Cortex visuel
Le cortex visuel occupe le lobe occipital du cerveau et est chargé de traiter les informations visuelles.
Le cortex visuel couvre le lobe occipital, sur les faces latérales et internes, et empiète sur le lobe pariétal et le lobe temporal.
L'étude du cortex visuel[1] en neurosciences a permis de le découper en une multitude de sous-régions fonctionnelles (V1, V2, V3, V4, MT, etc.) qui traitent chacune ou collectivement des multiples propriétés des informations provenant des voies visuelles (formes, couleurs, mouvements, etc.). Comme pour les autres sens, la décussation des voies nerveuses fait que la moitié droite du champ visuel est analysée par l'hémisphère gauche et inversement, ce qui explique la latéralisation des troubles visuels résultant d'une lésion cérébrale. Parmi ces troubles, les agnosies visuelles correspondent à l'incapacité de percevoir certaines propriétés d'un stimulus visuel tout en gardant une vision parfaite pour le reste. Cette condition pathologique résulte d'une lésion focale de l'aire responsable du traitement de l'information en question : par exemple, une destruction de l'aire V4 qui traite les couleurs rend le patient « aveugle » aux couleurs, c'est-à-dire achromate mais le reste de sa vision est parfaitement normale.
Cartographie corticale du champ visuel
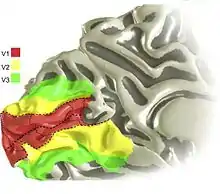
Environ la moitié du néocortex des primates non humains est couverte par des aires visuelles[2]. Chez l'Homme, on estime qu'elles représentent 20 à 25 % du cortex[3], et qu'y opèrent environ 5 milliards de neurones.
Plus d'une vingtaine d'aires visuelles ont été distinguées sur le cortex des primates (l'homme y compris) sur la base de leur architecture, de leur connexions, de leur topographie visuelle et de leurs propriétés fonctionnelles.
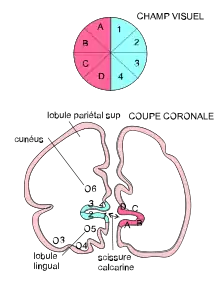
L'ancienne subdivision de Brodmann basée sur la forme des neurones (ou cytoarchitectonie), distinguait les aires 17, 18 et 19. L'aire 17 correspond approximativement au cortex visuel primaire V1 ou cortex strié. Sa délimitation ne pose pas de problème, en raison de frontières myéloarchitectoniques proéminentes et peu distordues. Elle constitue le point d'entrée dans le cortex visuel des informations visuelles provenant du corps genouillé latéral (du thalamus) et plus en amont de la rétine de l’œil. Elle est située sur la face interne des deux hémisphères, sur les berges de la scissure calcarine (fig. 0, 1 et 5).
Les aires de Brodmann 18 et 19 sont elles, plus difficiles à séparer[4]. Pour Brodmann, l'aire 18 (ou cortex parastrié) entoure l'aire visuelle primaire V1 et est entourée par l'aire 19 (ou cortex extrastrié). La séparation classiquement établie sur la base de la densité des cellules pyramidales de la sous-couche IIIb s'avère peu opérante. Par contre, cette région corticale présente la propriété remarquable de donner une représentation cartographique du champ visuel : les objets du champ visuel, associés à des figures d'activation de la rétine, se retrouvent projetés en figures plus ou moins continûment déformées dans différentes zones du cortex. Cette organisation dite rétinotopique permet de délimiter les aires visuelles rétinotopiques « classiques » V1, V2, V3, V3A, V3B, V4v couvrant en gros les aires de Brodmann visuelles.
Définition des cartes rétinotopiques
Une relation rétinotopique[N 1] est une application bijective continue, qui à deux points voisins de la rétine, fait correspondre deux points voisins de la surface corticale[5]. On peut considérer qu'il s'agit d'un homéomorphisme entre une partie du champ rétinien et une aire corticale. Mathématiquement, dire que deux objets sont homéomorphes revient à dire que l'on peut passer de l'un à l'autre par une déformation continue. Une carte rétinotopique corticale est donc une projection homéomorphe d'une partie du champ visuel (ou rétinien) sur le cortex.
