Origine du langage
L’origine du langage humain a toujours suscité l’intérêt des penseurs. De nombreux mythes tendent à donner aux langues une origine surnaturelle. La Bible explique ainsi la multiplicité des langues par le mythe de la Tour de Babel, selon lequel la langue unique des origines aurait été divisée en une multitude de langues pour apporter la discorde entre les hommes et les empêcher de se concerter en vue d'une action commune.
Pour éviter les querelles stériles et les thèses farfelues, la Société de linguistique de Paris avait en 1865 informé ses membres dans ses règlements qu'elle ne recevrait plus « aucune communication concernant […] l'origine du langage »[1]. Aujourd'hui, le sujet est étudié scientifiquement dans le cadre de plusieurs disciplines, notamment en paléontologie, en psychologie, en biologie moléculaire et en linguistique historique.
L’histoire de l’évolution du langage humain est longue et étroitement liée au cerveau, mais ce qui fait que le cerveau humain soit le seul à être adapté au langage n’est pas clair. Les régions cérébrales impliquées dans le langage chez les humains ont leurs analogues chez les singes et les grands singes, pourtant ces derniers n’utilisent pas le langage. Il pourrait aussi y avoir une composante génétique : des mutations du gène de la protéine Forkhead-P2 empêchent les humains concernés de construire des phrases complètes[2].
Selon John Maynard Smith et Eörs Szathmáry, le langage apparaît comme la plus récente des huit transitions majeures dans l’évolution de la vie, permettant l’émergence des sociétés humaines. Plus généralement, l’évolution de la communication permet l’émergence des différents niveaux de sociabilité.
Mythe biblique

Ce dessin se conforme à la tradition picturale : la punition divine frappe les humains pour leur ambition sans bornes à édifier Babel ; le motif mythologique est comparable à Prométhée, qui provoque la colère divine en dérobant le secret du feu.
Le mythe de la Tour de Babel vise à rendre compte de la multiplicité des langues :
« Toute la terre avait une seule langue et les mêmes mots. Comme ils étaient partis de l’orient, ils trouvèrent une plaine au pays de Chmunter, et ils y habitèrent. Ils se dirent l'un à l'autre : « Allons ! Faisons des briques, et cuisons-les au feu. » Et la brique leur servit de pierre, et le bitume leur servit de ciment. Ils dirent encore : « Allons ! Bâtissons-nous une ville et une tour dont le sommet touche au ciel, et faisons-nous un nom, afin que nous ne soyons pas dispersés sur la face de toute la terre. »
L'Éternel descendit pour voir la ville et la tour que bâtissaient les fils des hommes. Et l'Éternel dit : « Voici, ils forment un seul peuple et ont tous une même langue, et c’est là ce qu'ils ont entrepris ; maintenant rien ne les empêcherait de faire tout ce qu'ils auraient projeté. Allons ! descendons, et là confondons leur langage, afin qu’ils n’entendent plus la langue, les uns des autres. » Et l’Éternel les dispersa loin de là sur la face de toute la terre ; et ils cessèrent de bâtir la Ville. C’est pourquoi on l’appela du nom de Babel, car c’est là que l’Éternel confondit le langage de toute la terre, et c’est de là que l’Éternel les dispersa sur la face de toute la terre. »
— Traduction courante de la Genèse, au chapitre 11, versets 1 à 9
Approche scientifique : objets et méthodes d'étude
Le langage est très largement considéré comme strictement humain. C’est pourquoi l’approche comparative est privilégiée. Elle repose sur la recherche des points communs et différences entre les systèmes de communications non humains et le langage. L’absence de caractéristiques particulières chez les « langages non humains » est souvent retenue comme critère de singularité du langage humain. Or cette absence résulte peut-être d’une incapacité à détecter cette caractéristique chez le sujet d’étude. De plus, il n’est pas exclu que le fait d’étudier le langage en utilisant le langage introduise un biais.
Cependant, la comparaison de données peut aussi élargir la discussion (cf. le larynx descendu).
Aspects anthropologiques
À partir de reconstructions de contours de pharynx, certains scientifiques américains, dont Lieberman (en) et Crelin (en), ont rejeté l'hypothèse de l'existence d'un langage articulé chez les prédécesseurs d’Homo sapiens[3]. Leur théorie, fondée sur la nécessité d'un larynx en position basse[4] pour produire un langage articulé, est depuis infirmée : de nombreux mammifères produisent dans leurs signaux sonores des formants (ou fréquences de résonance) en mobilisant la partie haute de l'espace pharyngal ; des modélisations informatiques montrent que, même avec un larynx en position haute, les articulateurs (en) (langue, voile du palais, dents et lèvres) du tractus vocal ont un degré de liberté tel qu'il est possible de les coordonner afin de produire une large gamme de sons articulés ; certains fossiles de Néandertaliens montrent des larynx en position basse[5].
Selon Perreault et Mathew[6], le langage humain serait apparu il y a entre 350 000 et 150 000 ans chez Homo sapiens. Cette datation est estimée par la vitesse d’accumulation de phonèmes. L’étude des os, de la musculature cranio-faciale et des phénomènes culturels (sépultures) donne aussi des informations sur le moment de son apparition.
Aspects linguistiques et philosophiques
Source[7].
Le langage serait le propre de l'homme. Le langage s’appuie sur un vocabulaire dont les éléments en nombre fini peuvent se combiner d’une infinité de manières. De plus, ce vocabulaire n’est pas figé : tandis que certains des mots apparaissent, d’autres disparaissent ou voient leur sens changer. Ces deux caractéristiques du langage feraient de lui une faculté uniquement humaine. Le linguiste Emile Benveniste observe et interprète la danse des abeilles. Il en déduit que la différence entre la danse des abeilles et le langage humain réside dans le fait que la danse des abeilles n’est pas spontanée mais consiste en la réponse à un stimulus environnemental. De plus, la danse d’une abeille conduit les autres abeilles à avoir un comportement de butinage différent et non une réponse dansée. En outre il n’y a pas d’échange.
Enfin, le langage est indispensable au développement des caractéristiques essentielles de l’humanité (en particulier la pensée, la conscience réfléchie). On peut en effet penser aux enfants sauvages qui sont privés d’un développement intellectuel normal.
