Cortex cérébral
Le cortex cérébral (ou écorce cérébrale[1]), d'origine prosencéphalique, est la substance grise périphérique des hémisphères cérébraux. Il se compose de trois couches (pour l'archi- et le paléocortex) à six couches (pour le néocortex) renfermant différentes classes de neurones, d'interneurones et de cellules gliales. Le cortex peut être segmenté en différentes aires selon des critères cytoarchitectoniques (nombre de couches, type de neurones), de leur connexions, notamment avec le thalamus, et de leur fonction. On pense que l'existence de ces circuits serait à l'origine de la pensée.
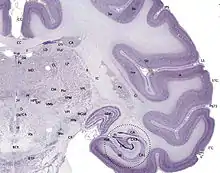
Le cervelet est lui enveloppé d'un cortex dit cérébelleux, à trois couches.
Antiquité et Moyen Âge
Pendant la haute Antiquité, le cerveau était considéré comme d'importance mineure dans le corps humain. Les Égyptiens ne se préoccupaient pas de conserver cet organe dans le processus de momification, néanmoins le papyrus Edwin Smith détaille des cas de traumatismes crâniens et médullaires et leurs conséquences. Le siège des pensées et des émotions était situé dans le cœur, croyance qui se retrouve aujourd'hui dans un certain nombre d'expressions ou de symboles. C'est au Ve siècle av. J.-C. avec les dissections d'Alcmaéon que le siège de la vision est localisé dans le cerveau. Par la suite Démocrite imagina l'âme sous forme de particules dispersées dans tout le corps, mais avec une forte concentration dans le cerveau. Aristote voyait essentiellement le cœur comme le siège de la pensée et des émotions. Le cerveau, quant à lui, servait uniquement de structure refroidissant le sang chauffé par le cœur puisqu'il avait cette tendance. Hippocrate apporta la vision selon laquelle le cerveau était le siège des sensations et des perceptions. Cette information eut l'influence dans les siècles suivants dans le domaine de l'apprentissage et de la neuroscience.
Le moment exact où le cortex fut connu des Antiques est indéterminé, mais Galien, grand médecin du IIe siècle, le connaissait. Il considérait cependant sous l'influence d'Hérophile que le siège de la pensée était localisé dans les ventricules cérébraux, les substances blanche et grise n'étant qu'un feutrage destiné à protéger ces dernières. Cette idée perdura pendant tout le Moyen Âge. Peu à peu, l'idée se fit que le cortex cérébral était à l'origine des fonctions cognitives complexes des Mammifères.
Renaissance et époque moderne
Les connaissances sur le cortex ont très peu évolué jusqu'à la Renaissance par manque d'un outil d'exploration efficace.
Au XVIIe siècle le microscope optique apparaît, puis devient pleinement fonctionnel moins d'un siècle plus tard. À cette époque, les moyens techniques étaient limités, mais les connaissances l'étaient encore plus. Il suffisait de pointer son microscope n'importe où pour faire des découvertes exceptionnelles. C'est ce que fit Malpighi dans les années 1660[2]. Il décrivit la première cellule du cortex. Mais les progrès restaient lents, car les cellules cérébrales sont très denses et difficiles à différencier par les colorations traditionnelles. Malgré ces difficultés, le Russe Vladimir Alexeïevitch Betz parvint en 1874 à identifier les cellules pyramidales[3]. C'est de Camillo Golgi que vint la solution, en 1873, quand il mit au point une coloration aux sels argentiques (la reazione nera) qui ne marquait que quelques cellules dans l'ensemble du tissu. Celles-ci apparaissaient donc pleinement différenciées de leurs voisines, avec toute leur arborescence bien visible. Toutefois, ce n'est pas lui, mais un contemporain, Santiago Ramón y Cajal qui se lancera dans l'exploration physiologique du cortex. Utilisant la technique de son confrère, il décrira les types cellulaires et l'organisation en six couches du néocortex. Les deux savants s'opposèrent longtemps, Golgi soutenant la théorie réticulaire (les neurones constituent un syncytium) du système nerveux, alors que Ramón y Cajal était un partisan de la théorie neuronale (les neurones sont des cellules indépendantes connectées entre elles par des synapses). Finalement, Golgi adopta les idées de Ramon y Cajal vers 1900[4] et ils reçurent conjointement le prix Nobel en 1906 pour leurs travaux histologiques sur le système nerveux. La théorie neuronale fut finalement confirmée au XXe siècle grâce au microscope électronique.