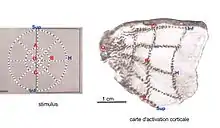
La détermination de ces cartes rétinotopiques a été faite directement sur le cerveau d'animaux en leur implantant des électrodes sur le cortex et en mesurant des centaines de champs récepteurs[2]. Une méthode tout aussi invasive mais plus simple consiste à utiliser un marqueur comme le 14C-2-désoxyglucose radioactif qui n'étant pas métabolisé, s'accumule dans les neurones actifs. Un autoradiogramme[N 2] d'une portion de la surface corticale aplatie peut ensuite révéler les régions les plus actives durant la stimulation[6]. Cette méthode permet de voir s'inscrire directement la carte rétinotopique sur la surface corticale en un temps assez court. Tootell et al (1988) ont pu ainsi obtenir des cartes très parlantes en stimulant la rétine de macaques avec des cercles parallèles et des rayons (méridiens) faits de carreaux noirs et blancs clignotants (fig. 2). Les points radiomarqués des cercles et des méridiens du stimulus visuel s'inscrivent ainsi sur le cortex sous forme de lignes de points plus ou moins orthogonales.
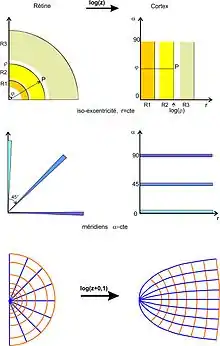
Par z→log(z), les cercles sont envoyés sur des verticales et les rayons sur des horizontales (schéma du haut et du milieu).
Schéma du bas : pour obtenir la convergence des méridiens sur le centre de la fovéa, il faut prendre une application z→log(z+a).
Les points du champ visuel sont facilement repérés par des coordonnées polaires (r, α) avec r l'excentricité[N 3] et α l'angle polaire (avec une droite). L'application rétinotopique peut être décrite dans une première approximation[7] - [8] comme une transformation conforme (conservant les angles) de type logarithme complexe, dans laquelle la composante r est déformée suivant la fonction logarithme pour refléter le grossissement de la région centrale de la fovéa sur le cortex[N 4] (fig. 3).
À ce concept théorique d'application rétinotopique, très éclairante pour bien comprendre le mécanisme sous-jacent, il faut ajouter un concept empirique, indispensable pour dresser les cartes rétinitopiques du cortex. Wandell et ses collègues[9] - [3] ont développé un concept opératoire des cartes rétinotopiques qu'ils appellent plus simplement cartes de champ visuel, cartes acquises par la méthode de l'onde mobile (présentée plus bas) qui doivent être distinguées des aires visuelles définies sur des critères différents (architecturaux, de connectivité, fonctionnel).
Une carte de champ visuel contient au plus une représentation de chaque point du champ visuel. Il en découle que deux parties du cortex répondant préférentiellement à la même position du stimulus dans le champ visuel, doivent se trouver sur des cartes différentes. Wandell demande aussi dans sa définition qu'une carte de champ visuel représente la totalité ou une partie substantielle du champ visuel. Enfin, l'application rétinotopique peut comporter quelques discontinuités : ainsi, bien que l'on puisse passer continûment de l'hémichamp visuel gauche à l'hémichamp droit, il n'en est pas de même dans le cortex où la projection saute d'un hémisphère à l'autre. De même les aires V2 et V3 sont divisées par le méridien horizontal dans chaque hémisphère, ce qui a pour conséquence que les cartes rétinotopiques sont divisées en 4 quadrants disjoints.