Théories de l'origine du langage
Origine gestuelle
La parole et la vocalisation auraient évolué à partir de la gestuelle chez l’être humain[8]. Cette hypothèse est soutenue par plusieurs arguments :
- les zones cérébrales qui contrôlent la production et le traitement du langage (aires de Broca et de Wernicke notamment), la main et le visage sont proches et interconnectées ;
- des neurones miroirs seraient impliqués dans la connexion de ces zones ;
- il y aurait un lien entre la latéralisation du langage et la préférence manuelle. Ce lien a récemment été remis en cause par le groupe d’imagerie neurofonctionnelle de Bordeaux[9] ;
- dans les langues des signes, ce sont les mêmes aires qui sont mises en jeu ;
- lorsque l’on parle, on utilise aussi ses mains pour s’exprimer (par exemple lors d’un discours) ;
- pour s’exprimer, les enfants humains montrent du doigt.
Les chimpanzés qui produisent des sons visant à attirer le rassemblement montrent une activité dans des régions cérébrales très similaires à la région de Broca chez l’être humain[10] - [11]. Même les mouvements des mains et de la bouche sans émission de sons produisent des schémas d’activité cérébrale très similaires dans l’aire de Broca chez les singes et les humains[12]. Quand les singes voient d’autres singes en mouvement, les neurones miroirs de l’aire de Broca homologue s’activent. Des groupes de neurones miroirs sont spécialisés pour ne répondre qu’à un seul type d’action observée, et l’on pense actuellement que cela pourrait être une origine de l’évolution des neurones qui sont destinés au traitement et à la production de la parole[13].
Le langage le plus ancien était strictement oral ; la lecture et l’écriture ne sont arrivées que bien plus tardivement[14].
Grammaire universelle
L’hypothèse du bioprogramme linguistique suggère que les humains ont une structure cognitive grammaticale innée leur permettant de développer et de comprendre le langage. Selon cette théorie, ce système est ancré dans la génétique humaine et étaye la grammaire de base de tous les langages[12]. Certaines preuves suggèrent qu’au moins une partie de nos capacités linguistiques serait contrôlée génétiquement. Des mutations du gène de la protéine Forkhead-P2 empêchent certains individus de transformer des mots et des sons en phrases[2]. Cependant, ces gènes sont présents dans le cœur, les poumons et le cerveau, et leur rôle n’est pas entièrement établi[2].
Il est possible que la capacité grammaticale des humains ait évolué depuis des comportements non sémantiques comme le chant[15]. Les oiseaux ont la capacité de produire, traiter et apprendre des sons complexes, mais les unités d’un chant d’oiseaux, lorsqu’elles sont supprimées du sens large et du contexte de ce chant dans son ensemble, n’ont pas de sens inhérent. Des hominidés primitifs ont peut-être développé des capacités à but non sémantique similaires, qui se seraient transformés ensuite en langage symbolique[16].
Ciment social
En 1996, dans son ouvrage Grooming, Gossip and the Evolution of Language (en), le professeur de psychologie évolutionniste Robin Dunbar formule une hypothèse sur l'évolution du langage chez les humains. De la même manière que les primates non humains maintiennent leurs liens sociaux à l'aide de l'épouillage, les humains maintiennent un lien social par la conversation et le langage. Dunbar suggère que les humains, en évoluant, ont privilégié le langage à l'épouillage car la taille de leurs groupes était devenue trop importante. Le langage comme ciment social aurait pris moins de temps et permis à l'individu de faire plusieurs choses à la fois[17]. C'est la théorie du « Gossip ». Gossip en anglais veut dire « potin ». L'idée étant que même les petites conversations anecdotiques qui ne transmettent pas d'informations primordiales nécessaires à la survie sont importantes dans ce qu'elles créent du lien social.
Proto-langues
Il est difficile d’envisager que le langage soit apparu d’un bloc. Certains soutiennent que le langage suit une évolution graduelle depuis un langage primitif. Les langues ne se fossilisant pas, il faut donc faire appel à des méthodes linguistiques pour reconstruire ce langage primitif et les langages intermédiaires (proto-langues). Deux hypothèses s'affrontent depuis le XIXe siècle, celles de la monogénèse et de la polygénèse des langues.
Les travaux récents en anthropologie, en archéologie, en génétique et en linguistique suggèrent l'hypothèse d'une langue originelle commune[18]. En se basant sur des ressemblances lexicales, les linguistes avaient déjà pu établir depuis plus d'un siècle l'arbre généalogique approximatif de la grande famille de langues issues de l'indo-européen. En 2003, Russell D. Gray et Quentin Atkinson ont proposé d'appliquer à 2 449 termes provenant de 87 langues de cette famille de langues une méthode phylogénétique informatisée comme celle qu'utilise la biologie moléculaire pour construire des arbres généalogiques à partir de l'ADN[19]. Cette méthode prend comme unité de base non pas les lexèmes mais les phonèmes présents dans les différentes langues. Considérant que le nombre de phonèmes d'une langue augmente en fonction du nombre de locuteurs qui la parlent, mais diminue lorsqu'un sous-groupe émigre loin de la famille mère[20], ils ont ainsi pu établir que l'expansion des langues indo-européennes correspond au développement de l'agriculture à partir du plateau anatolien entre 7 800 et 9 800 ans avant notre époque[19]. En 2011, Atkinson applique son modèle à un plus grand nombre de langues. Dans une base de 504 langues, il observe que certaines langues africaines comptent plus de 100 phonèmes, alors que le hawaïen, qui est la langue la plus éloignée du berceau africain, n'en compte que 13, contre 45 en anglais et 36 en français. Cette diminution de la diversité des phonèmes en fonction de la distance, comparable à celle qu'on observe dans les caractéristiques génétiques, impliquerait selon Atkinson que le langage humain aurait pris naissance dans le sud-ouest africain il y a entre 50 000 ans et 100 000 ans[20] - [21].
Théorie du bébé à terre
Selon la théorie de « bébé à terre » de Dean Falk, les interactions vocales entre les premières mères hominidées et leurs nourrissons auraient déclenché une séquence d'événements qui auraient mené, éventuellement, aux premiers mots de nos ancêtres[22]. L'idée fondamentale est que les premières mères humaines, contrairement à leurs homologues chez les autres primates, ne pouvaient pas se déplacer et se nourrir avec leurs nourrissons accrochés sur leur dos. La perte de fourrure a laissé les nourrissons sans aucun moyen de s'accrocher. Souvent, les mères ont alors dû mettre leurs bébés sur le sol. En conséquence, ces bébés devaient être rassurés. Les mères aurait répondu aux pleurs des bébés en développant un système communicatif dirigé par leur nourrisson englobant des expressions faciales, un langage corporel, le toucher, les tapotements et caresses, les rires, les chatouilles et les appels émotionnellement expressifs. Selon Dean Falk, la langue aurait pu apparaître de cette manière[22].
