Photophosphorylation
La photophosphorylation est l'ensemble des processus permettant la production d'ATP, la plupart du temps couplée à la production de NADPH par oxydation de l'eau, à partir de l'énergie lumineuse au cours de la photosynthèse. Elle regroupe les réactions directement dépendantes de la lumière et correspond à la « phase claire », ou « phase lumineuse », de la photosynthèse. Elle se déroule, chez les procaryotes et les cyanobactéries, dans la membrane des thylakoïdes, à l'intérieur des chloroplastes. Elle met en œuvre une chaîne de transport d'électrons à travers laquelle circulent les électrons excités par la lumière captée par la chlorophylle. Cette chaîne de transport d'électrons convertit l'énergie contenue dans ces électrons excités en gradient électrochimique à partir duquel une ATP synthase assure la phosphorylation de l'ADP en ATP. La photophosphorylation est en quelque sorte le pendant, pour la photosynthèse, de la phosphorylation oxydative pour la respiration cellulaire.
Dans les chloroplastes
Chez les plantes, la photophosphorylation se déroule dans la membrane des thylakoïdes, à l'intérieur des chloroplastes. Elle peut être non cyclique ou cyclique :
non cyclique H2O → photosystème II (P680 (en)) → plastoquinone → complexe cytochrome b6f → plastocyanine → photosystème I (P700 (en)) → ferrédoxine → ferrédoxine-NADP+ réductase → NADP+ ; cyclique photophosphorylation cyclique : (ferrédoxine →) plastoquinone → complexe cytochrome b6f → plastocyanine → photosystème I (P700 (en)) → ferrédoxine (→ plastoquinone).
Photophosphorylation non cyclique
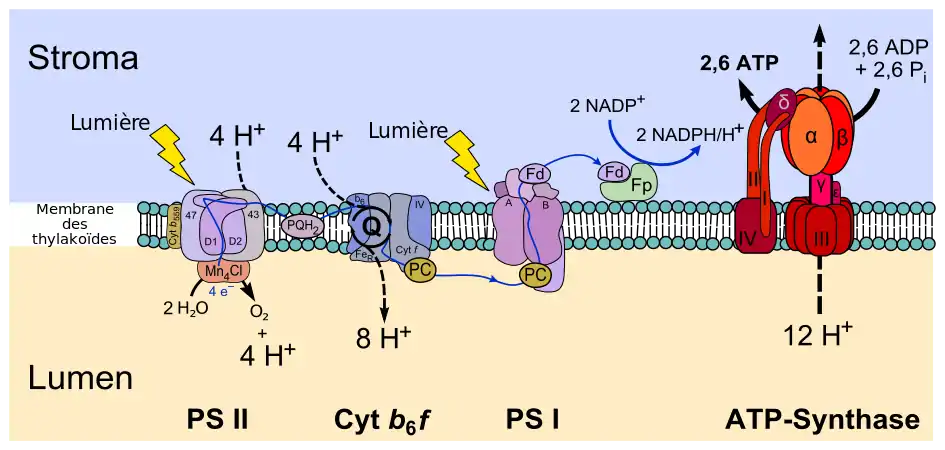
La photophosphorylation non cyclique consiste en un flux d'électrons depuis une molécule d'eau vers une molécule de NADP+ à travers une série d'enzymes membranaires et de protéines transporteuses avec génération d'un gradient de concentration en protons à travers la membrane des thylakoïdes au niveau du complexe cytochrome b6f. Ce gradient de concentration de protons génère un gradient électrochimique utilisé par l'ATP synthase pour réaliser la phosphorylation de l'ADP en ATP, ce qu'on appelle un couplage chimiosmotique. Ainsi, au cours de la photophosphorylation non cyclique, l'eau H2O est oxydée en oxygène O2 au niveau du photosystème II — plus précisément, du complexe d'oxydation de l'eau — et le NADP+ est réduit en NADPH par la ferrédoxine-NADP+ réductase.