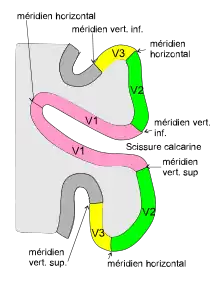
En raison du croisement des fibres provenant de l'hémirétine nasale au niveau du chiasma optique, les hémichamps visuels se projettent contralatéralement. Dans les aires visuelles de bas niveau, la fovéa se projette près du pôle occipital et les excentricités périphériques se projettent plus antérieurement. Le quadrant supérieur (resp. inférieur) est représenté sur la berge inférieure (res. supérieure) de la scissure calcarine et le méridien horizontal se retrouve le long du fond de la scissure calcarine. Le méridien vertical est projeté sur la frontière de V1 et V2 (fig. 5).
Méthode d'acquisition des cartes rétinotopiques
Il a longtemps été impossible d'obtenir des cartes rétinotopiques précises chez l'homme par des méthodes non invasives, en raison de la mauvaise résolution spatiale de la neuro-imagerie et des repliements importants de la surface corticale cachant les 2/3 du cortex. Un progrès significatif a été obtenu dans les années 1990, lorsque Engel et al.[10] - [11] et Sereno et al.[12] ont utilisé un stimulus périodique (plutôt qu'un stimulus fixe) parcourant le champ visuel et ont représenté la carte acquise par IRMf, sur une image du cortex mise à plat. Le stimulus formé par un anneau en expansion produit une vague d'activation parcourant le cortex visuel d'arrière en avant dont on mesure la phase. En analysant ces phases par la transformée de Fourier et les reportant sur la surface corticale dépliée en deux dimensions, on peut obtenir ensuite automatiquement des cartes avec des frontières clairement objectivées[13].
Dans la méthode de l'onde mobile (ou d'encodage de phase), on procède donc à deux types d'acquisitions :
- par une IRM à haute résolution, on obtient une image anatomique 3D qui sera ensuite segmentée[N 5] puis dépliée ;
- par une série IRMf, on extrait des cartes de phases en 3D d'isoexcenticité acquises par des stimuli d'anneaux en contraction et en expansion et des cartes de phases d'isopolarité (de même angle de direction) acquises par des stimuli formés de secteurs tournant dans le sens positif ou négatif. La combinaison des informations de ces deux cartes de phases sur la surface corticale dépliée permet de délimiter les aires visuelles (fig. 4).
Deux types de régions corticales sont distinguées suivant que leur carte rétinotopique forme une image en miroir du champ visuel (comme l'aire V1) ou au contraire une image non en miroir (comme l'aire V2). La représentation est à nouveau inversée en V3. On a ainsi une alternance des cartes en miroir / non en miroir d'une aire à son aire adjacente (fig. 5). On dit que le signe local du champ visuel est négatif pour V1 et V3 et positif pour les aires V2, V4 et V3A.
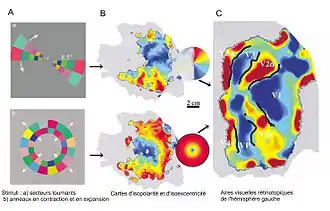
A : stimuli formés par (a) des secteurs tournant dans un sens ou l'autre, (b) des anneaux en expansion et contraction parcourant le champ visuel
B : carte de phases d'isopolarité dépliée: le quadrant supérieur, droit, codé en rouge et jaune se projette sur la partie inférieure de l'hémisphère gauche, la variation de phase se fait dans une direction perpendiculaire à la scissure calcarine
et carte de phases d'isoexcentricité dépliées: la zone de la fovéa se projettent en bleu dans la région postérieure, la phase varie en partie parallèlement à la scissure
C : cartes rétinotopiques obtenues par combinaison des cartes de phases, d'après Warnking et al.[13] (2002).
Les aires visuelles rétinotopiques
Les cartes de champ visuel corticales en préservant la structure spatiale de la scène facilitent les calculs impliquant des comparaisons entre neurones spécialisés dans le traitement de la couleur, du mouvement ou de l'orientation.
