialement décrit par Edward Turner Bennett sur la base de matériel envoyé de Madagascar par Charles Telfair en 1833[27]. Si son nom commun est le même que le nom de genre de la civette malgache (Fossa fossana), ce sont deux espèces proches mais distinctes. En raison de traits physiques communs avec les civettes, les mangoustes, et les félins, sa classification a été controversée. Bennett considère d'abord le fossa comme un type de civette de la famille des Viverridae, une classification qui persiste longtemps parmi les taxinomistes. Sa boîte crânienne compacte, ses grandes orbites, ses griffes partiellement rétractiles et sa dentition spécialisée de carnivore ont également conduit certains taxinomistes à placer l'animal parmi les félidés[3]. En 1939, William King Gregory et Milo Hellman créent une sous-famille propre au fossa au sein des félidés, celle des Cryptoproctinae. George Gaylord Simpson replace l'animal chez les Viverridae en 1945, toujours dans sa propre sous-famille, mais en reconnaissant qu'il partage de nombreuses caractéristiques de félins[4] - [28].
| Relations des Eupleridae avec les autres familles proches dans l'ordre des Feliformia[29] : | |
malgaches (Eupleridae)[30] :
|
En 1993, Géraldine Veron et François Catzeflis publient une étude utilisant l'hybridation de l'ADN et suggérant que le fossa est plus étroitement lié aux mangoustes (la famille des Herpestidae) qu'aux félins ou aux civettes[3] - [28]. Cependant une étude morphologique de Veron publiée deux ans plus tard replace à nouveau le fossa parmi la famille des Felidae[28]. En 2003, des études moléculaires phylogénétiques utilisant des gènes nucléaires et mitochondriaux menées par Anne Yoder et ses collègues ont montré que tous les carnivores malgaches indigènes sont un groupe monophylétique, partageant un ancêtre commun distinct des lignées des autres carnivores (ils forment un clade) et qu'ils sont le plus étroitement liés aux Herpestidae d'Afrique et d'Asie[30] - [31] - [29]. Pour tenir compte de ces relations, tous les carnivores malgaches sont maintenant placés dans une famille propre, celle des Eupleridae[32]. Dans cette famille le fossa est placé dans la sous-famille des Euplerinae, avec le fanalouc (Eupleres goudoti) et la civette malgache (Fossa fossana), mais ses relations exactes sont mal résolues[32] - [30] - [29].
Une espèce éteinte et proche du fossa est décrite en 1902 à partir de restes subfossiles, et décrite sous le nom de Cryptoprocta spelea en 1935. Cette espèce était similaire au fossa d'aujourd'hui mais plus grande (avec une estimation d'une masse corporelle à peu près deux fois plus importante)[4] - [33]. À Madagascar les gens distinguent plusieurs types de fossas : le grand fosa mainty (« fossa noir ») et le fosa mena (« fossa rougeâtre ») plus petit. Une forme blanche a été signalée dans le sud-ouest de l'île. Il est difficile de savoir si cela est purement folklorique, reflète une variation individuelle liée au sexe, à l'âge ou à des cas de mélanisme ou de leucisme ou s'il y a en effet plus d'une espèce de fossa vivante[4] - [33] - [2].
Le fossa et l'homme
Menaces et conservation
L'Union internationale pour la conservation de la nature (UICN) considère le fossa comme « vulnérable » (VU) depuis 2008 car sa population a probablement diminué d'au moins 30 % au cours des 21 dernières années ; il était précédemment considéré comme « en danger » (EN) en 2000, vulnérable en 1986 et 1996 et les « données insuffisantes » (DD) en 1988, 1990 et 1994[1]. L'espèce est dépendante de l'habitat forestier et est donc menacée par la destruction généralisée des forêts naturelles de Madagascar, mais elle est également capable de persister dans les zones perturbées[3] - [2]. Une série de marqueurs microsatellites (de courts segments d'ADN ayant une séquence répétée) a été développée pour aider les études de la santé génétique et la dynamique des populations chez le fossa à la fois à l'état sauvage et en captivité[34]. Plusieurs agents pathogènes ont été isolés à partir du fossa, dont certains, comme l'anthrax et la maladie de Carré, pourraient être transmis par les chiens ou les chats errants[2].
