Cortex insulaire
Le cortex insulaire ou insula — terme latin signifiant île — est une partie du cortex cérébral et constitue l'un des lobes du cerveau. Son rôle est encore mal connu mais il est généralement associé aux fonctions limbiques et interviendrait notamment dans le dégoût, la dépendance ou encore la conscience.

| Système | |
|---|---|
| Région |
Tête |
| Vascularisation |
| Nom latin |
Cortex insularis |
|---|---|
| MeSH |
D000087623 |
| TA98 |
A14.1.09.149, A12.2.07.053 |
| TA2 |
5502 |
| FMA |
67329 |
| Sujet | |
|---|---|
| Page |
823 à 827 |
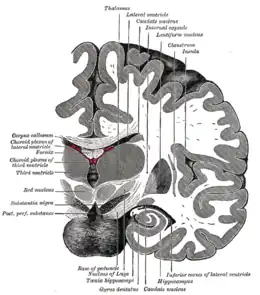
Le cortex insulaire est situé au fond du sillon latéral. Sa surface est marquée par cinq gyrus de l'insula : trois gyri courts en avant et deux autres plus longs en arrière. Il est encerclé par le sillon circulaire qui marque la séparation avec les parties operculaires des lobes temporaux, frontaux et pariétaux. L'ensemble du cortex qui le recouvre est appelé opercule et serait notamment impliqué dans la conscience.
Le cortex insulaire est divisé en deux parties : une large insula antérieure et une petite insula postérieure, dans laquelle ont été identifiées plus d'une douzaine d'aires différentes. La partie antérieure du lobe de l'insula, une composante des aires prémotrices, joue un rôle dans la production du langage et sa partie postérieure contient des aires somatosensorielles secondaires intégrant des informations en provenance de différentes modalités.
Le cortex insulaire joue un rôle dans diverses fonctions, principalement liées aux émotions ou à la régulation de l'homéostasie du corps. Ces fonctions incluent la perception, le contrôle moteur, la conscience du soi, le fonctionnement cognitif et les expériences interpersonnelles. Cette partie du cerveau est donc fortement impliquée dans certains dysfonctionnements psychopathologiques.
La dénomination latine du cortex insulaire est lobus insularis. Il est aussi désigné sous le nom insula de Reil (ou île de Reil) en référence au médecin allemand, Johann Christian Reil, qui en donna le premier une description anatomique en 1796[2].
Neuroanatomie
Connexions
L'insula antérieure reçoit une projection directe de la partie basale du noyau ventral médian du thalamus, ainsi que des afférences importantes provenant du noyau central de l'amygdale. De plus, l'insula antérieure, elle-même, projette vers l'amygdale.
Une étude chez le macaque rhésus a révélé d'importantes connexions réciproques (dans les deux sens) entre le cortex insulaire et presque tous les noyaux du complexe de l'amygdale. L'insula postérieure projette majoritairement vers la partie dorsale du noyau latéral et vers le noyau central de l'amygdale. À l'inverse, l'insula antérieure projette vers les aires antérieures de l'amygdale, ainsi que vers les noyaux médian, cortical, accessoire, basal magnocellulaire, médial-basal et latéral de l'amygdale[3].
L'insula postérieure est reliée réciproquement avec le cortex somatosensoriel secondaire (S2) et reçoit des afférences du noyau ventral postérieur inférieur du thalamus. Des travaux, plus récents[4] montrent que cette région reçoit également des informations du noyau ventromédian du thalamus (plus particulièrement, la partie postérieure de ce noyau) qui serait impliqué dans différentes fonctions émotionnelles ou homéostatiques, telles que la douleur, la régulation de la température, l'irritation, le niveau d'oxygénation local ou encore le sens du toucher.
Cyto-architecture
Le cortex insulaire est constitué d'une structure cellulaire (ou cytoarchitectonie) variable, passant de granulaire dans sa partie postérieure, à agranulaire dans sa partie antérieure.
Fonctions
Conscience intéroceptive
L'insula antérieure droite participe à la conscience intéroceptive du corps, notamment la capacité de mesurer son propre rythme cardiaque. De plus, il existe une corrélation entre le volume du cortex de cette zone et la précision avec laquelle on ressent l'intérieur de son corps[5]. Cette aire est également liée au contrôle de la pression sanguine, notamment pendant et après l'exercice[6]. Elle est ainsi plus activée lorsque le cerveau perçoit un effort important[7] - [8].
