Hérédité
L’hérédité est la transmission, au sein d'une espèce vivante ou d'une lignée de cellules, de caractéristiques d'une génération à la suivante. Les mécanismes de l'hérédité sont au cœur de la théorie de l'évolution car ils permettent l'accumulation des variations au fil des générations qui conduit à l'apparition de nouvelles espèces. En général, on associe l'hérédité aux gènes tels qu'ils furent découverts par Gregor Mendel mais d'autres mécanismes dits non mendéliens et épigénétiques peuvent aussi intervenir dans la transmission des caractères biologiques.
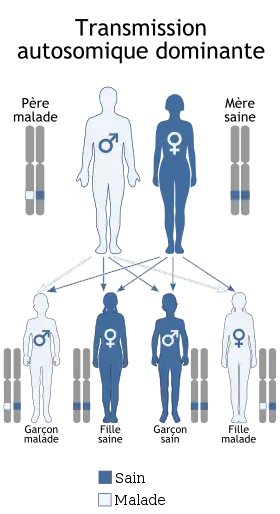
S'agissant d'espèces animales, y compris l'être humain, l'hérédité peut concerner des traits physiques mais aussi comportementaux bien que ces derniers puissent aussi être acquis par apprentissage ou imitation. La transmission des caractères acquis fit l'objet de nombreuses controverses dans l'histoire de la pensée évolutionniste, en particulier dans son application à l'homme et dans les tentatives eugénistes.
Aperçu général

Un exemple : l'hérédité de la couleur des yeux
Pour expliquer le concept d'hérédité, on peut prendre comme exemple la couleur des yeux dans l'espèce humaine ; il s'agit d'un trait biologique héréditaire. La couleur des yeux d'un individu dépend en effet d'un ensemble de gènes qui contrôlent la production de protéines dans l'iris qui lui donnent sa couleur. Il existe plusieurs variantes ou allèles de ces gènes, chacune donnant lieu à une couleur particulière des yeux, pour simplifier disons des allèles « yeux bruns », des allèles « yeux bleus », etc. L'ensemble des différentes variantes que possède un individu pour chacun des gènes de son génome est ce qu'on appelle son génotype. Le génotype de chaque individu est le résultat de la combinaison aléatoire au moment de la fécondation d'une partie du génotype de chacun de ses parents. Ainsi, si les deux parents possèdent des allèles « yeux bruns », les enfants auront dans leur génotype uniquement des allèles « yeux bruns ». Mais si l'un des parents possède des « yeux bleus » et l'autre des allèles « yeux bruns », les enfants hériteront chacun d'une partie des allèles « yeux bleus » de l'un et d'une partie des allèles « yeux bruns » de l'autre, dans des combinaisons à chaque fois différentes donnant lieu à des couleurs d'iris différentes pour chacun d'entre eux. Dans le cas de la couleur des yeux, c'est donc la transmission génétique qui gouverne l'hérédité de ce trait biologique.
Le phénotype combine hérédité et facteurs environnementaux
On appelle phénotype, l'ensemble des traits que l'on peut observer chez un individu : le fait d'avoir les yeux bruns est un trait phénotypique qui résulte du fait que cet individu possède l'allèle "yeux bruns" dans son génotype. Néanmoins, beaucoup de caractéristiques phénotypiques ne sont pas directement le reflet d'un génotype particulier. Le plus souvent les traits sont le résultat d'une combinaison de facteurs héréditaires transmis par les gènes et de facteurs environnementaux intervenant au cours de la vie de l'individu. Par exemple, la couleur de la peau dépend à la fois du génotype et de l'ensoleillement, si bien qu'on ne peut pas dire que le bronzage est héréditaire. Par contre la capacité à bronzer, c'est-à-dire à produire de la mélanine, est héréditaire puisqu'elle est gouvernée par des gènes qui sont transmis d'une génération à l'autre.
Les bases biologiques de l'hérédité

Le principal support de l'hérédité est donc les gènes. Le mécanisme par lequel l'information génétique est transmise repose sur la molécule d'acide désoxyribonucléique plus connue sous l'acronyme ADN. Cette longue molécule est un polymère composé de bases nucléiques. La séquence de ces bases nucléiques dans la molécule d'ADN code l'information génétique à la manière dont la séquence des lettres dans une phrase forme des mots qui lui donnent son sens. Ce sont des portions particulières de la molécule d'ADN qui forment les gènes, les différents allèles correspondant à des séquences proches mais pas strictement identiques du gène. Lorsqu'une mutation se produit qui transforme la séquence d'ADN, on obtient donc un nouvel allèle qui se traduira, sur le plan phénotypique, par une modification du trait biologique contrôlé par le gène muté. Si cette mutation est transmise aux descendants alors le nouvel allèle peut, au fil, des générations se répandre dans la population.
