Domaine vital
Le domaine vital est l’aire où un animal vit ordinairement et qui suffit à répondre à ses besoins primaires.
On peut parler du domaine vital d'un individu moyen, d'un couple type ou éventuellement d'une population au sein d'une métapopulation, pour tout ou partie de son cycle de vie (le domaine vital doit donc être défini pour un intervalle de temps donné (saison, année, ou durée de vie [1]).
Disposer d'un territoire et d'un domaine vital a un « coût » pour l'animal (ainsi rendu « dépendant » d'une aire particulière) et parfois un coût pour le groupe (risque plus élevé de surexploitation de l'aire, et de parasitose), mais procurerait selon Stamps (1995)[2] des bénéfices dépassant ces coûts pour les animaux territoriaux. Ces animaux explorent et exploitent mieux (à moindres coûts énergétiques) un environnement devenu familier. Par exemple, les coûts quotidiens d'alimentation, de surveillance, de maintien, de défense, de développement et mémorisation des ressources vitales et critiques sur lesquelles il base ses décisions pour établir un domaine vital, doivent être inférieurs ou égaux aux bénéfices acquis grâce au domaine vital. La proie pourrait sembler plus vulnérable à la prédation, mais connaissant mieux son territoire, elle peut mieux fuir ou s'y cacher, de plus, les pistes odorantes y sont en partie brouillées pour le prédateur qui les utilise.
Il existe des cas particuliers, dont :
- le domaine d'espèces bioconstructrices ;
- le domaine d'espèces-ingénieurs qui aménagent littéralement tout ou partie de leur territoire (ex : certains termites ou le castor et ses barrages) ;
- les espèces utilisant des places de parades sexuelles (sans intérêt alimentaire ou de cachette…) ; ex : paradisiers…
Éléments de définition

- Pour Burt (1943), le domaine vital est « la surface traversée par l’individu pendant ses activités normales de recherche de nourriture, de reproduction et de soin aux jeunes. Des sorties occasionnelles hors de la zone, telles que des explorations dans la nature, ne devant pas être considérées comme faisant partie du domaine vital »[3].
Cette définition en exclut non pas les migrations, mais tout déplacement important et inhabituel[4].
- La définition de Burt a été modifiée dans les années 1970, l'aire vitale étant alors estimée comme la probabilité de trouver un animal à une localisation particulière d’un plan[5] - [6] - [7].
- Pour les logiciels de calcul, cette probabilité est une « fonction de densité » appelée « Distribution d’Utilisation » (UD). Elle correspond généralement à la distribution des positions d’un animal dans le plan. « Un critère de niveau de probabilité peut être utilisé pour éliminer les sorties occasionnelles décrites par Burt (1943), en incluant seulement dans le domaine vital la plus petite aire où l’animal passe 95 % de son temps. Utiliser 95 % des domaines vitaux est en accord avec le seuil de probabilité communément admis pour valider statistiquement une hypothèse. Les données fréquemment utilisées pour quantifier le domaine vital d’un animal sont sous la forme d’observations visuelles ou déduites (localisations télémétriques ou traces) »[8].
- La notion de domaine vital ne doit pas être confondue avec celle de « territoire » d'un animal territorial, qui est la part du domaine vital défendu [9]) ; Les milieux non considérés comme « aires d’utilisation » sont généralement exclus de l'aire vitale, bien que des animaux puissent être familiers d'aires qu’ils n’utilisent pas.
Sauf cas particuliers (éléments du territoire utilisés comme point de marquage odorant ou visuel), la notion d'aire vitale ne prend pas non plus en compte (ou mal) le comportement de défense du territoire de la part de l'animal. - Pour les animaux moyennement ou peu territoriaux, selon Powell (2000), les frontières et les aires des domaines vitaux pourraient être floues, car peut-être imprécises pour les animaux eux-mêmes[1].