Parallèlement aux études histologiques furent menées des analyses fonctionnelles. À l'époque, la seule méthode était l'étude des conséquences d'une lésion du cortex cérébral sur les performances cognitives humaines. Les Égyptiens avaient déjà constaté que des traumatismes crâniens pouvaient entraîner des problèmes visuels. C'est Paul Broca qui, étudiant un patient aphasique, mettra en évidence le lien entre une lésion corticale et un déficit cognitif. Par la suite, la fonction de nombreuses zones cérébrales fut identifiée et à l'aube du XXe siècle, la localisation des aires visuelles, auditives, somatosensorielles et motrices était connue. À côté des lésions naturelles, les neurobiologistes étudièrent les effets des lésions provoquées, généralement dans un but prétendument thérapeutique, dont la principale et la plus connue n'est autre que la lobotomie. Une grande vague de lobotomie, évoquée dans le film Vol au-dessus d'un nid de coucou, qui déclinera à partir de 1950 (date de son interdiction en URSS) permit une meilleure connaissance du système nerveux, mais à un coût humain désastreux. Cette technique est aujourd'hui interdite en France, mais continue à être appliquée aux États-Unis, en Europe du Nord, en Inde et dans plusieurs autres pays.
Exploration fonctionnelle par imagerie cérébrale
En 1875, un médecin anglais, Richard Caton, est le premier à mesurer l'activité neuroélectrique du cortex cérébral en plaçant l'électrode d'un galvanomètre directement en contact avec la surface du cerveau d'animaux craniotomisés. Il montre ainsi que l'activité fonctionnelle (par exemple la vision) correspond à l'apparition d'une polarisation négative dans une zone circonscrite du cortex cérébral.
L'enregistrement de l'activité neuroélectrique chez l'être humain débutera avec l'apparition de l'électroencéphalographie (EEG) mise au point par Hans Berger au cours des années 1920. Cette technique permet pour la première fois d'étudier les corrélats neurophysiologiques des activités cognitives en temps réel avec une excellente résolution temporelle de l'ordre de la milliseconde. Cet examen reste incontournable pour le diagnostic et la classification des épilepsies.

C'est dans la seconde moitié du XXe siècle que se situe une grande révolution dans l'étude du cortex cérébral, avec la mise au point de méthodes d'imagerie cérébrale non invasives. Le médecin peut voir le cerveau fonctionner sans ouvrir le crâne. Jusque-là, les radiographies standards ne donnaient que des images inexploitables (le cerveau n'étant pas radio-opaque) et l'angiographie cérébrale ne permettait de voir que les axes vasculaires cérébraux. « Grâce à l’introduction des techniques de neuroimagerie, le niveau molaire de description a pu se substituer au niveau moléculaire dominant »[5].
Ce fut d'abord le scanner qui permit de visualiser, pour la première fois le cerveau et les zones corticales avec une précision remarquable, puis la résonance magnétique nucléaire (IRM) a considérablement modifié, à son tour, l'étude iconographique des structures corticales.
En 1938, le principe de résonance magnétique nucléaire est découvert par Isidor Isaac Rabi. Cette découverte aboutira, en 1973, à la mise au point de ce qui allait devenir l'imagerie par résonance magnétique ou IRM, simultanément par Paul Lauterbur et Peter Mansfield qui reçurent conjointement un prix Nobel (Prix Nobel de physiologie ou médecine) en 2003.
Évolution et développement du cortex
Évolution phylogénique
D'après la théorie de l'évolution, le cortex cérébral a subi une longue évolution depuis son apparition chez les premiers Crâniates ou leurs ancêtres[6]. On distingue chez les Mammifères le néocortex, aussi appelé néopallium et l'allocortex lui-même subdivisé en paléocortex (ou paléopallium) et archicortex (ou archipallium). L'archicortex est le plus ancien, il existe déjà chez les poissons dans le rhinencéphale, structure responsable du traitement des sensations olfactives. Chez l'être humain, on le trouve dans des structures très anciennes telles que l'hippocampe et le gyrus dentelé. Le paléocortex est plus récent. Il est bien représenté chez les reptiles où il atteint son développement maximum, mais il est encore présent chez les Mammifères dans des structures anciennes telles que les noyaux gris centraux ou le rhinencéphale. Le néocortex est phylogénétiquement le plus récent, bien qu'existant à l'état d'ébauche chez les reptiles. Il s'agît d'une couche superficielle lisse et peu développée chez les Oiseaux. Il va faire l'objet d'une croissance chez les Mammifères au point de repousser l'allocortex dans des zones réduites. C'est chez l'être humain qu'il atteint son développement maximum, constituant 80 % des neurones du système nerveux central.