Les mécanismes connus pour l'hérédité biologique sont de plusieurs types :
- la transmission génétique classique, dite aussi hérédité mendélienne, est le résultat de la réplication de l'ADN lors de la division cellulaire ;
- la transmission épigénétique procède par des modifications qui n'altèrent pas la séquence de l'ADN mais sa structure ;
- l'hérédité non mendélienne désigne les autres formes d'héritage biologique, notamment celles fondées sur les constituants du cytoplasme, comme les mitochondries ou les chloroplastes.
Parmi les autres formes de transmission biologique, on peut citer la transmission lamarckienne qui désigne la transmission des caractères acquis. Celle-ci ne passe pas par une modification génétique ou épigénétique mais par des processus autres tels l'apprentissage, l'imitation…
Histoire du concept
Une signification qui a évolué
Bien avant d'être reprise dans un cadre biologique, la transmission héréditaire a décrit un fait juridique : un fief, un titre, un office pouvaient être héréditaires, c'est-à-dire transmis par héritage. Il faut donc être très prudent dans l'usage de ce terme. Il est juste de dire que le métier est héréditaire dans une société où l'on est forgeron de père en fils. Cela ne signifie bien sûr pas que les descendants soient génétiquement des forgerons.
On peut faire la distinction entre l'hérédité sociale (ou acquise) et l'hérédité génétique (ou innée).
Dans l'hérédité acquise, on trouvera tous les caractères hors génétique qui viennent de l'imitation ou de l'opposition aux parents et au milieu social fréquenté : la langue maternelle, la culture, les valeurs, le métier, la fortune, les idées politiques…
L'hérédité innée concerne uniquement les caractères génétiques transmis par les parents. L'utilisation du terme hérédité en biologie ne s'est mise en place qu'au cours du XIXe siècle. Ainsi, Louis Pasteur parle d'hérédité pour la transmission de particules infectieuses entre deux générations de papillon[1]. Il parle d'« hérédité congénitale » pour sous-entendre le caractère génétique (le terme génétique n'existait pas à cette époque).
Cette confusion qui perdure jusqu'à la fin du XIXe siècle montre qu'il y eut longtemps confusion entre différents types d'hérédité.
La compréhension du mécanisme de l'hérédité
La discipline qui étudie l'hérédité biologique est la génétique. C'est une discipline récente, qui s'est constituée au cours des XIXe et XXe siècles, au fur et à mesure des découvertes conceptuelles et expérimentales.
Au XIXe siècle, Gregor Mendel (1822-1884) formule et démontre des règles de transmission des caractères, les lois de Mendel, qui sont à la base de la génétique formelle. Parallèlement, les premières expériences de biologie cellulaire montrent l'importance des chromosomes dans cette transmission des caractères.
Il fallut attendre en 1943 la démonstration par Oswald Avery que l'ADN était bien le support de l'information génétique, et la découverte de sa structure en double hélice par Watson et Crick en 1953, pour commencer à comprendre les fondements moléculaires des lois de Mendel.
Les différents modes de transmission biologique des caractères
L'hérédité mendélienne
L'hérédité mendélienne régit la plupart des caractères.
Chez les organismes diploïdes comme les humains, chaque individu possède deux copies de chaque gène, une copie venant de la mère et l'autre du père (exception : les gènes des chromosomes sexuels). Si ces deux versions sont identiques, l'individu est dit homozygote pour le gène considéré. Dans le cas contraire, il possède deux versions différentes (deux allèles différents), et il est dit hétérozygote pour ce gène.
Par exemple, un enfant possédant deux fois l'allèle « A » pour le gène du groupe sanguin sera de groupe A, alors qu'un enfant hétérozygote avec un allèle « A » et un allèle « B » sera de groupe AB. Ces deux allèles s'expriment également : on parle de codominance.
Mais tous les allèles ne sont pas codominants : l'allèle O (zéro, c'est-à-dire pas de marqueur) ne l'est pas. Un homozygote « O/O » est de groupe O, mais un hétérozygote « A/O » est de groupe A, « B/O » de groupe B. On dit que l'allèle O est récessif, et les allèles A et B dominants par rapport à O.
Hérédité non mendélienne
Il existe des caractères qui ne suivent pas les lois de la génétique mendélienne, on parle alors d'hérédité non mendélienne.
Il peut s'agir d'une forme d'hérédité extranucléaire, qui correspond à la transmission des caractères liés à l'ADN présent non dans le noyau mais dans des organites à doubles membranes comme les mitochondries (organites de la respiration cellulaire) et les chloroplastes (organites de la photosynthèse chlorophyllienne). En général ces organites sont apportés par l'ovule. Par exemple, le spermatozoïde humain laisse ses mitochondries hors de l'ovule lors de la fécondation. Les caractères sont donc issus des mitochondries maternelles : on parle de transmission mitochondriale matrilinéaire. On retrouve ce mode de transmission héréditaire dans les maladies mitochondriales.
Il existe également une autre voie d'hérédité qualifiée d'hérédité cytoplasmique où ce sont des composants comme les protéines qui sont transmis. C'est le cas pour la transmission de l'ESB et d'autres maladies à prions.