Bien que l'espèce soit largement répandue à Madagascar, elle est localement rare dans toutes les régions de l'île et ainsi particulièrement vulnérable face à l'extinction ; la fragmentation écopaysagère est un facteur aggravant le risque. Le fossa a une densité de population plus faible que l'on pourrait prédire à partir de la taille de l'animal, et est en outre menacé par la disparition rapide des forêts malgaches et le déclin des populations de lémuriens qui constituent une grande part de son alimentation. La disparition du fossa, localement ou totalement, pourrait grandement influer sur la dynamique des écosystèmes, conduisant éventuellement au surpâturage par certaines de ses proies. La population totale dans les aires protégées est estimée à moins de 2 500 adultes, mais ce chiffre est peut-être surestimé. Seules deux zones protégées contiendraient plus de 500 fossas adultes : le parc national de Masoala et le parc national de Midongy du sud, même si ces estimations pourraient également être surévaluées. Les informations recueillies sur les populations sont insuffisantes pour une analyse de viabilité des populations correcte, mais les estimations donnent à penser qu'aucune des aires protégées ne soutient une population viable. Si ce résultat est correct, il impliquerait un processus de déclin progressif des effectifs sur une centaine d'années, jusqu'à l'extinction de l'espèce. Pour que celle-ci survive, il est estimé qu'au moins 555 km2 sont nécessaires pour maintenir de petites populations viables sur le court terme, et au moins 2 000 km2 pour les populations de 500 adultes[21].
Dans la culture
Un tabou, nommé fady à Madagascar[35], offre une protection au fossa et à d'autres carnivores[36]. Dans le district de Marolambo (province de Tamatave), le fossa était traditionnellement haï et craint comme un animal dangereux. Il est décrit comme « avide et agressif », connu pour ses vols de volailles et de porcelets, et pour « prendre les petits enfants qui marchent seuls dans la forêt ». Certains ne le mangeaient pas, de peur que la bête ne transfère ses qualités indésirables à la personne qui la consomme[35]. À l'inverse l'animal était parfois pris comme viande de brousse[2] ; une étude publiée en 2009 rapportait que 57 % des villages (8 dans un échantillon de 14) dans la forêt de Makira consommaient de la viande de fossa. Les animaux sont généralement chassés à l'aide de frondes, de chiens, ou plus couramment en plaçant des collets sur les chemins qu'ils empruntent[37]. Près du parc national de Ranomafana, le fossa et plusieurs de ses cousins plus petits, ainsi que la civette indienne (Viverricula indica), introduite, sont connus pour se « nourrir sur les corps des ancêtres », qui sont enterrés dans la forêt dans des fosses peu profondes. Pour cette raison, la consommation de ces animaux est strictement interdite par le fady. Cependant, s'ils errent dans les villages à la recherche d'oiseaux domestiques, ils peuvent être tués ou pris au piège. Des pièges à petits carnivores ont été observés près des enclos de poules dans le village de Vohiparara[36].
Quelques zoos possèdent des fossas. La première reproduction en captivité a eu lieu en 1974 au parc zoologique de Lunaret, en France. En 1975, alors qu'il n'y avait que huit fossas en captivité à travers le monde, le zoo de Duisbourg, en Allemagne, a acquis un spécimen ; il a lancé par la suite un programme de reproduction, avec succès : la plupart des fossas de zoos actuels descendent des fossas de Duisbourg. Ce zoo a également fourni de nombreuses données sur les spécimens captifs[5].
Liens externes
Bibliographie
- (en) Edward Turner Bennett, « Notice of a new genus of Viverridous Mammalia from Madagascar », Proceedings of the Zoological Society of London, , p. 46 (lire en ligne)
- (en) Andrew Smith, « Contributions to the natural history of South Africa », South African Quarterly Journal, vol. 2, no 3, , p. 246-247 (lire en ligne)
- (en) Donald J. Borror, Dictionary of Word Roots and Combining Forms, Mayfield Publishing Company, , 134 p. (ISBN 978-0-87484-053-7)
- (en) Jörgen Ruud, Taboo : A Study of Malagasy Customs and Beliefs, Oslo University Press, , 2e éd.
- (fr) Roland Albignac, Faune de Madagascar, vol. 36 : Mammifères. Carnivores, , 206 p.