Adaptations neurobiologiques favorables au langage
Aires de Broca et de Wernicke
Ce sont les régions du cerveau où le langage se situe – tout depuis la parole à la lecture et l’écriture[23]. Le langage est lui-même fondé sur des symboles utilisés pour représenter des concepts dans le monde, et ce système semble être stocké dans ces zones du cerveau. Les régions du langage dans les cerveaux humains ressemblent fortement aux régions similaires chez d’autres primates, même si les humains sont les seuls capables d’utiliser le langage[14].
Les structures cérébrales des chimpanzés sont très similaires à celles des humains. Ces dernières possèdent toutes deux des homologues des aires de Broca et de Wernicke qui sont impliquées dans la communication. L’aire de Broca est en grande partie utilisée pour planifier et produire des sons chez les chimpanzés comme chez les humains. L’aire de Wernicke semble être l’endroit où les représentations linguistiques et les symboles sont affectés à des concepts spécifiques. Cette fonctionnalité est présente à la fois chez les chimpanzés et chez les humains ; l’aire de Wernicke du chimpanzé est bien plus similaire à son homologue humain que l’aire de Broca ne l’est au sien, ce qui suggère que l’aire de Wernicke évolue depuis plus longtemps que celle de Broca.
Neurones moteurs
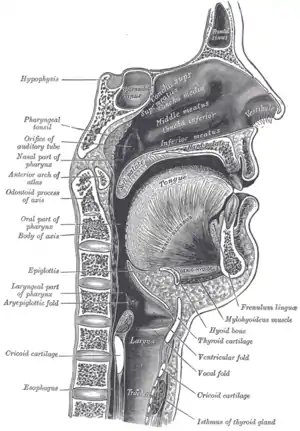
Afin de parler, le système respiratoire doit être volontairement détourné afin de produire des sons vocaux[14]. Ceci permet aux mécanismes respiratoires d’être temporairement désactivés afin de produire du chant ou de la parole. Le canal vocal a évolué pour être mieux adapté à la parole, avec un larynx plus bas, une inclinaison à 90° de la trachée, et une large langue arrondie[24]. Les neurones moteurs chez les oiseaux et les humains contournent les mécanismes de l’inconscient dans le tronc cérébral pour donner au cerveau un contrôle direct du larynx[16].
Gène FOXP2
Source[8].
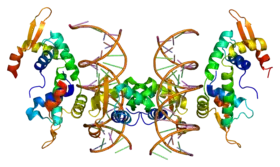
Ce gène code une protéine appartenant à la famille des régulateurs de transcriptions. Il a été mis en évidence chez la famille KE dont certains membres présentent des difficultés d’articulation et une incapacité à acquérir certaines règles de grammaire.
La protéine est impliquée dans le développement de structures cérébrales interconnectées et associées à l’activité motrice. C’est une protéine hautement conservée chez les oiseaux chanteurs et les mammifères.
Enfin, la dernière mutation importante aurait eu lieu il y a 200 000 à 100 000 ans chez l’ancêtre commun d’Homo sapiens et de l'Homme de Néandertal : cela coïncide avec l’estimation de l’apparition du langage articulé.
Le larynx descendu : caractère sexuel secondaire ou organe acoustique ?
Le larynx chez l’humain adulte se trouve plus bas dans la gorge par rapport aux autres mammifères. Cela implique que l’homme ne peut pas respirer et déglutir en même temps.
Ces chercheurs ont vu dans cette contre-adaptation apparente un avantage pour la communication vocale. En effet, un larynx descendu présente des propriétés acoustiques supérieures, permettant une augmentation de la diversité des sons de voyelles[25].
En 2001, Fitch et Reby[26] examinent aussi des données comparées. Ils remarquent que dans d’autres espèces de mammifères, les pressions de sélection actuelles favorisent un larynx descendu. Chez la biche rouge, le mâle possède un larynx descendu mais pas la femelle. On parle de dimorphisme d’un caractère sexuel secondaire. Un tel larynx permet de produire une vocalisation exagérément sonore par rapport à la taille de l’individu. Deux pressions de sélection peuvent alors entrer en jeu :
- compétition pour une partenaire et préférence des femelles : la vocalisation exagérée apporterait un avantage dans la compétition directe entre les mâles reproducteurs (intimidation), car elle serait un signal honnête de bons gènes, ce qui lui donnerait la préférence des femelles ;
- prédation : ce caractère serait un signal honnête de non profitabilité de la proie ; en effet, la vocalisation produite suggère que l’animal est en bonne condition physique et donc difficile à capturer.
Chez l’homme, il existe aussi un dimorphisme : l’homme et la femme ont tous deux un larynx descendu, mais celui de l’homme subit des modifications à la puberté (seconde descente et apparition de la pomme d’Adam).
Cette analyse comparative relativise la théorie du larynx comme adaptation au langage humain.
Apparition du langage et moteurs de cette évolution
Point de vue paléontologique

C’est l'Homo habilis, il y a plus de deux millions d’années, qui pourrait être le plus ancien préhumain à avoir employé un langage articulé[27], ce qui ne signifie pas pour autant que cet hominidé ait usé d’un langage comparable au nôtre. On suppose la préexistence d’une proto-langue chantée par l'Homme de Néandertal (the singing Neanderthal[28]) qui, au niveau actuel des connaissances, ne possédait pas de syntaxe.
La morphologie du crâne d’Homo habilis, marquée par l’apparition d’une flexure antéropostérieure jusqu’alors absente chez les Australopithèques, conduisait en effet à l’expansion des zones cérébrales impliquées aujourd’hui dans le langage articulé. Par ailleurs, le redressement du crâne chez l'Homo habilis abaissait les voies aériennes supérieures, pharynx et larynx (d’où l’apparition d’une pomme d'Adam), ce qui était une condition nécessaire pour pouvoir moduler la vocalisation et augmentait la hauteur de la voûte du palais, permettant à la langue d’articuler une plus large gamme de sons. Apparues avec le genre Homo, ces caractéristiques allaient se renforcer nettement par la suite, notamment chez l’espèce Homo erectus : au-delà de la bipédie, il se serait agi en fait d’une adaptation à la course à pied pour permettre de mieux contrôler son souffle, en même temps que l’élargissement du thorax pour renforcer la respiration et, sans doute, la perte de la majeure partie des poils pour réguler la température corporelle pendant l’effort.