Plus précisément, l'absorption d'un photon par une molécule de chlorophylle P680 du photosystème II conduit à l'excitation d'un électron, qui acquiert suffisamment d'énergie pour être cédé à un accepteur d'électrons par un phénomène de séparation de charges photoinduite. L'accepteur d'électrons primaire est une molécule de chlorophylle dépourvue d'atome de magnésium central appelée phéophytine. De là, l'électron excité passe sur une plastoquinone puis à travers le complexe cytochrome b6f avant d'être transporté sur une plastocyanine jusqu'à un photosystème I. Celui-ci contient un dimère de chlorophylle P700 capable d'exciter un électron par absorption d'un photon, électron transmis par la suite à une ferrédoxine, qui le cède à une ferrédoxine-NADP+ réductase pour réduire une molécule de NADP+ en NADPH.
Complexe d'oxydation de l'eau

L'eau H2O est la source d'électrons de la photosynthèse chez les plantes et les cyanobactéries. Deux molécules d'eau sont oxydées par quatre réactions successives de séparation de charges dans le photosystème II pour donner une molécule de dioxygène O2, quatre protons H+ et quatre électrons e−. Chacun des quatre électrons transite par un résidu de tyrosine jusqu'au P680 qui sert de donneur d'électrons primaire du centre réactionnel du photosystème II.
L'oxydation de l'eau est catalysée par un cluster oxométallique comprenant quatre cations de manganèse et un cation de calcium. Cette structure, appelée complexe d'oxydation de l'eau (Oxygen-Evolving Complex), se lie à deux molécules d'eau et permet les quatre oxydations successives aboutissant à la libération d'une molécule d'O2 et de quatre H+, ces derniers contribuant à l'établissement du gradient électrochimique à travers la membrane des thylakoïdes. Le photosystème II est la seule enzyme connue capable d'oxyder l'eau de cette façon. L'oxygène libéré est en fait un déchet, utilisé cependant par la majorité des êtres vivants pour leur respiration cellulaire, y compris les organismes photosynthétiques[2].
Schéma en Z
Le « schéma en Z » représente les variations d'énergie des électrons au cours de la photophosphorylation non cyclique :

On peut le tracer précisément à partir des valeurs du potentiel d'oxydoréduction E°' des différents couples rédox[3] :
| Couple rédox | Potentiel d'oxydoréduction (V) |
|---|---|
| H2O / O2 | + 0,82 |
| P680 / P680+ | + 0,9 |
| P680* / P680 | - 0,8 |
| Phéophytine réduite / oxydée | - 0,6 |
| Plastoquinone réduite / oxydée | 0 |
| Complexe cytochrome b6f | - 0,2 et + 0,2 |
| P700 / P700+ | + 0,4 |
| P700* / P700 | -1,3 |
| Ferrédoxine réduite / oxydée | - 0,42 |
| NADPH / NADP+ | - 0,32 |
Photophosphorylation cyclique
La photophosphorylation cyclique ne fait intervenir que le photosystème I, le complexe cytochrome b6f et la plastocyanine ; le photosystème I absorbe l'énergie lumineuse, ce qui permet d'exciter un électron, transféré à un complexe cytochrome b6f auquel il cède son énergie en générant un gradient électrochimique utilisé par une ATP synthase pour produire de l'ATP, avant de revenir au photosystème I via une plastocyanine. Ces réactions ne produisent ni O2 ni NADPH.
Chez les bactéries
Les bactéries sont des procaryotes et ne possèdent donc pas d'organites semblables aux chloroplastes des procaryotes. Les bactéries photosynthétiques — cyanobactéries, bactéries vertes sulfureuses (Chlorobi), bactéries vertes non sulfureuses (Chloroflexi), bactéries pourpres — présentent en revanche des ressemblances frappantes avec les chloroplastes eux-mêmes, ce qui est à l'origine de la théorie endosymbiotique.
Cyanobactéries
Les cyanobactéries sont les seules bactéries capables de photosynthèse oxygénique. À l'instar des chloroplastes, elles possèdent des thylakoïdes, dont la chaîne de transport d'électrons est essentiellement la même que celle des plantes — le cytochrome b6 est apparenté au complexe cytochrome b6f — à la différence près qu'elles utilisent le cytochrome c6 à la place de la plastocyanine :
non cyclique H2O → photosystème II (P680 (en)) → plastoquinone → cytochrome b6 → cytochrome c6 → photosystème I (P700 (en)) → ferrédoxine → NADP+ ; cyclique (ferrédoxine →) plastoquinone → cytochrome b6 → cytochrome c6 → photosystème I (P700 (en)) → ferrédoxine (→ plastoquinone).