Le cortex insulaire est également impliqué dans l'évaluation de l'intensité d'une douleur[9]. C'est également cette zone qui est activée lorsque la douleur est imaginée, par exemple en regardant des images d’événements douloureux, et penser comme si elle nous était directement infligée[10]. Les individus souffrant du syndrome du colon irritable auraient un traitement anormal de la douleur viscérale au niveau du cortex insulaire, liée à une inhibition de la douleur irrégulière au niveau du cerveau[11].
Une autre fonction évaluative de l'insula antérieure droite est le degré de chaleur (non-douloureuse)[12] ou de froid (non-douloureux)[13] ressentis au niveau de la peau. De même, la sensation de tension au niveau de l'estomac ou de l'intestin est corrélée avec une activité de cette zone du cerveau[14] - [15]. Le cortex insulaire s'allume également lorsque la vessie est remplie ou stimulée[16].
Une étude d'imagerie cérébrale suggère également que la gêne respiratoire ressentie subjectivement lors de dyspnée est traitée au niveau de l'insula antérieure droite et de l'amygdale chez l'Homme[17].
Le cortex cérébral responsable du système vestibulaire se prolonge jusqu'à l'insula[18], de petites lésions du cortex insulaire antérieure étant capables de faire perdre le sens de l'équilibre ou de provoquer le vertige[19].
L'insula est également activée pour d'autres perceptions non-intéroceptives, comme l'écoute passive de musique[20], le rire ou les pleurs[21], l'empathie et la compassion[22], et enfin le langage[23].
Contrôle moteur
Le cortex insulaire contribue au contrôle du mouvement des mains et des yeux[24] - [25], de la déglutition[26], de la motilité gastrique[27], ou encore de l'articulation du langage[28] - [29]. Il a également été identifié comme le « centre de commande » chargé de contrôler l'augmentation du rythme cardiaque et de la pression sanguine au début de l'exercice physique[30]. Certaines recherches sur la conversation lui attribuent un rôle dans la capacité de prononcer des phrases longues et complexes[31]. Il est également impliqué dans certaines formes d'apprentissage moteur[32].
Homéostasie
Dans l'homéostasie, c'est-à-dire le maintien d'un facteur physiologique à un niveau donné, le cortex insulaire contrôle des fonctions autonomes en régulant les systèmes nerveux sympathique et parasympathique[33] - [34]. Il a également un rôle dans la régulation du système immunitaire[35] - [36] - [37].
Conscience du Soi
Il a également été identifié un rôle de cette structure dans la conscience de son propre corps et la capacité de reconnaitre ce qui en fait partie ou non[38] - [39] - [40] et la sensation d'être l'auteur de ses actes (ou agentivité, sense of agency)[41].
Émotions sociales
L'insula antérieure est responsable du sentiment de dégout, que ce soit par les odeurs[42] ou par la vue d'une mutilation ou d'une infection[43], et ce même lorsque l'expérience est imaginée[44]. Ce mécanisme, ressemblant à celui supporté par les neurones miroirs fait donc le lien entre l'expérience intérieure et extérieure.
L'insula est également impliqué dans les comportements sociaux, notamment pour le sentiment de violation d'une norme[45], les processus émotionnels[46], l'empathie[47] ou encore l'orgasme[48].
Évolution
Le cortex insulaire est considéré comme un lobe à part entière du télencéphale par plusieurs neuroanatomistes. Néanmoins, d'autres sources le placent comme une partie du lobe temporal[49]. Il est même parfois rattaché aux structures limbiques, au sein d'un lobe limbique.
