Théorie endosymbiotique
La théorie endosymbiotique, ou hypothèse de l'endosymbiose, est l'hypothèse selon laquelle les deux organites intervenants dans la gestion de l'énergie (chloroplastes et mitochondries) des cellules eucaryotes proviennent de l’incorporation (endocytose) par certaines archées, des bactéries avec lesquelles elles auraient entretenu une relation endosymbiotique. Elle s'oppose à la théorie autogénique (ou hypothèse autogène)[1], qui postule que le système endomembranaire (chloroplaste, mitochondrie et noyau), ainsi que le cytosquelette des cellules eucaryotes, aient évolué de novo par invagination de la membrane plasmique et complexification à partir d'une cellule procaryote ancestrale[2].
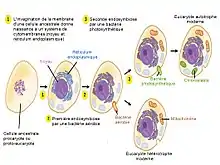
Les mitochondries et les chloroplastes sont des organites semi-autonomes de la cellule eucaryote, c'est-à-dire disposant d'un patrimoine génétique et capables de se diviser indépendamment de la cellule. Ces caractéristiques furent les premiers éléments à l'appui de la théorie endosymbiotique formulée par Antoine Béchamp (1869), Andreas Schimper[3] (1883) et Constantin Merejkovski (1905), et remise à l'honneur par Lynn Margulis dans les années 1960[4]. La théorie souligne que les types de bactéries incorporées auraient auparavant développé des spécialisations opérationnelles telles que la capacité de stocker de l'énergie sous forme biochimique (mitochondries) ou de capter l'énergie lumineuse (chloroplastes). Ces facultés sont en quelque sorte partagées, transmises puis héritées d'abord via la symbiose, puis en une synthèse résultant en un organisme plus complexe.
Selon Lynn Margulis, les processus résultant en l'apparition de nouvelles formes de vie à partir de symbioses, ou plus généralement synergies, entre formes de vie existantes, sont un moteur primordial de l'évolution, en particulier de sa complexification, et à l'origine de l'émergence des cellules de tous les organismes multicellulaires, dont l'espèce humaine.
Cette pluralité de l'individu a débouché sur la notion de l'holobionte ou du supraorganisme. Chaque organisme multicellulaire (plante, animal …) est une communauté symbiotique par son origine, mais aussi une chimère selon la théorie hologénomique de l'évolution (en). Cette endosymbiose s'apparente en effet à un processus de domestication favorisé par les transferts horizontaux de gènes d'origine évolutive différente (gènes archéens et bactériens) vers le génome nucléaire des eucaryotes. Ces gènes de procaryotes sont liés au métabolisme de l'ATP, aux ARNr, aux cytochromes et phytochromes, aux protéines qui synthétisent la cellulose de la paroi cellulaire[5] - [6].
Endosymbioses successives
Déroulement de l'endosymbiose en série : Une cellule procaryote ancestrale (hétérotrophe, anaérobie) va grossir ; il va y avoir une invagination de la membrane plasmique qui formera une ébauche de réticulum endoplasmique et de noyau. Cette cellule ancestrale va phagocyter une cellule procaryote et hétérotrophe mais qui est cette fois ci aérobie : c'est la première endosymbiose, elle explique l'apparition des mitochondries. La cellule nouvellement formée est un eucaryote ancestral (hétérotrophe et aérobie) ; c'est la cellule qui a été phagocytée qui est à l'origine de la mitochondrie de cet eucaryote[7].
Cette nouvelle cellule va à son tour phagocyter une cellule procaryote photosynthétique (donc autotrophe) et anaérobie (par exemple une cyanobactérie) : c'est la deuxième endosymbiose, elle explique l'apparition des chloroplastes. La cellule formée est un eucaryote photosynthétique (autotrophe) et aérobie[7].
On va pouvoir assister à un accroissement de la biodiversité.
Parasitisme initial ?
.svg.png.webp)
Les phylogénies des gènes composant les génomes des mitochondries ou celles issues des gènes nucléaires d'origine mitochondriale (beaucoup de ces gènes sont transférés au noyau lors du processus d'endosymbiose) montrent que ces mitochondries dérivent de bactéries appartenant au phylum des Alphaproteobacteria, plus précisément de l'ordre des rickettsiales[8]. De nombreuses espèces de ce groupe de bactéries sont des parasites intracellulaires obligatoires pénétrant dans la cellule par endocytose. Une hypothèse controversée (car elle va à l'encontre du principe de parcimonie[9]) suggère que les bactéries entrées dans la cellule possédaient probablement un flagelle dont elles ont perdu toute trace mais qui aurait facilité la pénétration du parasite. Les mitochondries seraient issues d'un parasitisme initial s'étant progressivement transformé en relation symbiotique[10].
Preuves appuyant la théorie

Cette théorie est aujourd'hui largement acceptée, de nombreux faits la corroborant ayant été découverts depuis sa formulation :
- Les mitochondries et les chloroplastes contiennent de l'ADN, lequel est assez radicalement différent de l'ADN du noyau cellulaire. Ils sont entourés de deux membranes au minimum, et la membrane interne montre des différences de composition importantes avec les autres membranes de la cellule. Elle est en fait plus proche d'une membrane de type bactérienne. Ces informations sont concordantes avec l'hypothèse de l'origine cellulaire des organites.