Il est possible que ces capacités physiologiques aient permis l’essor d’une communication orale à la complexité croissante, permettant aux populations de l’Homo habilis d’organiser leurs communautés en régulant leurs activités quotidiennes. L'Homo habilis est en effet le premier hominidé pour lequel il y ait évidence d'une organisation sociale structurée (campements, outils, habitats et sans aucun doute spécialisation des individus).
Ultérieurement, il faut noter que l’augmentation de la masse de l’encéphale[29], continue de l’Homo erectus à l’Homo sapiens, a été un point-clé dans la maturation du langage. Lors du passage à l'Homo sapiens sont apparues des aires de Broca sur une circonvolution frontale gauche, et de Wernicke sur une circonvolution temporale gauche qui ont suivi la mutation génétique d’au moins un gène dominant FOXP2[29], dit de la parole, qui a donné la capacité de l’homme de passer des mots à la syntaxe (ce facteur n’est pas suffisant en lui-même, car il existe chez d’autres espèces sans donner naissance à la parole) ; il faut mentionner que ce(s) gène(s) serai(en)t à l’origine de la maturation de ces deux zones : l'aire de Broca et l’aire de Wernicke[30]. Cependant, dans une étude publiée en 2014 dans la revue Brain[31], le neurochirurgien et neuroscientifique Hugues Duffau montre que « l'aire de Broca n'est pas l'aire de la parole » et que les fonctions langagières ne sont pas tant localisées dans une aire précise que dépendantes de connexions neuronales en reconfiguration constante[32].
Prédominent actuellement deux scénarios d’apparition de l’Homo sapiens, le scénario « Out of Africa » et un scénario pluricentripète (polygenèse). Certaines recherches en paléolinguistique ont identifié un fonds de vingt-sept mots communs à la racine de toutes les langues terrestres écrites au début du XXIe siècle[33], ce qui pousse à favoriser le scénario « Out of Africa » (monogenèse). En effet, plusieurs sources n’auraient pas eu de raison d’adopter la même protolangue de départ[34]. Les divers rameaux du moderne Homo sapiens qui sont partis d'Afrique il y a cent mille ans partageaient déjà une même fonction langagière, bien avant l'apparition de l'Homme de Cro-Magnon[35].
Enfin, l’Homo sapiens a dominé le monde, soit du fait de l’hypothèse productiviste[36], soit du fait de l’hypothèse sociologique[37].
Le langage pourrait avoir de multiples origines, les aires cérébrales du langages étant proches de celles mobilisées pour le travail manuel de précision (ce qui induit un codéveloppement des facultés langagières et manufacturières du genre humain), tandis que l’articulation de sons est par ailleurs souvent corrélée de façon réflexe à des mouvements du corps (à l’effort ou sous l’effet de la surprise, notamment) ; la perception de ces sons pouvait en retour être affinée par le développement du cerveau humain, libéré par la bipédie des limitations d’encombrement et de poids, puisque désormais littéralement « posé » sur la colonne vertébrale, ce qui permettait de charger de sens ces sons nouveaux que la nouvelle morphologie crânienne d’Homo habilis permettait de produire. D’un point de vue neurologique, le développement du langage semble provenir des mécanismes de reconnaissance du comportement, de la gestuelle et de l’action d’autrui.
Point de vue génétique
La substitution d’un seul des 715 acides aminés du gène FoxP2 entraîne de sérieuses pathologies affectant la phonation et, plus généralement, la forme du larynx. Ce gène est, en raison même du caractère sensible de ses mutations, demeuré d’une remarquable stabilité au cours de l’évolution, la séquence de la protéine humaine ne différant que pour deux acides aminés (sur 715) de celle des chimpanzés, des gorilles et des macaques rhésus, et pour un acide aminé supplémentaire avec la souris[38]. La mutation du gène FoxP2 intervenue chez Homo sapiens il y a cent à deux cent mille ans a donc certainement dû être déterminante, mais s’est inscrite dans une dynamique d’évolution commencée plusieurs millions d’années auparavant[39].
Huitième transition majeure
En 1997, John Maynard Smith et Eörs Szathmáry ont publié un livre s’intitulant The Major Transitions in Evolution. Ce livre va profondément influencer par la suite les travaux de biologie évolutive.
Ces deux chercheurs ont identifié huit transitions majeures dans l’évolution de la vie. L’apparition du langage est la plus récente des huit et consiste en un passage des sociétés de primates vers les sociétés humaines. Pour eux, le langage est uniquement présent chez les humains puisqu’il est à la base de leurs sociétés. Il s’agit d’un nouveau mécanisme de transmission d’informations permettant une transmission socioculturelle de ces informations, non limitée par l’hérédité.
Modèles FLB (Faculty of Language in the Broad sense) et FLN (Faculty of Language in the Narrow sense)
Un article de Marc Hauser, Noam Chomsky (linguiste et philosophe) et W. Tecumseh Fitch paru en 2002[40] a produit une énorme discussion en linguistique, en sciences cognitives et en théorie de l’évolution sur le langage. Dans l’article, les auteurs abordent la question de ce qu’il y a de commun et de différent entre l’espèce humaine et les autres espèces, ainsi que la question de l’évolution du langage. Ces chercheurs ont proposé un classement des différentes facultés de langage pour faciliter les discussions sur l’évolution. Ils ont créé les termes de FLB et FLN.
Division de la faculté de langage
Le FLB correspond à une faculté de langage au sens large. Cela inclut les propriétés du langage des hommes partagées avec les systèmes de communication des êtres « non humains » (les animaux). Il est composé d’un système sensorimoteur qui fait intervenir le système moteur et le système sensoriel et d’un système conceptuel-intentionnel permettant de représenter ce que l’on a l’intention de faire. Le FLB regrouperait les êtres possédant des systèmes biologiques qui sont nécessaires mais pas suffisants au langage humain (mémoire, respiration, digestion, circulation).
Le FLB inclut également le FLN qui est la faculté du langage au sens étroit. Cette faculté est basée sur le mécanisme de récursivité qui est la possibilité de créer une infinité d’expressions (idées, sentiments…) à partir d’un nombre fini d’éléments (les mots). Ces expressions sont ensuite transmises aux deux systèmes sensorimoteur et conceptuel-intentionnel. Le langage humain est doté d’une telle faculté.
Les trois chercheurs ont mis en avant trois hypothèses d'évolution
- Hypothèse 1 : le FLB est strictement homologue à la communication animale. Cela sous-entend que les composants fonctionnels permettant le langage des humains seraient identiques à ceux des systèmes de communication des autres espèces.