En tant que cortex paralimbique, le cortex insulaire est considéré comme une structure relativement ancienne à l'échelle de l'évolution. Il joue un rôle dans un grand nombre de fonctions hautement conservées, toutes liées aux besoins fondamentaux de survie, comme le goût, les sensations viscérales ou le contrôle autonome homéostatique. Mais plusieurs études tendent à montrer qu'en plus de ces fonctions conservées, l'insula pourrait jouer un rôle dans certaines fonctions « évoluées » présentes seulement chez l'humain et les autres grands singes. Il a été montré que l'insula antérieure contient une population de neurones appelés « neurones fusiformes » (spindle neurons). Ces neurones sont également présents au niveau du cortex cingulaire antérieur, une autre région très spécialisée chez les grands singes. Ces neurones sont présents en plus grande densité au niveau du cortex insulaire droit. Il a été supposé que ces neurones seraient impliqués dans des processus cognitifs/émotionnels chez les grands singes, comme l'empathie ou les émotions liées à la conscience du soi. Cette hypothèse est renforcée par des résultats d'imagerie fonctionnelle supportant un lien entre l'activité de l'insula antérieure droite avec la sensation de sentir son propre cœur battre ou encore de ressentir de l'empathie pour la douleur de quelqu'un. Cette fonction pourrait être liée au rôle de l'insula dans la prise de conscience des informations homéostatique.
Pathologies
Trouble de la personnalité narcissique
Une équipe de neurologues et de psychiatres ont étudié (grâce aux IRM) les implications possibles du cortex insulaire dans les manifestations comportementales de certains troubles de la personnalité : narcissique et borderline. Les résultats révèlent qu'il existe un manque de substance grise au niveau du cortex insulaire[50], plutôt qu'une absence totale de substance grise. Cette zone du cerveau étant impliquée dans les phénomènes d'empathie et de perception de la douleur, cela pourrait expliquer l'origine des difficultés que ces personnalités ont à se mettre à la place d'autrui.
Aphasie non-fluente progressive
L'aphasie non-fluente progressive est une détérioration du langage empêchant un individu de communiquer correctement, tout en conservant la capacité de comprendre les mots séparément et ses autres capacités cognitives. Elle est retrouvée dans de nombreuses pathologies neurodégénératives comme la maladie de Pick, la dégénérescence cortico-basale, la dégénérescence lobaire fronto-temporale ou encore la maladie d'Alzheimer. Elle est associée à un hypométabolisme[51] et une atrophie du cortex insulaire antérieur gauche[52].
Dépendance
De nombreuses études en imagerie fonctionnelle ont montré que le cortex insulaire était activé lorsque les toxicomanes sont exposés à des facteurs environnementaux déclenchant leur besoin de consommation. Cela a été montré pour de nombreuses drogues, comme la cocaïne, l'alcool, les opiacés et la nicotine. Néanmoins, cette implication reste négligée par la littérature sur l'addiction, principalement car l'insula n'est pas une cible directe connue du système mésotélencéphalique dopaminergique qui est le centre de la théorie du rôle du système de récompense dopaminergique dans la dépendance.
De récentes recherches ont montré que les fumeurs de cigarette qui ont subi un dommage au niveau de l'insula, par exemple à la suite d'un AVC, perdent pratiquement toute leur dépendance[53], néanmoins, la durée entre le traumatisme et l'étude étant d'en moyenne huit ans, les auteurs reconnaissent qu'un biais est possible[54]. Ils ont ainsi pu montrer que ces individus étaient 136 fois plus susceptibles de perdre leur addiction à la cigarette que les fumeurs touchés au niveau d'autres aires cérébrales. Les individus ont ainsi rapporté avoir quitté avec grande facilité, un jour seulement après la lésion, avec une très faible envie de rechute. Cela suggère que le cortex insulaire jouerait un rôle significatif dans le mécanisme neurologique des phénomènes d'addiction à la nicotine ou aux autres drogues, et en fait une cible d’intérêt pour le développement de nouveaux médicaments anti-addiction. Une autre possibilité évoquée est que les fonctions cognitives supportées par le cortex insulaire, comme la conscience, pourraient jouer un rôle majeur dans le maintien des addictions, même si cette hypothèse est délaissée par la littérature actuelle[55]. Une étude de 2007 chez le rat[56] semble corroborer ces découvertes en montrant qu'une inactivation réversible de l'insula désactive le conditionnement de localisation lié à la consommation d'amphétamine, un des modèles rongeurs de la dépendance aux drogues. Néanmoins, lors de cette étude, le placement conditionnel a été testé immédiatement après l'injection d'amphétamine, ce qui suggère que ce serait l'effet intéroceptif plaisant et immédiat de l'administration d'amphétamine, plutôt que l'effet tardif du manque d'amphétamine qui serait représenté dans l'insula.