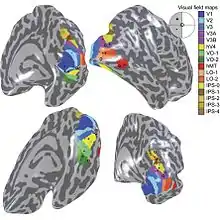
Grâce à la méthode de l'onde mobile, une quinzaine d'aires visuelles rétinotopiques[3], ont été caractérisées chez l'homme. Toutes les études s'accordent sur la localisation et les propriétés des aires visuelles V1, V2, V3 et V3A. Au-delà, pour les aires visuelles de plus haut niveau, certains désaccords entre les équipes ne sont pas encore complètement résolus[14].
- Les cartes médiales-postérieures : V1, V2, V3
L'aire V1 est située sur les berges de la scissure calcarine et s'étend parfois au-delà du pôle postérieur sur la face ventrale latérale[9]. C'est une carte rétinotopique de l'hémichamp contralatéral, impliquée dans la détection des contrastes.
Les cartes V2 et V3 forment deux rubans corticaux, de 1 à 3 cm de large, entourant V1. Elles comportent des cartes rétinotopiques des hémichamps contralatéraux avec une discontinuité au niveau du méridien horizontal. Ainsi l'aire V2 est composée d'une partie dorsale V2d et d'une partie ventrale V2v, de même V3 se décompose en V3d et V3v (parfois nommée VP). Le passage continu du quadrant supérieur au quadrant inférieur dans le champ visuel ne peut se faire continûment sur la carte. Chacune de ces parties cartographie donc isolément un quadrant du champ visuel. Elles occupent une partie de l'aire 18 de Brodmann et font partie du cortex extrastrié.
Dans V2, on distingue trois types de bandes : bandes fines, bandes épaisses et intermédiaires. Les bandes épaisses sont sensibles à l’orientation et à la direction du mouvement (voie M magno), tandis que les bandes fines sont plutôt sensibles à la couleur (voie P parvo).
L'aire V3 reçoit des afférences des bandes épaisses de V2, ainsi que des connexions directes à V1. L'aire V3 possède une sélectivité à l'orientation et au mouvement. Les neurones y ont une haute sensibilité au contraste.
- Cartes dorsales : V3A, V3B, IPS-x
Sur la face dorsale du cerveau, à l'arrière du sillon intrapariétal, se trouvent les cartes rétinotopiques V3A, V3B et dans le sillon intrapariétal V7 et IPS-1/2/3/4. La carte de l'aire V3A borde antérieurement la carte V3d et remonte sur la partie postérieure du sillon intrapariétal, sur la face dorsale. Une autre carte, nommée V3B se situe en position dorsale latérale de V3A[3] et partage une fovéa avec V3A. La carte V7 se situe sur la frontière antérieure de V3A, dans le sillon intrapariétal (IPS). Elle possède une représentation de la fovéa confluente avec la carte rétinotopique IPS-1, suggérant son autre nom de IPS-0. Plusieurs cartes rétinotopiques ont depuis été décrites le long du sillon intrapariétal (IPS) et nommées IPS-1/2/3/4, dans la région antérieure à IPS-0.
La région V3A, V3B répond aux taches liées à la perception du mouvement et de la profondeur. Des lésions dans cette région sont associées à des déficits de perception du mouvement.
- Cartes latérales : LO-1, LO-2, hMT
Il y a aussi un bon accord sur la présence d'une carte rétinotopique hMT+ (ou MT[N 6] ou aire V5) de sélection de mouvement, sur la face latérale du lobe occipital près du lobe temporal (Wandell[9]). Des lésions dans la région MT sont associées à l'impossibilité de percevoir le mouvement (akinétopsie). Une perturbation de cette région causée par stimulation magnétique transcranienne interfère spécifiquement avec la perception du mouvement.