- Hypothèse 2 : le FLB est une adaptation dérivée, spécifiquement humaine, pour le langage. C’est la sélection naturelle qui a joué un rôle dans la formation du FLB, ce processus étant sans parallèle chez les animaux non humains.
- Hypothèse 3 : seul le FLN est uniquement humain. Le FLN serait une évolution récente spécifiquement humaine. Les systèmes sensorimoteur et conceptuel-intentionnel sont alors combinés avec des composantes socioculturelles et communicatives.
Marc Hauser, Noam Chomsky et W. Tecumseh Fitch suggèrent que l’hypothèse 3 serait la plus plausible. Ils pensent que, si les hypothèses 1 ou 2 étaient valables, il faudrait qu’une série de mutations graduelles du FLB puissent mener à terme à une capacité du langage humain basé sur la récursivité. Des modifications mineures de ce système fondamental sembleraient insuffisantes pour y parvenir.
Apports de l'expérimentation
De nombreux résultats d’expériences d’autres chercheurs (neurologues, linguistes, psychologues, biologistes…) ont été étudiés dans leurs articles pour justifier leurs points de vue. Ces études portent notamment sur les systèmes sensorimoteur et conceptuel sensoriel qu’ont les animaux et les humains en commun mais aussi sur le principe de récursivité.
- chez les oiseaux chanteurs : la faculté à produire des signaux complexes comme chez les humains, qui est une faculté propre au FLB, a été retrouvée chez de nombreux animaux ayant la capacité à l’apprentissage vocal comme l’oiseau chanteur[41]. Il a été montré que certaines zones du cerveau d’un être humain pourraient être similaires à celles des oiseaux chanteurs (notamment le canari). Des problèmes et déficits de langages similaires à ceux des humains ont été repérés chez ces oiseaux lorsque a lieu une lésion dans une zone identique du cerveau. Il a été aussi montré que l’expression de gènes, certes différents dans la structure, a cependant un mécanisme d’activation identique dans différentes zones du cerveau de ces oiseaux et des humains via des tests d’ouïe et de parole. Que ce soit chez les humains ou les oiseaux, certains gènes sont activés ou non lorsqu’ils entendent un congénère de la même espèce s’exprimer. D’autres gènes sont activés dans les mêmes régions du cerveau (principalement le cortex moteur du visage) lorsqu’ils émettent des sons ;

- chez les tamarins : les tamarins sont-ils capables d’apprendre des phrases à structure complexe basée sur le principe de récursivité ? Les tamarins sont conditionnés la veille du test. On leur fait écouter des suites de séquences de syllabes respectant les règles d'un système soit à phrase simple (à état fini : ABAB), soit à phrase structurée (AAABBB) basée sur le principe de récursivité. Le système à phrase simple est basé sur une alternance d’éléments A et B. Le système à phrase structurée consiste en une chaîne d’éléments A et B dans laquelle chaque élément A et B peut être séparé par un élément intervenant. Il y a une intégration d’unités de représentation au sein de cette chaîne hiérarchisée. Le test consiste à faire écouter des nouvelles séquences de syllabes respectant ou non les règles d’un des deux systèmes. Les tamarins ayant été conditionnés pour le système à phrase simple réagissent quand la suite de syllabes écoutée ne correspond pas aux règles apprises (9 sur 10 tamarins: en regardant pendant un long moment le speaker; quand ils ne réagissent pas ils ne le regardent que très brièvement). Les tamarins ayant été conditionnés pour le système à phrase complexe réagissent moins quand une suite de syllabes ne respecte pas les règles du système. Les tamarins ont donc échoué au test. Cette étude montre que les tamarins pourraient ne pas posséder le principe de récursivité, mais cela ne prouve pas forcément que tous les êtres non humains ne le possèdent pas.
Critiques et limites du modèle FLN/FLB
Cette expérience est assez critiquée. Il a été montré que des étourneaux ont pu réussir ce test contrairement aux singes[42]. Le test a été réalisé avec des séries de huit gazouillis et huit hochets différents. Ces oiseaux ont été formés pour picorer leur nourriture correspondant à une récompense s'ils entendaient une série respectant le système de grammaire auquel ils ont été formés. S'ils picorent à tort, la récompense leur est interdite. Seuls deux étourneaux sur onze ont échoué au test. Dans ce test, il existe aussi de nombreux échecs chez les humains, qui sont pourtant censés être capables de récursivité.
D’autres chercheurs, comme Ray Jackendoff et Steven Pinker[43] ont aussi montré que la récursivité, bien qu’absente des autres systèmes de communication animale, se trouve dans la cognition visuelle. Or, on ne sait pas si les animaux « non humains » sont capables de récursivité visuelle, ce qui implique qu’elle ne peut être le seul développement évolutif qui permette le langage aux humains.
Ils contestent également principalement la 2e et la 3e hypothèse. Pinker et Jackendoff considèrent que leur caractérisation de la faculté du langage restreint (FLN) est problématique et ce, pour plusieurs raisons : tout d’abord, ils mettent en place une dichotomie des capacités cognitives : celles qui sont entièrement spécifiques aux humains et les capacités « non linguistiques » ou « non humaines », omettant ainsi les capacités cognitives qui auraient pu être en grande partie modifiées pendant l’évolution humaine. Fitch, Hauser et Chomsky omettent les traits qui sont des adaptations, ne considérant que le trait dans sa version ancestrale et actuelle. De plus, leur comparaison humain/animaux ne permet pas de faire une différence entre les similarités dues aux fonctions communes et les similarités dues à l’héritage d’un ancêtre commun récent. L’affirmation qu’un trait est « spécifique au langage » ou « spécifique aux humains » peut être interprétée de deux manières. Tout d’abord, le « tout ou rien ». Le trait est spécifique aux humains et il n’y a rien de similaire dans le règne animal. Il apparaît, à partir de rien, dans l’évolution. Cela peut être interprété avec des termes plus nuancés : le trait a été modifié au cours de l’évolution humaine à un tel degré qu’il en est différent de manière significative de son ancêtre évolutif (a priori, cela serait le résultat d’une adaptation à une nouvelle fonction pour laquelle le trait a été sélectionné). Fitch, Hauser et Chomsky attribuent au FLB n’importe quel trait retrouvé dans le monde animal tel que l’imitation vocale chez les oiseaux chanteurs qui a évolué indépendamment du langage humain. La comparaison entre « trait partagé avec animaux non humains » et « évolué récemment » n’a de sens que si les animaux en question ont un ancêtre commun récent (tels que les chimpanzés). S’il s’agit de moineaux ou de dauphins, alors le trait (comme l’apprentissage vocal), peut être partagé avec l’animal et évolué récemment.