- La taille et la configuration du domaine vital sont généralement déterminées par l'espace nécessaire pour répondre aux besoins en alimentaires de l’animal au moment de la reproduction et le cas échéant de l'accumulation de graisses en prévision de l'hivernage, d'une hibernation ou d'une migration. Cet espace varie selon les ressources locales, qui peuvent elles-mêmes varier selon les années (glandaies par exemple).
Par conséquent, plus un animal est gros, ou plus sa ressource alimentaire est rare ou éparpillée, plus son « domaine vital » aura tendance à être étendu[10]. - Au sein d'un territoire et d'une aire vitale observé pour la durée totale de vie d'un animal, le domaine vital peut changer d'une année sur l'autre (ex : pour le chevreuil (réputé très sédentaire[11] - [12])étudié par Pellerin (2005), le recouvrement d'une année sur l'autre était d'environ 60 % pour chaque individu [8]).
Déductions
L'analyse déductive de l'importance de la taille du domaine vital doit être interprétée selon l'espèce et à partir de différents facteurs : elle varie d’une espèce à l’autre, par exemple selon :
- le sexe (mâle et la femelle et différemment selon l'espèce) ;
- la période de l’année (qui module la disponibilité en ressources alimentaires et la capacité thermique d'accueil du milieu (en zone froide ou aride notamment) ;
- le poids de l'animal[13] ;
- l'âge[14] ;
- la partie concernée du cycle de vie de l’animal (subaquatique chez le têtard, mais aérienne et terrestre chez le crapaud adulte) ;
- la place dans la chaîne alimentaire. Les carnivores et grands prédateurs nécessitent généralement un territoire plus grand que les consommateurs primaires.
Ainsi, l'aire vitale de l'élan (Alces alces) montre une corrélation positive avec l’âge pour le mâle, qui reflète dans ce cas la dominance (Cederlund et Sand 1994), alors que la taille des domaines annuels et hivernaux diminuera avec l’âge chez la femelle du chevreuil ou du bouquetin révélant cette fois l’expérience du territoire acquise par la femelle au fur et à mesure du temps[15].
Les « domaines » des individus d’une même espèce peuvent se chevaucher.
S’il y a défense du territoire, les domaines vitaux des animaux ne se chevauchent généralement pas, mais il peut y avoir chevauchement lorsque les animaux ne se rencontrent pas ou exceptionnellement ou encore si le territoire défendu est plus petit que le domaine vital.
Histoire du concept
Cette notion semble être utilisée par les biologistes et éthologues au moins depuis la publication de 1943, de W.H. Burt[16], qui dessinait des cartographies de l'étendue de l'aire de vie des espèces qu'il étudiait et des limites extérieures des mouvements effectués par ces animaux au cours de leurs activités quotidiennes.
Divers auteurs ont associé au concept du domaine vital les notions de patterns (patrons) de distribution géographique dans l'aire de répartition[17] - [18] qui représentent par exemple la probabilité de trouver un animal dans une zone définie dans son domaine vital. La zone de vie d'un animal individuel est généralement construite à partir d'un ensemble de points de localisation qui ont été collectés sur une certaine période de temps. Elle identifie la position dans l'espace d'un individu pour plusieurs points dans le temps. Des données beaucoup plus nombreuses et précises sont maintenant recueillies automatiquement via des colliers, balises ou puces électroniques placés sur les individus, qui transmettent ces informations notamment par le biais des satellites ou en utilisant la technologie cellulaire mobile et les systèmes de positionnement global (GPS), à intervalles réguliers.
Utilisations et enjeux
Enjeux
La notion de domaine vital est mobilisée par les éthologues, les écologues (écologie du paysage), et pour l'étude du fonctionnement des populations animales (de vertébrés notamment).
- Elle est indispensable pour définir les règles de conservation ou « dimensionner » un projet d'exploitation durable de cette population[8]. C'est un des éléments de la chorologie (l'étude explicative de la répartition géographique des espèces vivantes et de ses causes).