Deux cartes sur le cortex latéral occipital (LO), situées entre V3 et MT, ont aussi été identifiées[15] sous divers noms et ici nommées LO-1 et LO-2[3]. Il s'agirait de représentation d'hémichamp complet. Cette région répond différemment à un stimulus d'onde mobile que les régions du cortex visuel primaire car les champs récepteurs sont considérablement plus grands (environ x5 que dans V1). La carte polaire de LO-1 est une image en miroir de V3 et celle de LO-2 est en miroir de LO-1 (comme il apparait clairement sur la carte de l'angle polaire de la figure 7).
- Cartes ventrales : V4, VO-1, VO-2
La face ventrale du cortex occipito-temporal est connue pour répondre puissamment aux taches de reconnaissance d'objets comme les visages, les objets, les textes ou des motifs colorés. Des lésions dans cette région sont associées à l'incapacité à reconnaître les visages familiers, des dysfonctions de reconnaissance des couleurs et de l'alexie. Des désaccords entre laboratoires subsistent dans l'identification des cartes de cette région car la méthode de l'onde mobile demande à être perfectionnée pour couvrir des zones avec de grands champs récepteurs couvrant la fovéa. Le groupe de Wandell[9] a reconnu dans cette région la carte d'hémichamp visuel hV4 (h pour humain), adjacente à la partie ventrale de V3v et possédant une représentation fovéale confluente avec V1, V2 et V3.
Deux cartes d'hémichamp VO-1 et VO-2 du cortex ventral-occipital ont aussi été décrites en position antérieure à hV4. Le groupe de Zeki[16] décrit un complexe V4, recouvrant les cartes hV4 et VO de Wandell, et jouant un rôle essentiel dans la perception des couleurs. Le débat sur la fonction de V4 n'est pas résolu.
Aires visuelles de moyen et haut niveau
La méthode de cartographie IRM par onde mobile permet de détecter les aires rétinotopiques V1, V2, V3,V3A, V3B, V4, V5 (ou MT), VO-1/2, LO-1/2, et IPS-x, avec encore quelques incertitudes pour certaines. En utilisant des stimuli optimisés pour activer les aires visuelles de moyen et haut niveau, on peut localiser de plus en plus d'aires visuelles faisant partie de ce qui était communément appelé « le cortex non-rétinotopique »[17] - [14]. On s'aperçoit maintenant que la représentation ordonnée du champ visuel se retrouve sur tout le cortex visuel, mais de façon moins précise au fur et à mesure que l'on monte dans la hiérarchie. On trouve même dans le lobe frontal des activations rétinotopiques (FEF, frontal eye fields[17]).
Organisation des cartes de champ visuel
De la rétine au cortex strié, via le CGL, les propriétés de forme, couleur et contraste du signal visuel sont traités en parallèle le long des voies optiques P, M et K. Ces voies aboutissent dans les colonnes de dominance du cortex strié V1 où ces propriétés sont ré-associées. Les informations une fois traitées par le cortex visuel primaire V1 sont transférées vers les aires dites extrastriées où des traitements plus approfondis ont lieu.
Quelques modèles plus ou moins élaborés ont été proposés. Deux principes gouvernent les explications : un principe de spécialisation fonctionnelle (des voies distinctes se spécialisent dans le traitement d'un certain type d'information comme dans le modèle des deux voies), un principe hiérarchique (le traitement graduel de l'information se fait dans une séquence allant vers des représentations plus abstraites).
- Modèle des deux voies (ventrale et dorsale) (Ungerleider et Mishkin 1982)

Les informations issues de l'aire V1 sont séparées en deux voies distinctes au-delà des aires V2 et V3 :
- La voie ventrale, dite aussi la « voie du Quoi ? » dans laquelle les informations de forme et de couleurs sont analysées afin d'aboutir à une reconnaissance des objets présents dans la scène visuelle.
- La voie dorsale, dite aussi la « voie du Où ? » dans laquelle les informations liées au mouvement et aux positions dans l'espace sont analysées de façon à permettre les mouvements en direction ou non des objets présents (saccade oculaire, mouvement de préhension...)
Ces deux types de traitement s'effectuent simultanément. Le long de ces voies les champs récepteurs sont de plus en plus grands et la largeur de la bande de perception chromatique diminue.