Le cortex moderne des Mammifères dérive du rhinencéphale des poissons[7]. Le fait que l'ancienne structure olfactive des poissons ait donné naissance aux hémisphères cérébraux reflète l'importance qu'avait l'olfaction chez les premiers Mammifères et qu'elle possède encore aujourd'hui pour beaucoup d'entre eux. Une hypothèse informelle suggère que cela pourrait être dû à l'absence d'informations portées par les sensations olfactives. Alors que les signaux auditifs peuvent par eux-mêmes fournir des informations sur la taille, la position et le mouvement des autres animaux, ce n'est pas le cas des signaux olfactifs. Pour être utiles, ils doivent être associés à des traces mnésiques qui les mettent en relation avec des souvenirs visuels ou auditifs, d'autant que les premiers Mammifères étaient vraisemblablement des animaux nocturnes de la taille d'une souris. Les hémisphères cérébraux se sont donc développées et ont reçu des efférences visuelles, auditives et somatiques en vue d'intégrer toutes ces informations avec les signaux olfactifs. Ceci n'est qu'une hypothèse et il n'existe aucun moyen de la vérifier, l'évolution permettant de constater la succession des étapes, mais pas d'en donner les raisons, surtout pour un organe fossilisant aussi mal que le cerveau.
Développement embryologique (évolution ontogénique)
Le cortex cérébral se forme dans la partie antérieure du tube neural, lui-même dérivant de la plaque neurale, une différenciation de l'ectoderme dorsal sous l'influence de la notochorde.
La première structure cérébrale à se différencier dans ce qui donnera les hémisphères cérébraux est le système ventriculaire. Les cellules souches des neurones se situent dans l'épithélium qui borde les ventricules. Dans un premier temps, les progéniteurs se divisent de façon symétrique pour se multiplier puis de façon asymétrique. Une des deux cellules migre alors à l'extérieur de la zone ventriculaire pour atteindre le cortex[8]. Elle se différencie alors en neurone[9]. L'autre cellule reste dans la zone ventriculaire et continue à se diviser. Les cellules gliales se multiplient de façon similaire, leurs progéniteurs sont différents de ceux des neurones.
Pendant les périodes fœtale et néonatale, les neurones du cortex cérébral immature (la plaque corticale) sont pris en sandwich entre la zone marginale à l'extérieur et la sous-plaque située juste en dessous à l'interface avec ce qui donnera la substance blanche. La sous-plaque a une existence transitoire. Elle disparaitra chez l'être humain, deux mois après la naissance. La zone marginale persistera, devenant la couche 1 du néocortex.
Structure
Histologie
L'étude histologique du cortex a commencé très tôt. Les types cellulaires ont été mis en évidence dès le XIXe siècle. De grands noms ont été associés à ces découvertes, tels que les prix Nobel Santiago Ramon y Cajal et Camillo Golgi.
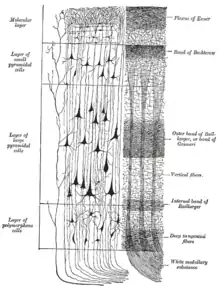
Dans cette image, les couches II et III ainsi que les couches IV et V ont été regroupées.
Au microscope, le néocortex humain apparaît subdivisé en six couches[10]. Ce nombre varie selon les espèces, par exemple cinq chez le dauphin, trois chez les reptiles.
Les couches sont numérotées depuis la surface. On distingue dans l'ordre :
- La couche moléculaire. Elle contient des axones et des dendrites. Les neurones des couches internes y envoient des dendrites courts et orientés perpendiculairement à la surface du cortex et des axones longs orienté parallèlement à cette surface[11]. Les prolongements neuronaux y présentent une structure similaire à celle trouvée dans le cortex cérébelleux, rappelant les mémoires toriques des ordinateurs des années 1950. On y trouve aussi quelques neurones de Cajal-Retzius et des neurones étoilés ;
- La couche granulaire externe contient des neurones granulaires. Elle reçoit les afférences d'autres aires du cortex. On parle en ce qui la concerne de connexions cortico-corticales afférentes ;
- La couche pyramidale externe. Constituée de cellules pyramidales, elle émet des connexions vers d'autres zones du cortex cérébral. Il s'agit ici de connexions cortico-corticales efférentes ;
- La couche granulaire interne. Elle contient des neurones étoilés et pyramidaux. C'est par cette couche que les informations en provenance de l'extérieur du cortex (par exemple du thalamus) entrent dans le cortex. Elle reçoit aussi les afférences en provenance de l'autre hémisphère cérébral ;
- La couche pyramidale interne. C'est également une couche envoyant des connexions efférentes mais qui sortent du cortex. C'est, par exemple, de cette couche que partent les neurones qui innervent les motoneurones ;
- La couche polymorphe, la couche la plus interne à l'état adulte. Elle envoie des prolongements axonaux en direction du thalamus permettant une rétroaction sur les entrées du cortex cérébral.