Ils concernent notamment la biologie de la conservation ;
- Il existe un lien entre la taille de l'aire vitale et les chances de survie. On a montré chez le chevreuil que les femelles (chevrettes) âgées peuvent diminuer leur aire vitale, probablement grâce à « l'expérience acquise par rapport à leur territoire et la connaissance des patches alimentaires riches au fil des mois » (Pellerin citant Stamps 1995[19], Dzieciolowski et al. 1998[20], Stamps et Krishnan 1999[2], Gautestad et Mysterud 2005[21]). Ceci laisse penser que le rajeunissement artificiellement entretenu par la chasse ou le braconnage (des trophées notamment) de certaines populations peut avoir une incidence importante sur la taille de leur aire vitale et par suite sur leur survie sur des territoires de plus en plus petits et écologiquement fragmentés.
- Comprendre et connaitre les contraintes et besoins en termes de « domaines vitaux » pour les espèces est nécessaire pour le bon positionnement et dimensionnement des corridors biologiques, des réservoirs de biodiversité et de certains écoducs dans le cadre de la protection ou restauration des réseaux écologiques, via la trame verte et bleue en France par exemple.
utilisations du concept
L'étude du domaine vital complète d'autres approches, ou y participe, dont :
- le suivi de population par capture-marquage-recapture ;
- le suivi d'individus par radio-tracking (radio-pistage) ;
- l'indice de Pression sur la Flore (I.P.F. de 1991 à 1997) ;
- l'indice de Consommation des ressources(I.C.) ;
- l'étude de la qualité de l'habitat sur la dynamique des populations par exemple chez des cervidés comme le chevreuil[22] - [23] - [24] - [25] - [26] et des traits d'histoire de vie[27] ;
- l'étude des densités et des mouvements d'individus.
Cartographie du domaine vital
Conditions initiales :
- Dans tous les cas, le cartographe et/ou le modélisateur doivent disposer de données géoréférencées acquises pour un échantillon d'une population statistiquement représentative de l'espèce que l'on veut étudier ;
Ces données sont parfois difficiles à collecter sur le terrain (par exemple pour les jeunes reptiles ou les jeunes amphibiens (ex : jeunes tritons) qui échappent facilement à l'observateur. Des études en cours ou récentes, utilisant des puces RFID ou dispositifs très légers de « tracking » devraient améliorer la connaissance de l'écologie et des déplacements de ces "juvéniles" qu'on ne ré-observe généralement à ce jour qu'au moment de la reproduction et qui avant cela échappent aux méthodes de type CMR (Capture-Marquage-Recapture) ; - Une caractérisation tridimensionnelle de la donnée peut être utile pour les études d'espèces vivant dans les océans ou les lacs, voire pour des espèces vivant dans plusieurs strates de l'étage ou du sous-étage forestier ;
- Des cartes de plus en plus précises des habitats ou milieux naturels et parfois librement disponibles en Open data (ex : projet ARCH[28]) peuvent parfois être d'emblée intégrées dans le SIG.
Visualisation d'Information concernant le domaine vital :
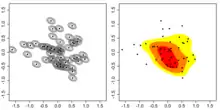
- La façon la plus simple de tracer les « frontières » (les limites) d'un domaine vital à partir d'un ensemble de données géoréférencées de localisation (« semis de points » disposés sur une carte) est de construire le polygone convexe le plus petit possible autour de ces points.
Cette approche est appelée le « polygone convexe minimum » (ou « MCP » pour « Minimum convex polygon ». Cette méthode est encore largement employée[29] - [30] - [31] - [32]mais avec de nombreux biais et inconvénients, dont selon Burgman celui d'une surestimation fréquente de la taille des domaines vitaux[33].
- Les méthodes les plus connues pour la construction de cartes de domaine vital et de distributions d'utilisation de l'espace sont dites estimations par le noyau bivariée gaussienne ou normale (bivariate Gaussian or normal distribution kernel density methods)[34] - [35] - [36].
Ce groupe de méthodes fait partie d'un groupe plus général de méthodes paramétriques du noyau.