Une dynamique à trois composantes
Selon Simon Kirby[44], le langage est un nouveau système de transmission d’information. Le langage n’est pas complètement inné, il doit être en très grande partie appris. La meilleure façon d’apprendre est d’observer son prochain (qui lui-même a appris en observant…) lorsqu’il utilise le langage, puis de le copier. Ce principe est appelé langage itératif.
Le langage évolue donc au fil du temps avec les observations et les interprétations du comportement de chaque individu dans l’utilisation de la langue sur chaque individu dans une société.
Le langage serait lui-même un mécanisme d’évolution comprenant trois systèmes dynamiques qui prennent part à la transmission d’information : l’évolution biologique, la transmission culturelle et l’apprentissage individuel qui a un plus faible impact dans ce mécanisme.
Ces trois dynamiques interagissent entre elles de façon cyclique et complexe. Les mécanismes d’apprentissage des langues font partie de notre héritage biologique et sont donc soumis à l’évolution biologique. Cette évolution a un impact sur la dynamique culturelle de transmission linguistique par l’apprentissage réitéré (apprentissage itératif). Les nouvelles structures de langages issus de la transmission culturelle ont alors en partie un effet sur la fitness des individus les possédant et donc un impact sur la trajectoire de leur évolution biologique dans les mécanismes d’apprentissage du langage…
Mise en évidence d'un paradoxe et système de communication déductif
Orrigi et Sperber[45] ont constaté que jusqu’à présent les différentes approches considèrent la faculté du langage comme une adaptation mais construisent leur modèle afin d’expliquer l’usage du langage ce qui n’est qu’une partie de cette faculté.
Paradoxe
Quand on considère la faculté du langage comme une adaptation biologique, on doit envisager une de ses fonctions proximales permettant l’ensemble des autres fonctions (compétence linguistique, communication verbale). Il s’agit de la fonction d’acquisition du langage. Or une adaptation est maintenue uniquement si elle apparaît dans un environnement dans lequel elle a une valeur adaptative. Dans cet environnement particulier, le langage doit préexister. La faculté de langage et le langage sont donc l’une pour l’autre une précondition.
Considérons un mutant qui a la faculté d’échanger avec ses semblables. Il en tire un bénéfice uniquement lorsqu’un autre individu présente cette aptitude. Habituellement, les mutants présentant une innovation adaptative bénéficient d’un avantage sélectif quand ils sont en petit nombre.
Trois hypothèses tentent de résoudre ce problème chez les langages non humains basés sur un code inné ; on accepte comme prérequis que le langage apporte un bénéfice :
- l’hypothèse du trait neutre (Sober, 1984[46]) : le trait « langage » n’apporte initialement pas de bénéfices, mais pas de coût non plus. Il peut se répandre dans la population, car il n’est pas contre sélectionné. Quand un nombre limite d’individus portant ce trait apparaissent et interagissent, ils en tirent bénéfices. Le trait est alors sélectionné ;
- l’hypothèse de la transition de fonction : la mutation est initialement sélectionnée, car elle a une autre fonction bénéfique et la fonction de code inné se surajoute à ou supplante la précédente ;
- l’effet Baldwin : cette hypothèse est controversée, car elle utilise les principes dans la sélection naturelle, mais elle se base sur une vision lamarckienne. Le code inné est acquis culturellement, et le posséder de façon innée (via les gènes) devient un avantage, et épargne des coûts d’apprentissage.
Orrigi et Sperber notent que ces hypothèses ne peuvent s’appliquer au langage humain, car l’inné et l’acquis sont très importants.
La part de l’inné n’étant pas prépondérante, la probabilité que deux mutants aient des langages compatibles est infime. De plus, l’apprentissage du langage peut se faire avec des erreurs.
Le langage comme système de communication déductif
Ces chercheurs proposent donc de concevoir le langage humain comme un système de communication déductif, C'est-à-dire que le destinataire est capable de reconstruire l’information du communicant en tenant compte de ses intentions, bien qu’il n’ait pas exactement la même correspondance signe-sens. La faculté de langage est le fait d’être disposé à traiter un élément linguistique non codé et de le stabiliser comme un signal compréhensible. Cela nourrit la cohérence entre les langages. (À noter qu’ils se sont basés sur les travaux de Ruth Millikan[47]).
Avantages du langage
La plupart du temps, les individus ayant une forme de communication notable appartiennent à des espèces ayant un mode de vie social. Cette constatation a amené certains auteurs à se demander si la socialité n’était pas un prérequis à l’apparition du langage[48]. En effet, pour une bonne coordination, il est logique que la sélection naturelle ait fini par faire émerger des capacités de communications efficaces, expliquant le succès évolutif de ces espèces. On peut toutefois se demander sur quels mécanismes ce caractère a pu être sélectionné.
Stratégies de chasse et de fourragement
Un langage élaboré permet d’émettre des informations de façon d’autant plus précise que le moyen de communication est précis. De cette façon, des individus chassant ou fourrageant en meute ou en groupe peuvent facilement décupler leur efficacité en élaborant des comportements plus ou moins complexes. Un exemple assez connu est celui de la danse des abeilles : une éclaireuse ayant localisé une source de nourriture va effectuer devant ses congénères une danse qui leur permettra de savoir où trouver la nourriture, mais aussi en quelle quantité et de quel type[49]. De cette façon, les espèces sociales sont avantagées par rapport aux espèces non sociales en cas de compétition. Ce type d'explication reposant sur l'avantage collectif se heurte toutefois au problème de la tragédie des biens communs pour les individus non apparentés.
Résistance à la pression de prédation
Avoir un langage permet de développer plusieurs adaptations contre les prédateurs comme des mécanismes de guet ou d'alerte. On peut par exemple citer le chien de prairie, qui en un seul cri et selon la durée et l’intensité de celui-ci, peut avertir ses congénères du nombre de prédateurs en vue, de sa distance, de son type (volant ou terrestre) et de la vitesse à laquelle il approche[50] - [51]. De la même façon, une fourmi peut alerter sa colonie à l’aide de phéromones adéquates.