Références
- (en) Henry Gray, Gray's Anatomy, (lire en ligne).
- (en) Binder DK, Schaller K, Clusmann H, « The seminal contributions of Johann-Christian Reil to anatomy, physiology, and psychiatry », Neurosurgery, vol. 61, no 5, , p. 1091–6; discussion 1096 (PMID 18091285, DOI 10.1227/01.neu.0000303205.15489.23, lire en ligne).
- (en) E MUFSON, « Insular interconnections with the amygdala in the rhesus monkey », Neuroscience, vol. 6, no 7, , p. 1231–1248 (DOI 10.1016/0306-4522(81)90184-6).
- Réalisés par l'équipe de Bud Craig.
- (en) Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ, « Neural systems supporting interoceptive awareness », Nat. Neurosci., vol. 7, no 2, , p. 189–95 (PMID 14730305, DOI 10.1038/nn1176).
- (en) Lamb K, Gallagher K, McColl R, Mathews D, Querry R, Williamson JW, « Exercise-induced decrease in insular cortex rCBF during postexercise hypotension », Med Sci Sports Exerc, vol. 39, no 4, , p. 672–9 (PMID 17414805, DOI 10.1249/mss.0b013e31802f04e0, lire en ligne).
- (en) Williamson JW, McColl R, Mathews D, Mitchell JH, Raven PB, Morgan WP, « Hypnotic manipulation of effort sense during dynamic exercise: cardiovascular responses and brain activation », J. Appl. Physiol., vol. 90, no 4, , p. 1392–9 (PMID 11247939, lire en ligne).
- (en) Williamson JW, McColl R, Mathews D, Ginsburg M, Mitchell JH, « Activation of the insular cortex is affected by the intensity of exercise », J. Appl. Physiol., vol. 87, no 3, , p. 1213–9 (PMID 10484598, lire en ligne).
- (en) Baliki MN, Geha PY, Apkarian AV, « Parsing pain perception between nociceptive representation and magnitude estimation », J. Neurophysiol., vol. 101, no 2, , p. 875–87 (PMID 19073802, DOI 10.1152/jn.91100.2008, lire en ligne).
- (en) Ogino Y, Nemoto H, Inui K, Saito S, Kakigi R, Goto F, « Inner experience of pain: imagination of pain while viewing images showing painful events forms subjective pain representation in human brain », Cereb. Cortex, vol. 17, no 5, , p. 1139–46 (PMID 16855007, DOI 10.1093/cercor/bhl023, lire en ligne).
- (en) Song GH, Venkatraman V, Ho KY, Chee MW, Yeoh KG, Wilder-Smith CH, « Cortical effects of anticipation and endogenous modulation of visceral pain assessed by functional brain MRI in irritable bowel syndrome patients and healthy controls », Pain, vol. 126, nos 1-3, , p. 79–90 (PMID 16846694, DOI 10.1016/j.pain.2006.06.017, lire en ligne).
- (en) Olausson H, Charron J, Marchand S, Villemure C, Strigo IA, Bushnell MC, « Feelings of warmth correlate with neural activity in right anterior insular cortex », Neurosci. Lett., vol. 389, no 1, , p. 1–5 (PMID 16051437, DOI 10.1016/j.neulet.2005.06.065, lire en ligne).
- (en) Craig AD, Chen K, Bandy D, Reiman EM, « Thermosensory activation of insular cortex », Nat. Neurosci., vol. 3, no 2, , p. 184–90 (PMID 10649575, DOI 10.1038/72131).
- (en) Ladabaum U, Minoshima S, Hasler WL, Cross D, Chey WD, Owyang C, « Gastric distention correlates with activation of multiple cortical and subcortical regions », Gastroenterol, vol. 120, no 2, , p. 369–76. (PMID 11159877, DOI 10.1053/gast.2001.21201, lire en ligne).
- (en) Hamaguchi T, Kano M, Rikimaru H. et al., « Brain activity during distention of the descending colon in humans », Neurogastroenterol. Motil., vol. 16, no 3, , p. 299–309 (PMID 15198652, DOI 10.1111/j.1365-2982.2004.00498.x, lire en ligne).