- Récemment, l'approche par le noyau a fait l'objet d'amélioration avec l'inclusion d'un certain nombre de méthodes non-paramétriques telles que
Logiciels et modélisation
De nombreux logiciels sont actuellement disponibles pour ce type de calcul que les Systèmes d'information géographique (SIG, éventuellement libres tels que SIG libre dont GRASS, QGIS) ont facilité ; les plus nombreux concernent les méthodes paramétriques (en raison de l'utilisation plus récente des méthodes non paramétriques. Selon une évaluation de Getz et al., les études disponibles démontrent que les méthodes « LoCoh » fournissent généralement des estimations plus exactes de la taille des domaines vitaux et présentent moins de biais que les méthodes paramétriques du noyau.
Des progiciels (software) pour la mise en œuvre des méthodes paramétriques et non paramétriques du noyau sont disponibles en ligne, dont :
- HRT : Home Range Tools pour ArcGIS 9.x (ESRI) (méthodes d’estimation : MCP et kernel)[39] ;
- LoCoH pour déterminer l'aire vitale d'une espèce[40] ;
- HoRAE (en open source), pour étudier le domaine vital [41] ;
- le logiciel R d'analyse spatiale (du projet AniMove visant à étudier les mouvements des animaux[42]) avec une suite de paquets (packages) spécifiques (« Adehabitat » et autres).
Ces outils permettent généralement au moins :
- la gestion des points, lignes et polygones et des cartes raster ;
- la gestion de données "individuelles" issues du radio-pistage (de plus en plus souvent par GPS) ; analyse factorielle des rapports de sélection, analyse OMI, analyse K-select, etc. ;
- l'analyse des semis de points[43] ;
- l'estimation surfacique et géographique du domaine vital ;
- estimation de la distance de Mahalanobis ;
- une analyse spatiale plus fine de la sélection de l’habitat par la faune sauvage, avec notamment l'analyse de la sélection de certaines ressources vitales (ou non vitales) ;
- l'analyse de dispersion par l'étude des « trajets »[44] - [45] incluant la représentation des distances entre localisations, les « turning angles », et l'intégration éventuelle de données d'activité ou 3D (profondeur sous l'eau pour un animal en plongée, élévation dans l'atmosphère pour un oiseau), etc., avec valorisation graphique -dynamique ou non) de la donnée « trajet » (« Trajdyn », « Plotltr », etc. ) ; très utile pour les études de fragmentation écopaysagère (ex : Fragmentation forestière), de corridors biologiques et la cartographie des corridors biologiques et des réservoirs de biodiversité et de leurs zones tampons, dans le cadre de la trame verte et bleue par exemple), pour vérifier la cohérence écologique d'une gestion ou d'un aménagement, ou encore pour révéler des situations de piège écologique ou de puits écologiques.
- le changement de projection (la carte reste cohérente dans un autre système de projection cartographique)
- le calcul de l'utilization distribution (UD).
Limites : Ce sont surtout en amont la pression d'échantillonnage[46] et en aval la mémoire informatique et la puissance de calcul.
Voir aussi
Articles connexes
- Éthologie
- Comportement
- Espace vital
- Fragmentation du territoire (ex : fragmentation des forêts
- Habitat naturel
- Niche écologique
- Ressource naturelle
- écologie du paysage
- Réservoir de biodiversité, corridor biologique
- cartographie des corridors biologiques
- Applications des systèmes d'information géographique
- Système d'information taxonomique intégré
- Théories de distribution des espèces
- Théorie source-puits
Liens externes
- (en) liste de discussion spécialisées R-sig-Geo (R Special Interest Group on using Geographical data and Mapping)
Bibliographie
- (fr) Clément Calenge (ONCFS) L'analyse de l'utilisation de l'espace par la faune sauvage ; Séminaire, du Muséum d'Histoire naturelle national de Paris, 2009-05-14 (présentation PPT/PDF, 73 pp)
- Urbano, Cagnacci, Calenge, Cameron, Dettki & Neteler. In prep. How not to drown in data : a new approach for managing wildlife location data.
- Cagnacci & Urbano. 2008. Managing wildlife : A spatial information system for GPS collars data. Environmental Modelling & Software 23. 957-959.