Meilleure adaptation aux contraintes environnementales
Certaines fourmis sont capables de s’agréger et de former des ponts vivants au-dessus de l’eau, voire des ilots flottant[52]. Ce comportement est une réponse à une contrainte environnementale, qui ne serait pas possible sans un minimum de « concertation » et donc de langage. On peut donc dire que face à un environnement induisant un stress, les espèces possédant un langage sont susceptibles de mieux réagir que les autres.
Ces paramètres constituent une liste non exhaustive des bénéfices qu’apportent le langage et la socialité. Ceci permet de dire que dans beaucoup de cas, les espèces ayant un langage et donc sociales auront tendance à avoir une meilleure compétitivité que les non sociales pour une même niche écologique. Cela a par exemple été montré sur une espèce d’abeille charpentière, Ceratina australensis[53]. L'argument de l'avantage écologique collectif est toutefois problématique dans une espèce où les individus interagissant ne sont pas apparentés.
Langage et signal
L'existence du langage humain pourrait trouver une explication dans la théorie du signal honnête, issue des travaux de l'éthologue Amotz Zahavi[54] - [55]. Le fait de fournir une information pertinente à ses congénères démontre qu'on la détient. Ce comportement d'affichage par le langage a pu évoluer dans un contexte où la capacité à s'informer constitue une qualité sociale recherchée. La théorie du signal honnête explique pourquoi les humains trouvent un intérêt à offrir des informations (et ne se contentent pas de prendre les informations des autres).
Langage humain, valeur sélective et culture
Ces observations sont parfaitement déclinables à l’Homme, qui possède la particularité unique du langage articulé. Il a en effet développé des stratégies de chasse en groupe et de défense contre les prédateurs, qui n’étaient possibles que grâce à ce moyen de communication, sur lequel il a bâti par la suite toute une société organisée, jusqu’à celle que l'on connaît aujourd’hui.
Le langage affecte la valeur sélective via l’expression et la communication de pensées. Ainsi, le langage contribue aux performances cognitives de l’individu.
De plus, le langage est mis en jeu dans la manipulation, la séduction, le maintien de relations sociales, etc.
Enfin, chaque individu bénéficie des perceptions et raisonnements des autres. L’individu dispose d’un très large éventail de savoirs et de connaissances qui traversent le temps et qu’il ne pourrait acquérir seul.
« Le caractère du langage est de procurer un substitut de l'expérience apte à être transmis sans fin dans le temps et l'espace, ce qui est le propre de notre symbolisme et le fondement de la tradition linguistique[56] ».
Notes et références
- Cet article est partiellement ou en totalité issu de l'article intitulé « Évolution du langage » (voir la liste des auteurs).
- Cet article est partiellement ou en totalité issu de l'article intitulé « Origines neurobiologiques du langage » (voir la liste des auteurs).
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Neurobiological origins of language » (voir la liste des auteurs).
- L'origine des langues.
- G. Marcus et Rabagliati, H., « What developmental disorders can tell us about the nature and origins of language », Nature Neuroscience, vol. 9, no 10, , p. 1226–1229 (DOI 10.1038/nn1766, lire en ligne)
- (en) Philip Lieberman & Edmund S. Crelin, « On the Speech of Neanderthal Man », Linguistic Inquiry, vol. 2, no 2, , p. 203-222.
- Lier, comme le fait Lieberman, l'émergence du langage à la descente du larynx n'est qu'une interprétation a posteriori qui n'explique en rien les origines du langage. « Dans un certain nombre d'espèces, la pression de la sélection sexuelle a conduit à deux phénomènes : d'une part, le fait que le larynx, placé relativement haut en situation de repos, descend lors de la production du signal ; d'autre part, le fait que le larynx dans certaines espèces s'est placé définitivement en position basse ». Chez l'homme, le larynx subit « une première descente vers la fin de la première année, en subit une seconde à la puberté, très minime chez les filles et, comme on le sait, importante chez les garçons, dont la voix change radicalement et descend vers les fréquences graves ». Cf Anne Reboul, Langage et cognition humaine, Presses universitaires de Grenoble, , p. 262.
- (en) Francesco d'Errico, Christopher Henshilwood, Graeme Lawson, Marian Vanhaeren, Anne-Marie Tillier; Marie Soressi; Frédérique Bresson; Bruno Maureille, April Nowell, Joseba Lakarra, Lucinda Backwell, Michèle Julien, « Archaeological Evidence for the Emergence of Language, Symbolism, and Music–An Alternative Multidisciplinary Perspective », Journal of World Prehistory, vol. 17, no 1, , p. 1–70.
- (en) Charles Perreault et Sarah Mathew, « Dating the Origin of Language Using Phonemic Diversity », PloS one, vol. 7, no 4, (DOI 10.1371/journal.pone.0035289)
- « En quoi le langage est-il spécifiquement humain ? », sur www.ac-grenoble.fr
- (en) Fisch, « Communication and Language in Animals. », Springer protocols Organism Models of Autism Spectrum Disorders, no Volume 100 of the series Neuromethods, , p. 265-282
- « Préférence manuelle et langage : existe-t-il vraiment un hémisphère dominant ? », sur www2.cnrs.fr,
- M. Arbib et Bota, M., « Language evolution: neural homologies and neuroimformatics », Neural Networks, vol. 16,
- J. P. Taglialatela, Russell, J. L., Schaeffer, J. A. et Hopkins, W. D., « Chimpanzee Vocal Signaling Points to a Multimodal Origin of Human Language », PLoS ONE, vol. 6, (DOI 10.1371/journal.pone.0018852)
- D Falk, « The Evolution of Broca's Area »
- L. Fadiga, Craighero, L. et D'Ausilio, A., « Broca's Area in Language, Action, and Music », The Neurosciences and Music III—Disorders and Plasticity, vol. 1169, , p. 448–458 (DOI 10.1111/j.1749-6632.2009.04582.x)
- David Boulton, « Terrence Deacon, Ph.D. - The Co-evolution of Language and the Brain » (consulté le )
- T.W. Deacon, « A role for relaxed selection in the evolution of language capacity », PNAS Early Edition, (lire en ligne)
- M. I. Sereno, « Language origins without the semantic urge », Cognitive Science Online, vol. 3, no 1, , p. 1–12 (lire en ligne)
- Jean-François Dortier, L'Homme, cet étrange animal. Aux origines du langage, de la culture et de la pensée, Éditions Sciences Humaines, , p. 147.
- Merritt Ruhlen, L'Origine des langues, Débats Belin, 1997 (ISBN 2-7011-1757-7).