Une septième couche existe transitoirement pendant l'embryogenèse. Elle disparait avec la maturation cérébrale.
Ces couches corticales ne sont pas un simple empilement de neurones. Les neurones s'organisent en unités fonctionnelles prenant la forme de colonnes perpendiculaires à la surface du cortex, chacune assurant une fonction précise. Cependant il n'est pas possible de les distinguer par des méthodes histologiques ; ce sont par des études fonctionnelles du cortex visuel que cette structure a été mise en évidence avant d'être généralisée à l'ensemble du néocortex[12]. Le paléocortex et l'archicortex, structurellement plus simples, ne sont pas stratifiés et ne présentent pas cette structure en colonnes.
Anatomie
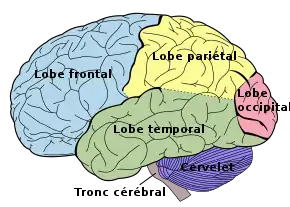
Chez l'être humain, l'épaisseur du cortex est comprise entre 1 et 4,5 millimètres[13] et sa surface avoisine 2 600 centimètres carrés (0,26 m2)[12], soit une surface dix fois plus importante que le singe[14]. Il contient environ 16[15] des 86 (voire 100[16]) milliards de neurones qui composent le cerveau humain.
Afin de pouvoir loger dans la boîte crânienne, le cortex est plissé par des sillons (sulcus en latin scientifique) ou scissures, de profondeur variable, délimitant des crêtes appelées gyrus[17] ou circonvolutions cérébrales.
Ce genre de disposition est dit gyrencéphale, par opposition aux lissencéphales existant par exemple chez le rat, qui présente un cortex lisse, dépourvu de circonvolution. Cette propriété n'est pas en relation avec la complexité du cerveau, mais est liée à la taille de l'individu. Lorsque la taille d'un individu double, son volume est en gros multiplié par huit (8=23). Si le cortex restait lisse, sa surface ne serait multipliée que par quatre. Pour préserver la proportion, le cortex doit se plisser en fonction de l'augmentation de taille. À ceci, s'ajoutent des spécificités liées aux lignées : par exemple, à taille égale, les carnivores disposent en général d'un cortex plus grand que les herbivores..
Les scissures les plus profondes divisent le cortex en lobes. Selon leur situation, on parle de lobe frontal, pariétal, occipital, insulaire et temporal.
Sous le cortex se trouve la substance blanche composée d'axones qui établissent les connexions entre corps cellulaires du cortex et d'autres parties du cerveau.
Organisation
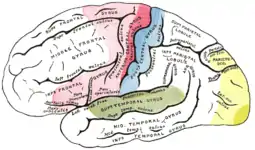
| Aire motrice primaire | |
| Aire motrice supplémentaire préfrontale | |
| Aire sensorielle primaire | |
| Aire d'association sensorielle | |
| Aire visuelle primaire | |
| Aire auditive primaire |
Le cortex cérébral est divisé en zones fonctionnelles, appelées aires, chacune assurant une fonction cognitive précise. Ce sont les études de Paul Broca en 1861 qui ont pour la première fois suggéré l'existence d'une telle organisation du néocortex[18]. Ces zones sont à peu près identiques pour tous les individus d'une même espèce, mais présentent de petites différences. Cette spécialisation est floue chez les mammifères primitifs et se précise au fur et à mesure que l'on se rapproche de l'être humain.
On distingue trois grands types de zones : les aires sensorielles, les aires motrices et les aires d'association.
Aires sensorielles chez l'humain
Trois aires cérébrales sont spécialisées dans le traitement des données sensorielles : le cortex auditif dans le lobe temporal, le cortex visuel situé dans le lobe occipital[19], et le cortex somatosensoriel dans le lobe pariétal.
Le cortex auditif est organisé de façon similaire au cortex visuel. Il est situé dans le lobe temporal. Il comprend une aire auditive primaire qui identifie les fréquences et une aire auditive secondaire qui reconstitue les sons.
Le cortex visuel est divisé en deux zones : le cortex visuel primaire qui est une projection directe de la rétine et effectue un traitement de bas niveau sur les données visuelles (identification des lignes, des couleurs, des sens de déplacements) et un cortex visuel secondaire qui rassemble ces éléments pour obtenir des objets ayant une forme, une couleur et un mouvement précis. Comme pour les informations motrices, les afférences de cette aire sont croisées, mais de façons différentes : l'hémisphère gauche ne reçoit pas les données en provenance de l'œil droit, mais en provenance de la partie droite du champ visuel de chaque œil.