- Les nouvelles mitochondries et chloroplastes se forment uniquement par division. Chez certaines algues, les chloroplastes peuvent être détruits par certaines molécules ou l'absence prolongée de lumière sans aucun autre impact sur la cellule. Dans ce cas, les chloroplastes ne sont pas régénérés.
- La structure interne et la biochimie des chloroplastes (en particulier la présence des thylakoïdes et de certaines chlorophylles) sont très similaires à celles des cyanobactéries. Les études phylogénétiques suggèrent également que les génomes des chloroplastes sont très proches de ceux des cyanobactéries.
- Les analyses de séquences d'ADN et la phylogénie suggèrent que certains gènes de l'ADN nucléaire pourraient provenir des chloroplastes.
- Certains gènes du noyau sont transportés vers l'organite, et les organites (qu'il s'agisse des mitochondries ou des chloroplastes) ont des génomes inhabituellement petits comparés à ceux des autres organismes. Ceci renforce l'idée d'une dépendance accrue de la cellule hôte eucaryote après formation de l'endosymbiose.
- Plusieurs groupes de protistes présentent des chloroplastes, et ces groupes sont généralement plus souvent proches d'autres groupes de protistes sans chloroplastes, que de groupes de protistes avec chloroplastes. Ceci suggère que si les chloroplastes ont une origine cellulaire propre, ils sont apparus plusieurs fois en divers embranchements ; dans ce cas, leurs similitudes les uns par rapport aux autres seraient difficiles à expliquer.
- Le nombre de membranes des chloroplastes varie selon les groupes. Ceci suggère la possibilité d'endosymbiose en série. Chez certaines algues (Cryptophytes, Chromophytes) le chloroplaste dispose de 4 membranes. D'un point de vue évolutif, une cellule eucaryote disposant d'un chloroplaste (provenant d'une première endosymbiose) aurait été absorbée par une autre cellule eucaryote (seconde endosymbiose) : aux deux membranes plastidiales s'ajoutent ainsi la membrane limitant l'eucaryote phagocyté et la membrane de phagocytose[11].
- Le photosystème II est proche du photosystème des cyanobactéries, le photosystème I est proche du photosystème des bactéries vertes sulfureuses.
- Le rapport protéine/lipide des membranes des organites semi-autonomes et des bactéries Gram+ est similaire et différent de celui des autres membranes biologiques (rapport protéine/lipide de 3.0 pour les organites et bactéries contre en moyenne 1 pour les autres membranes).
Bibliographie
- General textbook: Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts and Peter Walter, Molecular Biology of the Cell, Garland Science, New York, 2002. (ISBN 0-8153-3218-1).
- Discusses theories on how mitochondria and chloroplast genes are transferred into the nucleus, and also what steps a gene needs to go through in order to complete this process: Jeffrey L. Blanchard and Michael Lynch (2000), Organellar genes: why do they end up in the nucleus? Trends in Genetics, 16 (7), pp. 315-320.
- Recounts evidence that chloroplast-encoded proteins affect transcription of nuclear genes, as opposed to the more well-documented cases of nuclear-encoded proteins that affect mitochondria or chloroplasts. Paul Jarvis (2001), Intracellular signalling: The chloroplast talks! Current Biology, 11 (8), pp. R307-R310.
Notes et références
- (en) F. J. R. Taylor, « Autogenous Theories for the Origin of Eukaryotes », Taxon, vol. 25, no 4, , p. 377-390 (DOI 10.2307/1220521).
- Thierry Lefevre, Michel Raymond et Frédéric Thomas, Biologie évolutive, De Boeck Superieur, (lire en ligne), p. 38.
- (de) A F. W. Schimper, 1883, « Über die Entwickelung der Chlorophyllkörner und Farbkörper », Bot. Z 41, 105–120, 121–136, 137–152, 153–162
- Édouard Boureau, La Terre, mère de la vie ?, Larousse, , p. 121.
- Marc-André Selosse, La symbiose. Structures et fonctions, rôle écologique et évolutif, Vuibert, , p. 121.
- (en) Ilana Zilber-Rosenberg, Eugene Rosenberg, « Role of microorganisms in the evolution of animals and plants: the hologenome theory of evolution », FEMS Microbiology Reviews, vol. 32, no 5, , p. 723–735 (DOI 10.1111/j.1574-6976.2008.00123.x).
- Marc-André Selosse et Susan Loiseaux-de Goër, « La saga de l'endosymbiose », La Recherche, no 296, , p. 36-41.
- (en) M. W. Gray & W. F. Doolittle, « Has the endosymbiot hypothesis been proven? », Microbiological Reviews, vol. 46, no 1, , p. 1–42
- Janlou Chaput, « La théorie de la mitochondrie parasite refait surface », sur futura-sciences.com, .
- (en) Davide Sassera et al., « Phylogenomic Evidence for the Presence of a Flagellum and cbb3 Oxidase in the Free-Living Mitochondrial Ancestor », Molecular Biology and Evolution, vol. 28, no 12, , p. 3285–3296 (DOI 10.1093/molbev/msr159)
- (en) Susan E. Douglas, « Eukaryote-eukaryote endosymbioses: insights from studies of a cryptomonad alga », Biosystems, vol. 28, nos 1-3, , p. 57-68 (DOI 10.1016/0303-2647(92)90008-M).