- (en) M. Köhncke et K. Leonhardt, « Cryptoprocta ferox », Mammalian Species, no 254, , p. 1–5 (lire en ligne)
- (en) Steven M. Goodman (dir.) et Jonathan P. Benstead (dir.), The Natural History of Madagascar, Chicago (Ill.), University of Chicago Press, , 1709 p. (ISBN 0-226-30306-3), chap. 13 (« Mammals »)
- (en) Anne D. Yoder et John J. Flynn, « Origin of Malagasy Carnivora », dans , , p. 1253–1256
- (en) Clare E. Hawkins, « Cryptoprocta ferox, Fossa, Fosa », dans , , p. 1360–1363
- (en) Nick Garbutt, Mammals of Madagascar, A Complete Guide, A&C Black Publishers, , 304 p. (ISBN 978-0-300-12550-4)
- (en) Luke Dollar, Jörg U. Ganzhorn et Steven M. Goodman, « Primates and other prey in the seasonally variable diet of Cryptoprocta ferox in the dry deciduous forest of Western Madagascar », dans Sharon L. Gursky et Anna Nekaris, Primate Anti-Predator Strategies (Developments in Primatology: Progress and Prospects), Springer, (ISBN 978-0-387-34807-0), p. 63-76
- (en) David W. Macdonald, The Princeton Encyclopedia of Mammals, Princeton, N.J., Princeton University Press, , 936 p. (ISBN 978-0-691-14069-8)
- (en) Steven M. Goodman, « Family Eupleridae (Madagascar Carnivores) », dans D. Wilson et Russell Alan Mittermeier, Handbook of the Mammals of the World, vol. 1 : Carnivores, Lynx Edicions, (ISBN 978-84-96553-49-1, lire en ligne)
Références taxinomiques
- (en) Référence Mammal Species of the World (3e éd., 2005) : Cryptoprocta ferox Bennett, 1833 (consulté le )
- (en) Référence Catalogue of Life : Cryptoprocta ferox Bennett, 1833 (consulté le )
- (en) Référence CITES : espèce Cryptoprocta ferox Bennett, 1833 (+ répartition sur Species+) (consulté le )
- (fr+en) Référence ITIS : Cryptoprocta ferox Bennett, 1833 (consulté le )
Liens externes
- (en) Référence Animal Diversity Web : Cryptoprocta ferox (consulté le )
- (en) Référence NCBI : Cryptoprocta ferox (taxons inclus) (consulté le )
- (en) Référence UICN : espèce Cryptoprocta ferox Bennett, 1833 (consulté le )
- (fr) Référence CITES : taxon Cryptoprocta ferox (sur le site du ministère français de l'Écologie) (consulté le )
- (en) Référence Fonds documentaire ARKive : Cryptoprocta ferox (consulté le )
- Aves sur le Fossa
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Fossa (animal) » (voir la liste des auteurs).
- Union internationale pour la conservation de la nature, consulté le 2 avril 2012
- Goodman (2009)
- Hawkins (2003), p. 1360–1363
- Köhncke et Leonhardt (1986)
- (en) A. Winkler, « Neueste Erkenntnisse zur Biologie, Haltung und Zucht der Fossa (Cryptoprocta ferox) », Der Zoologische Garten, vol. 73, no 5, , p. 296–311
- (en) J. Mueller, A. Sironen et K.E. Lukas, « Infant development and behavior in the Fossa Cryptoprocta ferox », Small Carnivore Conservation, vol. 37, , p. 11–17 (lire en ligne)
- Macdonald (2009), p. 668–669
- (en) C.M. Drea, N.J. Place, M.L. Weldele, E.M. Coscia, P. Licht et S.E. Glickman, « Exposure to naturally circulating androgens during foetal life incurs direct reproductive costs in female spotted hyenas, but is prerequisite for male mating », Proceedings of the Royal Society B, vol. 269, no 1504, , p. 1981–1987 (PMID 12396496, DOI 10.1098/rspb.2002.2109, lire en ligne)
- (en) L. Dollar, « Preliminary report on the status, activity cycle, and ranging of Cryptoprocta ferox in the Malagasy rainforest, implications for conservation », Small Carnivore Conservation, vol. 20, , p. 7–10 (lire en ligne)
- (en) E.R. Patel, « Silky Sifaka predation (Propithecus candidus) by a Fossa (Cryptoprocta ferox) », Lemur News, vol. 10, , p. 25–27 (lire en ligne)
- (en) P.C. Wright, « Demography and life history of free ranging Propithecus diadema Edwardsi in Ranomafana National Park, Madagascar », International Journal of Primatology, vol. 16, no 5, , p. 835–854 (DOI 10.1007/BF02735722, lire en ligne)
- (en) P.C. Wright, S.K. Heckscher et A.E. Dunham, « Predation on Milne Edward's sifaka (Propithecus diadema edwardsi) by the fossa (Cryptoprocta ferox) in the rainforest of southeastern Madagascar », Folia Primatologica, vol. 68, no 1, , p. 34–43 (DOI 10.1159/000157230)