Le cortex somatosensoriel est le reflet exact du cortex moteur primaire. Chaque organe y projette des afférences façon somatotopique. La taille de la zone affectée à chaque partie du corps est proportionnelle à la discrimination spatiale de la zone : main et visage y disposent donc des surfaces les plus importantes. Cette disposition est matérialisée par le concept d'homonculus sensitif[20]. En revanche, les différents types de sensation ne semblent pas séparés à ce stade.
Aires motrices chez l'être humain
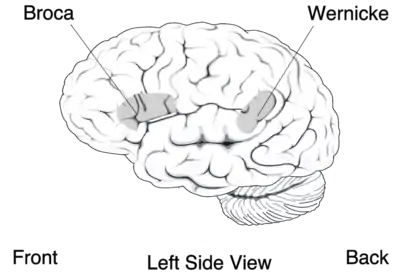
On distingue deux zones du cortex spécialisées dans la motricité, l'une et l'autre dans le cortex frontal.
La principale est l'aire motrice primaire qui occupe toute la partie postérieure du lobe frontal, juste en avant du sillon central. Elle est organisée de façon somatotopique (chaque zone du corps recevant une afférence d'une partie précise de cette aire), la surface associée à un muscle étant proportionnelle à la précision des mouvements dont il est capable : le visage et la main sont donc fortement représentés. Comme pour la somesthésie, il existe ici un homoncule moteur. On y trouve l'aire de Broca (cf. image de droite) dont l'atteinte est responsable de l'aphasie de Broca[21], une maladie dans laquelle une personne peut exprimer sa pensée sous forme de phrases cohérentes, mais ne peut pas les prononcer. Le rôle de l'aire motrice primaire est d'effectuer les mouvements volontaires. Les afférences des neurones de cette zone sont croisées : l'hémisphère gauche commande les mouvements de la partie droite du corps et inversement.
Il existe aussi une aire motrice supplémentaire, située dans le cortex préfrontal, et qui sélectionne les mouvements volontaires.
D'autres aires motrices ont été découvertes dans le cortex pariétal et préfrontal, participant à l'intégration spatiale du mouvement et aux relations entre mouvement et pensée.
Aires d'association chez l'être humain
Les aires d'association représentent la majeure partie du cortex cérébral chez l'être humain et sont le principal facteur de la taille de son cerveau. En fait, ce terme désigne toutes les aires néocorticales qui ne sont ni motrices, ni sensorielles ; leurs fonctions sont donc très diverses.
On en trouve dans trois lobes cérébraux :
- le lobe temporal dispose d'aires impliquées dans la mémorisation. Il contribue à des fonctions évoluées telles que le langage ou l'identification des visages ;
- le lobe pariétal contient la zone d'association sensorielle qui intègre les données en provenance de tous les systèmes sensoriels pour donner une image de l'ensemble de l'environnement. Il comporte aussi une petite zone impliquée dans le langage, l'aire de Wernicke, dont la lésion entraîne l'aphasie de Wernicke qui se différencie de l'aphasie de Broca par le fait que le malade peut former des phrases grammaticalement et syntaxiquement justes, mais sans signification ;
- le lobe préfrontal est celui qui a subi le plus fort développement dans la lignée humaine. C'est ici que l'on situe le siège de l'intelligence humaine. Le fait est qu'il est quasiment absent chez la plupart des mammifères alors qu'il constitue presque un quart de la surface du cortex chez l'être humain. Il reçoit des afférences de toutes les zones du cerveau et assure leur intégration pour matérialiser la pensée et aboutir à une prise de décision.
Autres zones
La zone de cortex du sillon médian, entre les deux hémisphères, est appelée le cortex cingulaire : c'est une zone de paléocortex appartenant au système limbique, un système impliqué dans la mémorisation et les émotions. Il est d'une grande importance dans les relations sociales.
Exemple de fonctionnement du cortex
Pour analyser une scène et fournir une réponse adaptée, toutes les aires cérébrales vont collaborer. Prenons l'exemple d'un chat qui miaule pour demander à manger.
L'aire visuelle primaire va identifier une série de lignes, de courbes et de taches de couleurs grises, rousses et blanches. L'aire visuelle secondaire va organiser ces éléments disparates en un objet immobile gris, roux et blanc. La zone d'intégration visuelle va reconnaître cet objet comme étant un chat assis. Parallèlement, l'aire auditive primaire va capter un certain nombre de fréquences ; l'aire auditive secondaire va organiser ces fréquences pour obtenir un son avec un timbre et une tessiture précises. L'aire d'intégration auditive va reconnaître un miaulement. L'aire d'association pariétale va identifier un chat qui miaule. Avec l'aide du lobe temporal, il va identifier le chat et la nature du miaulement. Tous les éléments du chat sont maintenant identifiés.