- Russell D. Gray et Quentin D. Atkinson, « Language-tree divergence times support the Anatolian theory of Indo-European origin », Nature, no 426, , p. 435-439 (lire en ligne)
- Nicholas Wade, « Phonetic Clues Hint Language Is Africa-Born », The New York Times, (lire en ligne)
- Global Post, «World's languages traced back to single African mother tongue: scientists», 2011-04-15
- Dean Falk, « Prelinguistic evolution in early hominins: Whence motherese? », Behavioral and Brain Sciences, vol. 27, no 4, , p. 491–503 (ISSN 1469-1825 et 0140-525X, DOI 10.1017/S0140525X04000111, lire en ligne, consulté le )
- (en) E Hoff, Language Development, Belmont, CA, Wadsworth, , 528 p. (ISBN 978-0-495-50171-8, lire en ligne)
- (en) Stephen Anderson, Doctor Dolittle's Delusion : animals and the uniqueness of human language, Londres, Yale University Press, , 25–26 p. (ISBN 0-300-10339-5)
- (en) Lieberman, « Vocal tract limitations on the vowel repertoires of rhesus monkey and other nonhuman primates », Science, no 164, , p. 1185-1187
- (en) Fitch, « The descended larynx is not uniquely human », Proceedings of the Royal Society, no 268, , p. 1669-1675
- Voir le chapitre « Aux sources du langage » in Picq 2008.
- Jean-Jacques Hublin, « La langue des premiers hommes, in Hombert 2005.
- Philippe Vernier, « Évolution du cerveau et émergence du langage », in Hombert 2005.
- Voir à ce sujet la théorie de Changeux 1983 et le chapitre « Propriété des ensembles neuronaux » in Changeux 1979.
- (en) Matthew C. Tate, Guillaume Herbet, Sylvie Moritz-Gasser, Joseph E. Tate et Hugues Duffau, « Probabilistic map of critical functional regions of the human cerebral cortex: Broca’s area revisited », Brain, Oxford University Press, vol. 137, no 10, , p. 2773-2782 (DOI 10.1093/brain/awu168)
- Hugues Duffau et Estelle Saget (interviewer), « Hugues Duffau : Le cerveau se répare lui-même », L'Express, (lire en ligne, consulté le )
- Ruhlen 1994
- Christophe Coupé, « L’impossible Quête de la Langue Mère », in Hombert 2005.
- Pinker 1994, p. 353.
- Joseph Donato, « La Variation linguistique », in Linguistique sous la direction de Frédéric François, PUF, 1980.
- Bernard Victorri, « Les Mystères de l’émergence du langage », in Hombert 2005.
- (en) FOXP2 and the evolution of language.
- (en) Enard W, Przeworski M, Fisher SE, Lai CS, Wiebe V, Kitano T, Monaco AP et Pääbo S, « Molecular evolution of FOXP2, a gene involved in speech and language », Nature, vol. 418, no 6900, , p. 869–872 (DOI 10.1038/nature01025).
- (en) Hauser, « The faculty of language: what is it, who has it, how did it evolve? », Science, no 298, , p. 1569–1579
- (en) Jarvis, « Learned Birdsong and the Neurobiology of Human Language », Annals of the New York Academy of Sciences, no 1016, , p. 749-777
- (en) Traxler, « What's special about human language? The contents of the narrow language faculty revisited », Lang Linguist Compass,
- (en) Ray Jackendoff, « The nature of the language faculty and its implications for evolution of language (Reply to Fitch, Hauser, and Chomsky) », Cognition: International journal of cognitive science, no 97, , p. 211-225
- (en) Kirby, « The evolution of language », Oxford Handbook of Evolutionary Psychology,
- (en) Gloria Orrigi, Evolution, communication and the proper function of language, Carruthers, P. and Chamberlain, A., editors. Evolution and the Human Mind: Modularity, Language and Meta-Cognition, p. 140-169
- (en) Elliott Sober, The Nature of Selection : Evolutionary Theory in Philosophical Focus
- (en) Ruth Millikan, Language, Tought and Other Biological Categories
- (en) Steels, « Is sociality a crucial prerequisite for the emergence of language ? », Oxford University Press, , p. 36-57
- (en) Riley et al., « The flight paths of honeybees recruited by the waggle dance », Nature, no 435, , p. 205-207
- (en) « New Language Discovered: Prairiedogese », sur www.npr.org,
- (en) « Prairie dogs may have the most complex language », sur phys.org,
- Bert Holldobler et Edward O. Wilson (trad. de l'anglais), Voyage chez les fourmis, Galiax/Paris, Seuil, , 247 p. (ISBN 2-02-025628-2)
- (en) Rehan et al., « Fitness consequences of ecological constraints and implications for the evolution of sociality in an incipiently social bee. », Biological Journal of the Linnean Society, no 103, , p. 57-67
- (en) Amotz Zahavi et Avishag Zahavi (trad. de l'hébreu), The handicap principle - A missing piece in Darwin's puzzle, New York, Oxford University Press, , 286 p. (ISBN 0-19-510035-2)
- (en) Alan Grafen, « Biological signals as handicaps », Journal of Theoretical Biology, , p. 144, 517-546 (lire en ligne)
- Émile Benveniste, Problèmes de linguistique générale, Gallimard
Voir aussi
Articles connexes
- Essai sur l'origine des langues (Jean-Jacques Rousseau)
- Synopsis Universae Philologiae
- Langue originelle : recherches ethnologiques
- Langue adamique : utopie d'une langue biblique retrouvée
- Origine de la vie
- Proto-langue
Liens externes
- Notice dans un dictionnaire ou une encyclopédie généraliste :
Bibliographie
- Jean-Pierre Changeux, Théories du langage et théories de l'apprentissage, Paris, Seuil, .
- Jean-Pierre Changeux, L'homme neuronal, Paris, Fayard, .
- Jean-Louis Dessalles, Aux origines du langage : Une histoire naturelle de la parole, Paris, Hermès, .
- Jean-Marie Hombert (dir.), Aux origines des langues et du langage, Paris, Fayard, .
- Pascal Picq, Laurent Sagart, Ghislaine Dehaene et Cécile Lestienne, La plus belle histoire du langage, Paris, Hermès, .
- (en) Steven Pinker, The Language Instinct : The New Science of Language and Mind, Londres, The Penguin Press, , 494 p.
- (en) Merritt Ruhlen, The Origin of language : Tracing the evolution of the mother tongue, New York, John Wiley & Sons, , 239 p. (ISBN 0-471-58426-6).