- (en) Matsuura S, Kakizaki H, Mitsui T, Shiga T, Tamaki N, Koyanagi T, « Human brain region response to distention or cold stimulation of the bladder: a positron emission tomography study », J. Urol., vol. 168, no 5, , p. 2035–9 (PMID 12394703, DOI 10.1097/01.ju.0000027600.26331.11, lire en ligne).
- (en) A. von Leupoldt, « The Unpleasantness of Perceived Dyspnea Is Processed in the Anterior Insula and Amygdala », American Journal of Respiratory and Critical Care Medicine, vol. 177, no 9, , p. 1026–1032. (DOI 10.1164/rccm.200712-1821OC, lire en ligne).
- (en) Kikuchi M, Naito Y, Senda M. et al., « Cortical activation during optokinetic stimulation — an fMRI study », Acta Otolaryngol., vol. 129, no 4, , p. 440–3. (PMID 19116795, DOI 10.1080/00016480802610226, lire en ligne).
- (en) Papathanasiou ES, Papacostas SS, Charalambous M, Eracleous E, Thodi C, Pantzaris M, « Vertigo and imbalance caused by a small lesion in the anterior insula », Electromyogr Clin Neurophysiol, vol. 46, no 3, , p. 185–92 (PMID 16918202).
- (en) Brown S, Martinez MJ, Parsons LM, « Passive music listening spontaneously engages limbic and paralimbic systems », Neuroreport, vol. 15, no 13, , p. 2033–7 (PMID 15486477, DOI 10.1097/00001756-200409150-00008, lire en ligne).
- (en) Sander K, Scheich H, « Left auditory cortex and amygdala, but right insula dominance for human laughing and crying », J Cogn Neurosci, vol. 17, no 10, , p. 1519–31. (PMID 16269094, DOI 10.1162/089892905774597227, lire en ligne).
- « http://ccare.stanford.edu/node/89 »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ).
- (en) Bamiou DE, Musiek FE, Luxon LM, « The insula (Island of Reil) and its role in auditory processing. Literature review », Brain Res. Brain Res. Rev., vol. 42, no 2, , p. 143–54 (PMID 12738055, DOI 10.1016/S0165-0173(03)00172-3, lire en ligne).
- (en) Anderson TJ, Jenkins IH, Brooks DJ, Hawken MB, Frackowiak RS, Kennard C, « Cortical control of saccades and fixation in man. A PET study », Brain, vol. 117, no Pt 5, , p. 1073–84 (PMID 7953589, DOI 10.1093/brain/117.5.1073, lire en ligne).
- (en) Fink GR, Frackowiak RS, Pietrzyk U, Passingham RE, « Multiple nonprimary motor areas in the human cortex », J. Neurophysiol., vol. 77, no 4, , p. 2164–74 (PMID 9114263, lire en ligne).
- (en) Sörös P, Inamoto Y, Martin RE, « Functional brain imaging of swallowing: an activation likelihood estimation meta-analysis », Hum Brain Mapp, vol. 30, no 8, , p. 2426–39 (PMID 19107749, DOI 10.1002/hbm.20680).
- (en) Penfield W, Faulk ME, « The insula; further observations on its function », Brain, vol. 78, no 4, , p. 445–70 (PMID 13293263, lire en ligne).
- (en) Dronkers NF, « A new brain region for coordinating speech articulation », Nature, vol. 384, no 6605, , p. 159–61 (PMID 8906789, DOI 10.1038/384159a0).
- (en) Ackermann H, Riecker A, « The contribution of the insula to motor aspects of speech production: a review and a hypothesis », Brain Lang, vol. 89, no 2, , p. 320–8 (PMID 15068914, DOI 10.1016/S0093-934X(03)00347-X, lire en ligne).
- (en) Nowak M, Holm S, Biering-Sørensen F, Secher NH, Friberg L, « "Central command" and insular activation during attempted foot lifting in paraplegic humans », Hum Brain Mapp, vol. 25, no 2, , p. 259–65 (PMID 15849712, DOI 10.1002/hbm.20097).
- (en) Borovsky A, Saygin AP, Bates E, Dronkers N, « Lesion correlates of conversational speech production deficits », Neuropsychologia, vol. 45, no 11, , p. 2525–33 (PMID 17499317, DOI 10.1016/j.neuropsychologia.2007.03.023, lire en ligne).