À partir de là, nous allons obtenir une réaction comportementale. Le système limbique situé, en partie entre les deux hémisphères, nous signale l'existence d'un attachement pour cet animal. Il va nous pousser à nous désintéresser de notre action en cours pour nous occuper en priorité de ses besoins. À noter que si l'individu agit parce que les miaulements du chat l'agacent, le résultat est le même, c'est le même système qui gère ces deux émotions. Le cortex préfrontal va prendre la décision de nourrir le chat (ou de le chasser selon les cas). L'aire motrice supplémentaire va organiser le déroulement de l'acte et l'aire motrice primaire va commander les mouvements nécessaires à son accomplissement. L'aire somatosensorielle primaire, en relation avec l'aire visuelle, va guider le déroulement des opérations en signalant à chaque instant les changements dans l'environnement (le chat a tendance à se mettre dans nos jambes) et la position des différents segments du corps dans l'espace.
Finalement, par la collaboration de toutes les aires cérébrales, un comportement coordonné a permis la résolution d'un problème. Signalons au passage que ce n'est pas la nature du cortex qui différencie l'être humain des autres animaux, mais la taille du cortex qui permet des réactions bien plus complexes que celles décrites ici. Notamment, les fonctions cognitives les plus évoluées telles que le langage ou la pensée symbolique ne sont pas mises en jeu ici. À l'inverse, le cortex ne fait pas tout. Par exemple, le mouvement de rotation de la tête pour regarder le chat n'est pas sous le contrôle du cortex, mais des tubercules quadrijumeaux, structure ancienne du mésencéphale, qui, chez les reptiles, avaient la fonction dévolue aujourd'hui aux aires visuelles et auditives corticales chez les mammifères.
Glutamate
Le glutamate est l'un des neurotransmetteurs les plus utilisés dans le cerveau puisque plus du tiers des neurones y fait appel. Les neurones l'utilisent tel quel ou par l'un de ses métabolites, le GABA. Côté postsynaptique, il existe trois types de récepteurs : le récepteur AMPA, le récepteur NMDA et le récepteur kaïnate, ainsi nommés en raison des molécules pharmacologiques capables de les activer sélectivement en l'absence de glutamate. Les deux premiers sont impliqués dans les phénomènes de mémorisation ; le rôle du troisième est moins bien compris.
Ces récepteurs sont des canaux ioniques : sodiques en ce qui concerne les récepteurs AMPA et kaïnate, calciques pour le NMDA. Leurs effets sont tous excitateurs sur les éléments postsynaptiques, ce qui signifie qu'ils vont favoriser l'émission d'un potentiel d'action par le neurone cible.
Ces récepteurs sont la cible de certaines drogues qui vont les activer en continu, ce qui va provoquer des hallucinations, et pour le NMDA, de par la cytotoxicité du calcium, une mort du neurone par apoptose.
GABA
L'acide gamma-amino butyrique (GABA) est le principal neurotransmetteur inhibiteur du système nerveux central. C'est un neuromodulateur qui est reconnu comme étant inhibiteur à l'âge adulte, mais qui est excitateur lors du développement embryonnaire. Il possède par ailleurs un rôle neurotrophique, c'est-à-dire qu'il favorise la croissance de certains neurones.
Acétylcholine
L'acétylcholine est l'un des premiers neurotransmetteurs découverts[22]. Son fonctionnement a été surtout étudié au niveau de la plaque motrice, mais il est présent partout dans le système nerveux. Il existe deux récepteurs de l'acétylcholine, tous deux présents dans le cortex : le récepteur nicotinique, dont l'agoniste est la nicotine et le récepteur muscarinique sensible à la muscarine. D'autres drogues peuvent distinguer des sous-types au sein de ces deux grandes familles de récepteurs.
Le récepteur nicotinique est un récepteur canal qui laisse passer les ions sodium quand il est activé. En revanche, le récepteur muscarinique est un récepteur de type métabotropique, ce qui signifie qu'il ne va pas ouvrir un canal ionique, mais synthétiser une molécule qui va avoir un effet sur le fonctionnement du neurone. Ce genre de récepteur est plus impliqué dans les phénomènes de régulation à moyen et long terme que dans la transmission du potentiel d'action. On parle ici plutôt de neuromodulation.