- (en) C.E. Hawkins et P.A. Racey, « Food habits of an endangered carnivore, Cryptoprocta ferox, in the dry deciduous forests of western Madagascar », Journal of Mammalogy, vol. 89, no 1, , p. 64–74 (DOI 10.1644/06-MAMM-A-366.1)
- (en) S.M. Goodman, « The carnivores of the Reserve Naturelle Integrale d'Andringitra, Madagascar », Fieldiana Zoology, no 85, , p. 289–292 (ISSN 0015-0754)
- (en) S.M. Goodman et M. Pidgeon, « Carnivora of the Reserve Naturelle Integrale d'Andohahela, Madagascar », Fieldiana Zoology, no 94, , p. 259–268 (ISSN 0015-0754)
- (en) R.M. Rasoloarison, B.P.N. Rasolonandrasana, J.U. Ganzhorn et S.M. Goodman, « Predation on vertebrates in the Kirindy Forest, western Madagascar », Ecotropica, vol. 1, , p. 59–65 (lire en ligne, consulté le )
- (en) M.T. Irwin, J.L. Raharison et P.C. Wright, « Spatial and temporal variability in predation on rainforest primates: do forest fragmentation and predation act synergistically? », Animal Conservation, vol. 12, no 3, , p. 220–230 (DOI 10.1111/j.1469-1795.2009.00243.x)
- Dollar, Ganzhorn et Goodman (2007), p. 63–76
- (en) C.E. Hawkins et P.A. Racey, « A novel mating system in a solitary carnivore: the fossa », Journal of Zoology, vol. 277, , p. 196–204 (DOI 10.1111/j.1469-7998.2008.00517.x)
- vidéo sur youtube.com
- (en) C.E. Hawkins et P.A. Racey, « Low population density of a tropical forest carnivore, Cryptoprocta ferox: implications for protected area management », Oryx, vol. 39, no 1, , p. 35–43 (DOI 10.1017/S0030605305000074)
- (en) M.-L. Lührs et M. Dammhahn, « An unusual case of cooperative hunting in a solitary carnivore », Journal of Ethology, vol. 28, no 2, , p. 379–383 (DOI 10.1007/s10164-009-0190-8)
- Garbutt (2007), p. 211–214
- Albignac (1973), p. 1–206
- Borror (1960), p. 39
- (en) R.M. Blench et M. Walsh, Faunal names in Malagasy : their etymologies and implications for the prehistory of the East African coast, Aussois, France, Eleventh International Conference on Austronesian Linguistics (11 ICAL), , 31 p. (lire en ligne)
- Bennett (1833), p. 46
- Yoder et Flynn (2003), p. 1253–1256
- (en) E. Barycka, « Evolution and systematics of the feliform Carnivora », Mammalian Biology, vol. 72, no 5, , p. 257–282 (DOI 10.1016/j.mambio.2006.10.011)
- (en) A.D. Yoder, M.M. Burns, S. Zehr, T. Delefosse, G. Veron, S.M. Goodman et J.J. Flynn, « Single origin of Malagasy Carnivora from an African ancestor », Nature, vol. 421, no 6924, , p. 734–737 (PMID 12610623, DOI 10.1038/nature01303, lire en ligne)
- (en) G. Veron, M. Colyn, A.E. Dunham, P. Taylor et P. Gaubert, « Molecular systematics and origin of sociality in mongooses (Herpestidae, Carnivora) », Molecular Phylogenetics and Evolution, vol. 30, no 3, , p. 582–598 (PMID 15012940, DOI 10.1016/S1055-7903(03)00229-X, lire en ligne)
- Mammal Species of the World, consulté le 2 avril 2012
- (en) S.M. Goodman, R.M. Rasoloarison et J.U. Ganzhorn, « On the specific identification of subfossil Cryptoprocta (Mammalia, Carnivora) from Madagascar », Zoosystema, vol. 26, no 1, , p. 129–143 (lire en ligne)
- (en) B.R. Vogler, C.A. Bailey, G.D. Shore, S.M. McGuire, S.E. Engberg, J. Fickel, E.E. Louis Jr. et R.A. Brenneman, « Characterization of 26 microsatellite marker loci in the fossa (Cryptoprocta ferox) », Conservation Genetics, vol. 10, no 5, , p. 1449–1453 (DOI 10.1007/s10592-008-9758-z)
- Ruud (1970), p. 101
- J.P.G. Jones, M.A. Andriamarovolona et N.J. Hockley, « Taboos, social norms and conservation in the eastern rainforests of Madagascar »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?), 9th International BIOECON Conference on "Economics and Institutions for Biodiversity Conservation",
- (en) C.D. Golden, « Bushmeat hunting and use in the Makira Forest, north-eastern Madagascar: a conservation and livelihoods issue », Oryx, vol. 43, , p. 386–392 (DOI 10.1017/S0030605309000131)