Hérédité épigénétique[2]
Autrefois cru impossible, il semblerait que dans certains cas il est possible pour une progéniture d’hériter de l’état d’un caractère épigénétique développé selon les conditions environnementales auxquelles le parent fut soumis. L’héritabilité épigénétique, ou plasticité phénotypique intergénérationnelle, réfère donc à la transmission transgénérationnelle de traits dont l’expression n’est pas déterminée de façon proximale par l’ADN. Ainsi, ce phénomène est plutôt lié à la transmission entre les générations des modifications sur la chromatine et sur la régulation des gènes qui sont sensibles aux stimuli environnementaux. L’héritabilité épigénétique est étudiée dans deux optiques principales, soit l’effet maternel où la transmission de molécules maternelles aurait comme fonction de maintenir ou établir des modifications à la chromatine chez la progéniture, et l’héritabilité directe de matériel génétique dont l’état d’expression est altéré par des états modifiés de la chromatine. Enfin, de plus en plus d’études récentes démontrent la possibilité d’hériter de caractères acquis par les parents, ou « hérédité douce » (traduction de l’anglais soft inheritance), phénomène qui est fondamentalement différent des concepts d’hérédité classique basée sur l’ADN parental.
Il est d’ailleurs possible d’étudier ce phénomène afin d’élucider son mécanisme d’action, par exemple au niveau des défenses visant les herbivores présentes chez plusieurs plantes. En identifiant les gènes responsables de l’expression de ces défenses, il devient possible de comparer leur expression entre les individus et tracer un lien entre l’abondance relative des défenses entre les générations en fonction des conditions environnementales parentales. En effet, en comparant les progénitures de plantes de Mimulus guttatus ayant subi des dommages simulés aux feuilles à celles de plantes n’ayant subi aucun dommage (contrôle), il fut observé que les progénitures dont les parents avaient subi des dommages exprimaient davantage de trichomes foliaires que les progénitures du groupe contrôle, et ce même si elles ne subissaient elles-mêmes aucun dommage. Ainsi, l’augmentation dans la densité de trichomes foliaires est une réponse épigénétique héritable aux dommages liés à l’herbivorie qui est exprimée de façon différentielle chez la progéniture selon l’état d’expression par le parent chez Mimus gutatus. Cette hérédité épigénétique serait liée à la variation dans l’héritabilité épigénétique de l’induction de ce trait. Dans le cadre de la biologie évolutive du développement, ce changement dans l’expression de la densité des trichomes correspondrait à une hétérométrie.
Le mécanisme sous-jacent à ce phénomène fut élucidé en mesurant et en comparant chez les progénitures de 4 lignées recombinantes homozygotes l’expression relative de gènes potentiellement responsables du développement des trichomes chez cette espèce. Ces gènes candidats furent identifés en comparaison avec des gènes connus responsables de la formation des trichomes chez des espèces de plantes modèles telles que Arabidopsis. En évaluant les interactions entre la variation génétique et épigénétique au niveau phénotypique et en comparant les associations entre ces variations et l’expression de gènes candidats, une association fut tracée entre l’expression du gène MYB MIXTA-like 8, la densité en trichomes et la réponse au dommage parental. Ce gène serait alors un régulateur négatif potentiel du gène GL2, responsable du développement des trichomes chez Mimulus guttatus. Les dommages foliaires chez le parent induiraient une régulation négative de l’expression de MYB MIXTA-like 8 chez la progéniture, ce qui permettrait l’expression accrue de GL2, ce qui est associé à l’hérédité épigénétique de la densité foliaire accrue des trichomes. Ainsi, il est probable que l’héritabilité épigénétique soit liée à la transmission d’un état d’expression d’un gène en réponse aux conditions parentales plutôt qu’à la transmission du gène lui-même.
Ces découvertes, et le concept d’héritabilité épigénétique même, impliquent que la variabilité héritable n’est pas nécessairement le résultat de différences nucléotidiques et que le hasard n’est pas la seule source de variation du matériel héritable, mais que l’environnement peut exercer un effet directionnel vers des phénotypes spécifiques.
Notes et références
- Pasteur, Œuvre tome 4 – Étude sur la maladie des vers à soie, p. 63
- (en) Alison G. Scoville, Laryssa L. Barnett, Sarah Bodbyl-Roels et John K. Kelly, « Differential regulation of a MYB transcription factor is correlated with transgenerational epigenetic inheritance of trichome density in Mimulus guttatus », New Phytologist, vol. 191, no 1, , p. 251–263 (ISSN 0028-646X, DOI 10.1111/j.1469-8137.2011.03656.x, lire en ligne, consulté le )
Voir aussi
Bibliographie
- J. J. Matras et Georges Chapouthier, L'inné et l'acquis des structures biologiques, coll. "Le Biologiste", Presses Universitaires de France éditeur, Paris, 1981
Articles connexes
Liens externes
- Ressources relatives à la santé :
- (en) Medical Subject Headings
- (no + nn + nb) Store medisinske leksikon
- Notices dans des dictionnaires ou encyclopédies généralistes :