Les neurones cholinergiques sont parmi les neurones les plus touchés dans la maladie d'Alzheimer et les premiers à avoir été mis en évidence dans cette pathologie.
Neuromédiateurs
Les neuromédiateurs sont des molécules similaires aux neurotransmetteurs (parfois une molécule peut avoir les deux rôles), mais qui sont émis non pas dans une fente synaptique, mais dans l'environnement cérébral. Ces molécules atteignent les neurones de façon non spécifique. Leur rôle n'est pas de propager un potentiel d'action au travers d'une synapse, mais de créer une ambiance moléculaire qui mettra le système nerveux dans un état précis.
Dans le cortex, on connaît plusieurs molécules de ce type :
- la noradrénaline qui a le rôle d'un système d'urgence, potentialisant l'attention, la mémorisation et le rappel ;
- la sérotonine ou l'hydroxy-tryptamine impliquée dans les cycles veille/sommeil, mais aussi le comportement alimentaire, sexuel et bien d'autres. Certains antidépresseurs et médicaments psychotropes agissent sur la recapture de la sérotonine et potentialisent donc son effet ;
- la dopamine impliquée dans le système de récompense ;
- la mélatonine impliquée dans la régulation du cycle circadien, le contrôle hormonal et jouant un rôle dans la dépression hivernale.
Tous ces neuromédiateurs sont sécrétés par des neurones dont le corps cellulaire est la plupart du temps dans le tronc cérébral. Cela permet à des structures phylogéniquement plus anciennes d'exercer un certain contrôle sur le cortex.
Pathologies
Malformations congénitales
La plupart des maladies qui affectent le cortex ne sont pas spécifiques de cet organe. C'est le cas de l'anencéphalie, l'hydrocéphalie, la macrocéphalie et des nombreuses autres malformations de l'encéphale ; d'autres pathologies telles que les tumeurs sont encore plus générale. Toutefois, une classe de maladies est vraiment spécifique du cortex, il s'agit de celles qui affectent le bon déroulement de la gyration, c'est-à-dire la formation des circonvolutions cérébrales au cours de l'embryogenèse. On en compte plusieurs (lissencéphalie, polymicrogyrie, pachygyrie), c'est-à-dire soit un déficit de gyration conduisant à un cortex insuffisamment plissé et donc trop petit, soit à l'inverse une gyration excessive conduisant à des sillons petits et nombreux ou grands mais peu nombreux.
Épilepsies
Les épilepsies sont des maladies du cortex variées dans leurs symptômes, mais ayant une seule origine : une activation explosive du cortex cérébral : selon la zone du cortex perturbé, les résultats en seront très différents, depuis l'attaque convulsive connue sous le nom de grand mal jusqu'à des hallucinations, des mouvements involontaires brutaux ou des absences temporaires de conscience. Cette maladie a une composante génétique, mais elle peut aussi résulter d'une infection ou d'un traumatisme. S'il n'existe aucun moyen de stopper une crise en cours, des traitements médicamenteux ou chirurgicaux peuvent prévenir la survenue de nouvelles attaques.
Destructions localisées
Les causes peuvent être multiples :
- tumeur intracrânienne (bénigne ou maligne) compressive ;
- traumatisme crânien : accident ou par arme (arme à feu, le plus souvent), avec hématome ou lésions tissulaires ;
- accident vasculaire cérébral ;
- altération par des produits neurotoxiques : alcool, ectasy, plomb, mercure…, ou un manque de sommeil (particulièrement paradoxal) prolongé.
Le siège des lésions et leur étendue détermineront l'importance et le type de troubles neurologiques.
Neurodégénérescences liées à l'âge
Parmi ces maladies, dont le diagnostic souvent tardif et peu discriminant rend le traitement difficile, il faut citer la maladie d'Alzheimer et la maladie de Parkinson.
Maladie d'Alzheimer
Cette maladie, d'abord caractérisée par ses symptômes (une diminution précoce des capacités cognitives du sujet), correspond à une dégénérescence du cortex cérébral. Le tissu nerveux se parsème progressivement de « plaques amyloïdes », agrégats formés d'une protéine issue des membranes cellulaires et incomplètement dégradée, qui provoque l'invalidité par compression des neurones environnants. Sa prévalence a une composante génétique, mais des facteurs environnementaux peuvent influer sur son apparition et son évolution.
Notes et références
- Informations lexicographiques et étymologiques de « Cortex » dans le Trésor de la langue française informatisé, sur le site du Centre national de ressources textuelles et lexicales, Étymologiquement le mot cortex signifie bien écorce, mais l'expression tombe en désuétude.