- Modèle hiérarchique
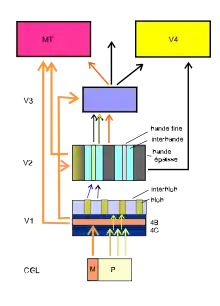
Van Essen et ses collègues (1991) ont proposé un modèle hiérarchique du cortex visuel du macaque sous forme d'un graphe organisant hiérarchiquement les aires visuelles. Pour le cortex humain, Vasseur[14] propose aussi un schéma simplifié de la connexion de quelques cartes rétinotopiques classiques.
La voie M part des cellules ganglionnaires en parasol de la rétine, fait relais dans les deux couches magnocellulaires ventrales du corps géniculé latéral (CGL) puis envoie des projections géniculo-striées sur la couche 4B, 4Cα de l'aire V1. La ségrégation du traitement de l'information assurée dans V1 est aussi préservée dans l'aire V2. La carte V2 est constituée de trois types de bandes : bande épaisse, inter-bande et bande fine. Les bandes épaisses, sensibles à l’orientation et à la direction du mouvement, reçoivent l'information de la sous-couche 4B[4]. Cette voie se prolonge dans la partie dorsale de V3 et dans MT (V5). Ce sont des neurones sensibles au mouvement.
La voie P part des cellules ganglionnaires naines de la rétine, fait relais dans les 4 couches parvocellulaires du CGL puis aboutit sur la couche 4Cβ de V1 où elle passe par les interblobs pour aboutir sur les interbandes de V2. De là, elle se dirige vers V3 et V4. Ces neurones sont sensibles à l'orientation et la forme.
La voie K part des cellules ganglionnaires bistratifiées de la rétine, fait relais dans les 6 sous-couches koniocellulaires du CGL, puis aboutit dans les couches 2, 3 (avec ses blobs) de l'aire V1. De là, des connexions aboutissent sur V3 et V4. Ces neurones sont sensibles à la couleur.
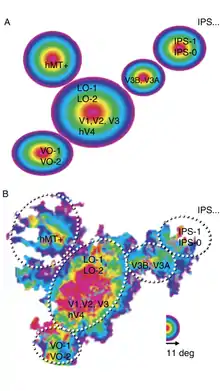
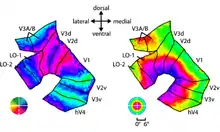
B : Cortex de l'hémisphère gauche d'un sujet mis à plat avec les mesures d'excentricité prises entre 1 et 11°. Les pointillés délimitent les agrégats, d'après Wandell et al.[3].
Wandell et ses collègues proposent d'associer les cartes rétinotopiques en clusters (agrégats). Un agrégat est un groupe de cartes de champ visuel ayant des représentations d'excentricité parallèles.
Un bel agrégat « prototypique » constitué par V1, V2, V3, V4, LO-1 et LO-2 a été mis en évidence par la carte d'excentricité (fig. 7) de Larsson et Heeger[15] dans une représentation où le cortex est mis à plat. On voit ces six aires visuelles confluer sur la même région fovéale (sur la carte d'excentricité) alors que les aires V3A et V3B sont repoussées sur le côté et forment ensemble un autre agrégat. Les autres agrégats dégagés sur la figure 10 par Wandell et al. sont l'agrégat ventral {VO-1, VO-2}, latéral {MT} (isolé des cartes LO-1/2) et celui du sillon intrapariétal {IPS-1, IPS-0} (les cartes IPS-2/3/4 n'ont pas été retenues dans ces données).
L'hypothèse des auteurs est que différentes cartes s'organiseraient en un agrégat pour faciliter le traitement de l'information de la mosaïque neurale.
Plasticité neurologique
Le système visuel cortical montre, à l'instar des autres structures corticales, une grande plasticité. Ainsi, lors d'une atteinte sélective du champ visuel (tache aveugle), l'aire neuronale correspondant à la lésion se réduit progressivement au profit des cellules rétiniennes voisines : les neurones non stimulés sont connectés aux neurones voisins, ce qui a pour effet de « masquer » le déficit.