- (en) Mutschler I, Schulze-Bonhage A, Glauche V, Demandt E, Speck O, Ball T, « A rapid sound-action association effect in human insular cortex », PLoS ONE, vol. 2, no 2, , e259 (PMID 17327919, PMCID 1800344, DOI 10.1371/journal.pone.0000259, lire en ligne).
- (en) Oppenheimer SM, Gelb A, Girvin JP, Hachinski VC, « Cardiovascular effects of human insular cortex stimulation », Neurology, vol. 42, no 9, , p. 1727–32 (PMID 1513461).
- (en) Critchley HD, « Neural mechanisms of autonomic, affective, and cognitive integration », J. Comp. Neurol., vol. 493, no 1, , p. 154–66 (PMID 16254997, DOI 10.1002/cne.20749)
- (en) Pacheco-López G, Niemi MB, Kou W, Härting M, Fandrey J, Schedlowski M, « Neural substrates for behaviorally conditioned immunosuppression in the rat », J. Neurosci., vol. 25, no 9, , p. 2330–7 (PMID 15745959, DOI 10.1523/JNEUROSCI.4230-04.2005, lire en ligne).
- (en) Ramírez-Amaya V, Alvarez-Borda B, Ormsby CE, Martínez RD, Pérez-Montfort R, Bermúdez-Rattoni F, « Insular cortex lesions impair the acquisition of conditioned immunosuppression », Brain Behav. Immun., vol. 10, no 2, , p. 103–14 (PMID 8811934, DOI 10.1006/brbi.1996.0011, lire en ligne).
- (en) Ramírez-Amaya V, Bermúdez-Rattoni F, « Conditioned enhancement of antibody production is disrupted by insular cortex and amygdala but not hippocampal lesions », Brain Behav. Immun., vol. 13, no 1, , p. 46–60 (PMID 10371677, DOI 10.1006/brbi.1998.0547, lire en ligne).
- (en) Karnath HO, Baier B, Nägele T, « Awareness of the functioning of one's own limbs mediated by the insular cortex? », J. Neurosci., vol. 25, no 31, , p. 7134–8 (PMID 16079395, DOI 10.1523/JNEUROSCI.1590-05.2005, lire en ligne).
- (en) Craig AD, « How do you feel—now? The anterior insula and human awareness », Nat. Rev. Neurosci., vol. 10, no 1, , p. 59–70 (PMID 19096369, DOI 10.1038/nrn2555).
- (en) Tsakiris M, Hesse MD, Boy C, Haggard P, Fink GR, « Neural signatures of body ownership: a sensory network for bodily self-consciousness », Cereb. Cortex, vol. 17, no 10, , p. 2235–44 (PMID 17138596, DOI 10.1093/cercor/bhl131, lire en ligne).
- (en) Farrer C, Frith CD, « Experiencing oneself vs another person as being the cause of an action: the neural correlates of the experience of agency », Neuroimage, vol. 15, no 3, , p. 596–603 (PMID 11848702, DOI 10.1006/nimg.2001.1009, lire en ligne).
- (en) Wicker B, Keysers C, Plailly J, Royet JP, Gallese V, Rizzolatti G, « Both of us disgusted in My insula: the common neural basis of seeing and feeling disgust », Neuron, vol. 40, no 3, , p. 655–64 (PMID 14642287, DOI 10.1016/S0896-6273(03)00679-2, lire en ligne).
- (en) Wright P, He G, Shapira NA, Goodman WK, Liu Y, « Disgust and the insula: fMRI responses to pictures of mutilation and contamination », Neuroreport, vol. 15, no 15, , p. 2347–51 (PMID 15640753, DOI 10.1097/00001756-200410250-00009, lire en ligne).
- (en) Jabbi M, Bastiaansen J, Keysers C, « A common anterior insula representation of disgust observation, experience and imagination shows divergent functional connectivity pathways », PLoS ONE, vol. 3, no 8, , e2939 (PMID 18698355, PMCID 2491556, DOI 10.1371/journal.pone.0002939, lire en ligne).