Références
- Powell, R. A. 2000. Animal home ranges and territories and home range estimators. Research Techniques in Animal Ecology: Controverses and Consequences (eds L. Boitoni and T. K. Fuller), Colombia University Press, New York
- Stamps, J. A., and V. V. Krishnan. 1999. A learning-based model of territory establishment. Quaterly Review of Biology 74:291-318.
- Burt, V. H. 1943. Territoriality and home range as applied to mammals. Journal of Mammalogy 24:346-352
- (en) P. R. Moorcroft, « Animal Home Ranges », dans Jørgensen, Sven Erik et Brian D. Fath, Encyclopedia of ecology, Amsterdam ; London, Academic Press, , 174-180 p. (ISBN 9780080454054, présentation en ligne).
- Jennrich, R. I., and F. B. Turner. 1969. Measurement of non-circular home range. Journal of Theoretical Biology 22:227-237.
- Dunn, J. E., and P. S. Gipson. 1977. Analysis of radio telemetry data in studies of home range. Biometrics 33:85-101.
- Anderson, D. J. 1982. The home range: A new non-parametric estimation technique. Ecology 63:103-112.
- Thèse de Maryline Pellerin (doctorat en Physiologie, Biologie des organismes, Populations, Interactions, Université de Poitiers), Utilisation et sélection de l'Habitat chez le chevreuil à différentes échelles spatio-temporelles (2005-12-09), 290 pages
- (en) J. L. Rachlow, « Wildlife Ecology », dans Jørgensen, Sven Erik et Brian D. Fath, Encyclopedia of ecology, Amsterdam ; London, Academic Press, , 3790-3794 p. (ISBN 9780080454054, présentation en ligne)
- David McFarland (trad. de l'anglais par Guy Schoeller), Dictionnaire du comportement animal, Paris, Laffont, , 1013 p. (ISBN 2-221-05281-1, OCLC 22441745).
- Strandgaard, H. 1972: The roe deer (Capreolus capreolus) population at Kalo and the factors regulating its size. - Danish Review of Game Biology 7:1-205.
- Liberg, O., A. Johansson, R. Andersen, and J. D. C. 1998. The function of male territoriality in roe deer. The European roe deer: the biology of a success (eds R. Andersen, P. Duncan and J. D. C. Linnell), Scandinavian University Press, p. 221-256.
- Harestad, A. S., and F. L. Bunnell. 1979. Home range and body weight – a reevaluation. Ecology 60:389-402.
- Pettorelli, N., J. M. Gaillard, P. Duncan, D. Maillard, G. Van Laere, and D. Delorme. 2003. Age and density modify the effects of habitat quality on survival and movements of roe deer. Ecology 84:3307-3316
- S., F. Parrini, B. Bassano, S. Siriano, and M. Apollonio. 2003. Habitat selection in adult males of Alpine ibex, Capra ibex ibex. Folia Zoologica 52:113–120.
- Burt, W. H. 1943. Territoriality and home range concepts as applied to mammals. Journal of Mammalogy 24:346–352.
- ennrich, R. I. and Turner F. B. 1969. Measurement of non-circular home range. J. Theoretical Biology 22:227-237
- Ford, R. G. and Krumme D. W. 1979. The analysis of space use patterns. - J. Theoretical Biology 76:125-157.
- Stamps, J. A. 1995. Motor learning and the value of familiar space. The American Naturalist 146:41-58.
- Dzieciolowski, R., M. Wasilewski, J. Przypasniak, P. Havet, E. Taran, and J. C. Berthos 1998. Home ranges of roe deer (Capreolus capreolus) inhabiting a fine-grained landscape. Gibier Faune Sauvage 15:555-563.
- Gautestad, A. O., and I. Mysterud. 2005. Intrinsic scaling complexity in animal dispersion and abundance. The American Naturalist 165:44-55.
- Coulson, T., S. D. Albon, F. E. Guinness, J. Pemberton, and T. H. Cluton-Brock. 1997. Population substructure, local density, and calf winter survival in red deer. Ecology 78:852-863.