Des expériences récentes menées chez le hamster nouveau-né indiquent également que le cortex auditif peut se substituer au cortex visuel en cas d'absence (embryonnaire) de ce dernier.
Cortex visuel et endohormones cérébrales
Il a été maintes fois constaté et démontré que l'œstrogène 17β-estradiol (E2) affecte le traitement perceptif des indices visuels[18].
On a d'abord supposé que c'était l'E2 d'origine gonadique qui influençait ce processus, sans d'ailleurs comprendre pourquoi[18].
Une hypothèse plus récente (2011) est que dans la zone cérébrale consacrée à cette tâche, le « traitement des images » puisse aussi être modulée par des hormones ayant le même récepteur, mais sécrétées par le cerveau lui-même[18].
Et effectivement, on a expérimentalement récemment montré chez la souris de laboratoire que des circuits neuronaux associés aux œstrogènes sont très répandues dans le cortex visuel primaire (V1)[18] ; Une équipe universitaire américaine a montré - via d'une part l'aromatase utilisée comme marqueur de neurones producteurs d'œstrogène, et d'autre part via les récepteurs classiques des œstrogènes (ERa et ERß[19] utilisés comme marqueurs de neurones sensibles aux œstrogènes - que les deux parties (monoculaire et binoculaire) du cortex primaire (V1) de la souris sont très riches en aromatase et en neurones sensibles aux œstrogènes. Ceci laisse penser que le cortex visuel V1 est effectivement à la fois un site de production d'œstrogènes et un site sensible à des œstrogènes produits localement. Les fonctions locales cette hormone restent à éclaircir[18].
La fluorescence double hybridation in situ a précisé l'identité neurochimique de ces œstrogènes et montré l'existence d'une population hétérogène neuronale associée. La même étude a montré que l'expérience visuelle engageait une large population de neurones positifs à l'aromatase (producteurs d'œstrogènes) et, mais dans une moindre mesure, de neurones récepteurs, ce qui suggère que les niveaux d'E2 pourraient être régulés par un apport visuel dans le cortex visuel primaire (V1)[18]. Pourtant, des épisodes d'expérience visuelle intense ne semblent pas affecter la densité des circuits associés aux œstrogènes sous leur distribution, et les auteurs signalent que des souris adultes ayant été élevés depuis leur naissance dans le noir présentent une distribution normale de l'aromatase et des récepteurs aux œstrogènes dans le cortex visuel primaire (V1), ce qui signifierait que la mise en place et l'entretien de ces circuits neuronaux liés aux œstrogènes produits localement sont indépendants de l'expérience visuelle[18]
Notes et références
Notes
- Quand on désire distinguer précisément le champ visuel (plan) du champ rétinien (convexe), on parle pour le premier de relation visuotopique, mais en général, les cartes rétinotopiques sont faites à partir du champ visuel.
- Un enregistrement laissé sur un support photographique par un objet contenant un produit radioactif.
- ou angle visuel θ exprimé en degré (voir champ visuel), la distance à la fovéa r est proportionnelle.
- Au point z du champ visuel z=ρ ei α on fait correspondre par le logarithme complexe le point log(z)=log(ρ) + i α du cortex. Pour éviter la singularité du logarithme complexe au point z=0, on prend l'application monopôle w=k log(z+a) qui déplace la singularité en z=-a. Une meilleure approximation, tenant compte de la forme de la frontière antérieure de l'aire V1, est obtenue avec l'application dipôle w=log(z+a)-log(z+b), avec par exemple a=1,5 et b=170, voir Balasubramanian et als, 2002. D'autres améliorations ont été apportées à ces modèles qui ne nous intéressent pas ici.
- Segmentation de l'image en 3 classes : matière blanche, matière grise, liquide céphalo-rachidien.