Elles utilisent en revanche des pigments différents dans leurs antennes collectrices pour capter l'énergie électromagnétique de la lumière incidente, préférant les phycobilines aux chlorophylles. Elles sont également capables d'utiliser leur chaîne de transport d'électrons comme chaîne respiratoire et générer de l'ATP par oxydation du NADH avec conversion concomitante d'une molécule d'oxygène O2 en deux molécules d'eau H2O :
- NADH → NADH déshydrogénase → plastoquinone → cytochrome b6 → cytochrome c6 → cytochrome aa3 → O2.
Cette chaîne présente des analogies avec la chaîne respiratoire mitochondriale dans la mesure où on y retrouve une NADH déshydrogénase apparentée au complexe I, le couple plastoquinone / plastoquinol apparenté au couple ubiquinone / ubiquinol, le cytochrome b6 apparenté à la coenzyme Q-cytochrome c réductase (complexe III), le cytochrome c6 qui est un cytochrome c et le cytochrome aa3 apparenté à la cytochrome c oxydase (complexe IV).
Bactéries vertes sulfureuses
Les bactéries vertes sulfureuses n'utilisent qu'un photosystème, semblable au photosystème I des plantes :
non cyclique H2S → P840 → ferrédoxine → NAD+ ; cyclique (ferrédoxine →) ménaquinone → cytochrome bc1 → cytochrome c553 → P840 → ferrédoxine (→ ménaquinone).
La photophosphorylation cyclique de ces bactéries présente des analogies avec la chaîne respiratoire mitochondriale dans la mesure où le couple ménaquinone / ménaquinol est apparenté au couple ubiquinone / ubiquinol, le cytochrome bc1 est apparenté à (mais distinct de) la coenzyme Q-cytochrome c réductase (complexe III) et le cytochrome c553 est un cytochrome c.
Bactéries pourpres
Les bactéries pourpres utilisent un photosystème unique apparenté au photosystème II des cyanobactéries et des chloroplastes :
- photophosphorylation cyclique : (cytochrome c2 →) P870 → ubiquinone → cytochrome bc1 (→ cytochrome c2).
Il s'agit d'un processus cyclique entre un centre réactionnel P870 de bactériochlorophylle et une pompe à protons au niveau du cytochrome bc1, semblable à la coenzyme Q-cytochrome c réductase (complexe III) de la chaîne respiratoire mitochondriale mais néanmoins distincte de celle-ci.
Le NADPH est produit par une chaîne de transport d'électrons fonctionnant à partir de donneurs d'électrons tels que l'hydrogène H2, le sulfure d'hydrogène H2S, l'anion sulfure S2−, l'ion sulfite SO32−, ou des composés organiques tels que le succinate −OOC–CH2–CH2–COO− et le lactate H3C–CHOH–COO−.
Notes et références
- (en) Bessel Kok, Bliss Forbush et Marion McGloin, « Cooperation of charges in photosynthetic O2 evolution – I. A linear four step mechanism », Photochemistry and Photobiology, vol. 11, no 6, , p. 457-475 (PMID 5456273, DOI 10.1111/j.1751-1097.1970.tb06017.x, lire en ligne)
- (en) Yulia Pushkar, Junko Yano, Kenneth Sauer, Alain Boussac et Vittal K. Yachandra, « Structural changes in the Mn4Ca cluster and the mechanism of photosynthetic water splitting », Proceedings of the National Academy of Sciences of the United States of America, vol. 105, no 6, , p. 1879-1884 (PMID 18250316, PMCID 2542863, DOI 10.1073/pnas.0707092105, Bibcode 2008PNAS..105.1879P, lire en ligne)
- 12 - Le Schéma en Z, sur le site de l'UFR de biologie de l'université Pierre-et-Marie-Curie à Paris, France.