- Coulson, T., S. D. Albon, J. Pilkington, and T. H. Cluton-Brock. 1999. Small-scale spatial dynamics in a fluctuating ungulate population. Journal of Animal Ecology 68:658-671
- Milner-Gulland, E. J., T. N. Coulson, and T. H. Clutton-Brock. 2000. On harvesting a structured ungulate population. Oikos 88:592-602.
- Pettorelli, N., J. M. Gaillard, P. Duncan, J. P. Ouellet, and G. Van-Laere. 2001. Population density and small-scale variation in habitat quality affect phenotypic quality in roe deer. Oecologia 128:400-405
- Pettorelli, N., J. M. Gaillard, G. Van Laere, P. Duncan, P. Kjellander, O. Liberg, D. Delorme, and D. Maillard. 2002. Variations in adult body mass in roe deer: the effects of population density at birth and of habitat quality. Proceedings of the Royal Society of London series B 269:747-753)
- Stearns, S. C. 1992. The evolution of life histories. Oxford University Press, Oxford.
- Projet européen Arch (Interreg) intitulé Assessing Regional Habitat Change, associant les régions européennes, anglaise et française du Kent et du Nord-Pas-de-Calais au Centre régional de phytosociologie agréé conservatoire botanique national de Bailleul, avec la DREAL Nord-Pas-de-Calais
- Baker, J. 2001. Population density and home range estimates for the Eastern Bristlebird at Jervis Bay, south-eastern Australia. – Corella 25:62-67
- Creel, S. and Creel N. M. 2002. The African Wild Dog: Behavior, Ecology, and Conservation. - Princeton University Press, Princeton, New Jersey, 341 p.
- Meulman, E. P. and Klomp N. I. 1999. Is the home range of the heath mouse Pseudomys shortridgei an anomaly in the Pseudomys genus? - Victorian Naturalist. 116:196-201
- Rurik, L. and Macdonald D. W. 2003. Home range and habitat use of the kit fox (Vulpes macrotis) in a prairie dog (Cynomys ludovicianus) complex. - J. Zoology, 259:1-5
- Burgman, M. A. and Fox J. C. 2003. Bias in species range estimates from minimum convex polygons: implications for conservation and options for improved planning. -Animal Conservation 6:19-28
- Silverman, B. W. 1986. Density estimation for statistics and data analysis. - Chapman and Hall, London, UK
- Worton, B. J. 1989. Kernel methods for estimating the utilization distribution in home-range studies. - Ecology 70:164–168.
- Seaman, D. E. and Powell R. A. 1996. An evaluation of the accuracy of kernel density estimators for home range analysis. - Ecology 77:2075–2085
- Getz, W. M. and C. C. Wilmers, 2004. A local nearest-neighbor convex-hull construction of home ranges and utilization distributions. Ecography 27:489-505.
- Getz, W.M, S. Fortmann-Roe, P. C. Cross, A. J. Lyonsa, S. J. Ryan, C.C. Wilmers, in review. LoCoH: nonparametric kernel methods for constructing home ranges and utilization distributions.
- Rodgers, A.R., A.P. Carr, H.L. Beyer, L. Smith, and J.G. Kie. 2007. HRT: Home Range Tools for ArcGIS. Version 1.1. Ontario Ministry of Natural Resources, Centre for Northern Forest Ecosystem Research, Thunder Bay, Ontario, Canada
- LoCoH: Powerful algorithms for finding home ranges
- HoRAE - Home Range Analysis and Estimation Toolbox (open source)
- AniMove - Animal movement methods
- Calenge (2005) Des outils statistiques pour l’analyse des semis de points dans l’espace écologique Université Claude Bernard Lyon 1.
- Calenge (2006) Ecological Modelling, 197 :516-519.
- Martin, Calenge, Quenette & Allainé (2008) Ecological Modelling, 213 : 257-262.
- Girard, I., J. P. Ouellet, R. Courtois, C. Dussault, and L. Breton. 2002. Effects of sampling effort based on GPS telemetry on home-range size estimations. Journal of Wildlife Management 66:1290-1300.