- abr. de Middle Temporal, temporal moyen.
Références
- (en) Brian A. Wandell, Jonathan Winawer, « Imaging retinotopic maps in the human brain », Vision Research, vol. 51, , p. 718-737.
- (en) Martin I. Sereno, Colin T. McDonald, and John M. Allman, « Analysis of retinotopic maps in extrastriate cortex », Cerebral Cortex, vol. 4, , p. 601-620..
- (en) Wandell BA, Dumoulin SO, Brewer AA, « Visual field maps in human cortex », Neuron, vol. 56, no 2, , p. 366-83.
- (en) Rudolf Nieuwenhuys, Jan Voogd, Chr. van Huijzen, The Human Central Nervous System, Springer, 1978, 2008, 967 p..
- Jan Warnking, Délinéation des aires visuelles rétinotopiques chez l’homme par IRM fonctionnelle, Grenoble, thèse de physique, Université Joseph Fourier, .
- (en) R B Tootell, E Switkes, M S Silverman, S L Hamilton, « Functional Anatomy of Macaque Striate Cortex. II Retinotopic organization », Journal of Neuroscience, vol. 8, no 5, , p. 1531-1568.
- (en) Kalanit Grill-Spector et Rafael Malach, « The human visual cortex », Annu Rev Neurosci, vol. 27, .
- (en) Mukund Balasubramanian, Jonathan Polimeni, Eric L. Schwartz, « The V1-V2-V3 complex: quasiconformal dipole maps in primate striate and extra-striate cortex », Neural Networks, vol. 15, no 10, , p. 1157-1163.
- Wandell BA, Brewer AA, Dougherty RF., « Visual field map clusters in human cortex. », Philos Trans R Soc Lond B Biol Sci., vol. 360, , p. 693-707.
- S. A. Engel, D. E. Rumelhart, B. A. Wandell, A. T. Lee, G.H. Glover, , E. J. Chichilnisky, and M. N. Shadlen., « fMRI of human visual cortex », Nature, vol. 369, , p. 525.
- (en) Stephen A. Engel, Gary H. Glover, & Brian A. Wandell, « Retinotopic organization in human visual cortex and the spatial precision of functional MRI », Cereb Cortex, vol. 2, .
- (en) M.I. Sereno, A.M. Dale, J.B. Reppas, K.K. Kwong, J.W. Belliveau, T.J. Brady, B.R. Rosen, and R.B.H. Tootell, « Borders of multiple visual areas in humans revealed by functional MRI », Science, vol. 268, , p. 889-893.
- (en) Warnking J, Dojat M, Guérin-Dugué A, Delon-Martin C, Olympieff S, Richard N, Chéhikian A, Segebarth C., « fMRI retinotopic mapping--step by step », NeuroImage, vol. 17, no 4, , p. 1665-83.
- Flor Vasseur, Un cadre unique de représentation pour des études visuelles de groupe sur la base de cartes rétinotopiques acquises en IRMf à 3 T, Thèse, Université Joseph Fourier, Grenoble 1, .
- Jonas Larsson and David J. Heeger, « Two Retinotopic Visual Areas in Human Lateral Occipital Cortex », The Journal of Neuroscience, vol. 26, no 51, , p. 13128-13142.
- Andreas Bartels and Semir Zeki, « The architecture of the colour centre in the human visual brain: new results and a review », Eur. J. Neurosci, vol. 12, no 1, , p. 172-193.
- Saygin, A.P. and M.I. Sereno, « Retinotopy and attention in human occipital, temporal, parietal, and frontal cortex », Cerebral Cortex, vol. 18, , p. 2158-2168.
- Jeong JK, Tremere LA, Burrows K, Majewska AK, Pinaud R (2011), The mouse primary visual cortex is a site of production and sensitivity to estrogens ; PLoS One. 2011; 6(5):e20400. Epub 2011 May 24 (résumé)
- ER signifie « Estrogen Receptors ».