- (en) M. Malpighi, « An Account of Some Discoveries Concerning the Brain, and the Tongue, Made by Signior Malpighi, Professor of Physick in Sicily », Philosophical Transactions (1665-1678), vol. Volume 2 - 1666/1667, (DOI 10.1098/rstl.1666.0033, lire en ligne)
- (en) W. Betz, « Anatomischer Nachweis zweier Gehirnzentra », Zentralbl Med Wiss, vol. 12, , p. 578-580, 595-599
- (en) C. Golgi, The neuron doctrine : theory and facts, Nobel Lecture, (lire en ligne)
- Marc Jeannerod, « La création de l’Institut des sciences cognitives du CNRS (1992-1998) », La Revue pour l'histoire du CNRS, no 10, mai 2004. [lire en ligne].
- (en) Ann B. Butler, « Chordate evolution and the origin of craniates: An old brain in a new head », Anat. Rec., vol. 261, no 3, , p. 111-125 (DOI 10.1002/1097-0185(20000615)261:3<111::AID-AR6>3.0.CO;2-F, résumé, lire en ligne)
- Pierre Teilhard de Chardin, La place de l'Homme dans la nature, 1949, p.47. [lire en ligne] [PDF]
- (en) J. Angevine and R. Sidman, « Autoradiographic study of cell migration during histogenesis of cerebral cortex in the mouse », Nature, vol. 192, , p. 766-768 (résumé)
- (en) P. Rakic, « Specification of cerebral cortical areas », Science, vol. 241, no 4862, , p. 170-176 (résumé)
- Santiago Ramon y Cajal, « La fine structure des centres nerveux », Proc. R. Soc. Lond,
- (en) Jones EG and Powell TPS, « electron microscopy of the somatic sensory cortex of the cat : II the fine structure of layer I-II », Philos Trans R Sot Lond [Biol], vol. 257, , p. 13-21
- (en) V. Mountcastle, « The columnar organization of the neocortex », Brain, vol. 120, , p. 701-722 (DOI 10.1126/science.3291116, lire en ligne)
- B. Fischl et A. M. Dale (2000). Measuring the thickness of the human cerebral cortex from magnetic resonance images. Proc Natl Acad Sci U S A, 97(20):11050–5.
- Lucile Guittienne et Marlène Prost, Homme-femme : de quel sexe êtes-vous ?, Presses Universitaires de Nancy, , p. 65.
- source?
- Neurosciences 5ème édition, De Boeck
- le terme de latin anatomique gyrus a pour pluriel gyri, en terminologie française le pluriel est gyrus.
- R. Saban, Conception de la physiologie cérébrale de François-Joseph Gall à Paul Broca : les localisations des fonctions cérébrales, in Biométrie humaine et anthropologie - Tome 20, numéros 3-4, juillet - décembre 2002, p. 195-203. [lire en ligne], (Cf. marque 32)
- (en) JS Bolton, « The Exact Histological Localisation of the Visual Area of the Human Cerebral Cortex », Philosophical Transactions of the Royal Society of London, vol. 193, , p. 165-222Ces travaux ont été menés à partir de résultats médicaux antérieurs obtenus par d'autres auteurs.
- (en) W Penfield et T Rasmussen, The cerebral cortex of man, New York, Macmillan, c'est dans cet ouvrage que sont décrits pour la première fois les homoncules moteurs et sensoriels
- P Broca, « Remarques sur le siège de la faculté du langage articule, suivies d'une observation d'aphemie », Bulletin de la Société Anatomique de Paris,
- (en) HH Dale, W Feldberg and M Vogt, « Release of acetylcholine at voluntary motor nerve endings », The Journal of Physiology, vol. 86, no 4, , p. 353-380 (lire en ligne)
Voir aussi
Articles connexes
- Néocortex
- Lobes du cerveau
- Cortex sensoriel et plasticité cérébrale
Liens externes
- Ressources relatives à la santé :
- FMA
- TA2
- Uberon
- (en) Medical Subject Headings
- (en) NCI Thesaurus
- (la + en) TA98
- (cs + sk) WikiSkripta
- Notices dans des dictionnaires ou encyclopédies généralistes :
- Émission sur le cortex cérébral de BioTV accompagnée de quelques illustrations légendées sur le cortex.
- Fonctionnement du cerveau sur le site de l'université McGill (en plus d'explications sur le cortex cérébral, différents niveaux d'explications (débutant, intermédiaire, avancé) sont disponibles)
- (en) Marino L, Connor RC, Fordyce RE, Herman LM, Hof PR, et al., « Cetaceans Have Complex Brains for Complex Cognition », PloS Biology