- (en) Sanfey AG, Rilling JK, Aronson JA, Nystrom LE, Cohen JD, « The neural basis of economic decision-making in the Ultimatum Game », Science, vol. 300, no 5626, , p. 1755–8 (PMID 12805551, DOI 10.1126/science.1082976, lire en ligne).
- (en) Phan KL, Wager T, Taylor SF, Liberzon I, « Functional neuroanatomy of emotion: a meta-analysis of emotion activation studies in PET and fMRI », Neuroimage, vol. 16, no 2, , p. 331–48 (PMID 12030820, DOI 10.1006/nimg.2002.1087, lire en ligne).
- (en) Singer T, « The neuronal basis and ontogeny of empathy and mind reading: review of literature and implications for future research », Neurosci Biobehav Rev, vol. 30, no 6, , p. 855–63 (PMID 16904182, DOI 10.1016/j.neubiorev.2006.06.011, lire en ligne).
- (en) Ortigue S, Grafton ST, Bianchi-Demicheli F, « Correlation between insula activation and self-reported quality of orgasm in women », Neuroimage, vol. 37, no 2, , p. 551–60 (PMID 17601749, DOI 10.1016/j.neuroimage.2007.05.026, lire en ligne).
- Kolb, Bryan; Whishaw, Ian Q. (2003). Fundamentals of human neuropsychology (5th ed.). [New York]: Worth. (ISBN 0-7167-5300-6).
- Lars Schulze, Isabel Dziobek, Aline Vater et Hauke R. Heekeren, « Gray matter abnormalities in patients with narcissistic personality disorder », Journal of Psychiatric Research, vol. 47, no 10, , p. 1363–1369 (ISSN 1879-1379, PMID 23777939, DOI 10.1016/j.jpsychires.2013.05.017, lire en ligne, consulté le )
- (en) Nestor PJ, Graham NL, Fryer TD, Williams GB, Patterson K, Hodges JR, « Progressive non-fluent aphasia is associated with hypometabolism centred on the left anterior insula », Brain, vol. 126, no Pt 11, , p. 2406–18 (PMID 12902311, DOI 10.1093/brain/awg240, lire en ligne).
- (en) Gorno-Tempini ML, Dronkers NF, Rankin KP, et al., « Cognition and anatomy in three variants of primary progressive aphasia », Ann. Neurol., vol. 55, no 3, , p. 335–46 (PMID 14991811, PMCID 2362399, DOI 10.1002/ana.10825).
- Nasir H. Naqvi, David Rudrauf, Hanna Damasio, Antoine Bechara., « Damage to the Insula Disrupts Addiction to Cigarette Smoking », Science, vol. 315, no 5811, , p. 531–4 (PMID 17255515, DOI 10.1126/science.1135926, lire en ligne [abstract]).
- (en) Vorel SR, Bisaga A, McKhann G, Kleber HD, « Insula damage and quitting smoking », Science, vol. 317, no 5836, , p. 318–9; author reply 318–9 (PMID 17641181, DOI 10.1126/science.317.5836.318c, lire en ligne).
- Steven E. Hyman, « Addiction: A Disease of Learning and Memory », Am J Psychiatry, vol. 162, no 8, , p. 1414–22 (PMID 16055762, DOI 10.1176/appi.ajp.162.8.1414, lire en ligne [abstract]).
- Marco Contreras, Francisco Ceric, Fernando Torrealba, « Inactivation of the Interoceptive Insula Disrupts Drug Craving and Malaise Induced by Lithium », Science, vol. 318, no 5850, , p. 655–8 (PMID 17962567, DOI 10.1126/science.1145590, lire en ligne [abstract]).
- (en) Paulus MP, Stein MB, « An insular view of anxiety », Biol. Psychiatry, vol. 60, no 4, , p. 383–7 (PMID 16780813, DOI 10.1016/j.biopsych.2006.03.042, lire en ligne).
- (en) Thayer JF, Lane RD, « A model of neurovisceral integration in emotion regulation and dysregulation », J Affect Disord, vol. 61, no 3, , p. 201–16 (PMID 11163422, DOI 10.1016/S0165-0327(00)00338-4, lire en ligne).
Bibliographie
- Olivier Houdé, Bernard Mazoyer et Nathalie Tzourio-Mazoyet, Cerveau et psychologie : Introduction à l'imagerie cérébrale anatomique et fonctionnelle, PUF, .