Socialité
La socialité désigne la tendance d'une population animale à vivre en groupe. Il en existe plusieurs niveaux, allant jusqu'à la formation de sociétés animales.
Comme dans tout procédé évolutif, il existe une certaine variabilité de caractères entre les individus d'une population. Lorsque certains d'entre eux affectent de manière positive la fitness, c'est-à-dire la valeur reproductive, de ceux-ci, ces caractères prendront une part relative plus importante au sein de la population et seront transmis aux descendants, selon la sélection naturelle. La socialité est également soumise à cette sélection, et les théories et modèles qui expliquent son origine et son maintien sont diverses.
Pour parler de socialité il est nécessaire que certains critères soient respectés : d'une part des comportements de coopération et, d'autre part, une attraction réciproque entre les individus, c'est-à-dire un regroupement d'animaux du à des facteurs venant de leurs congénères et non de l'environnement.
La socialité est présente sous différentes formes qui se distinguent par leur degré d'évolution :
- stade grégaire : caractérisé par une forme très primitive de la socialité, il se limite à de l’attraction réciproque ou interattraction. Ce stade de la socialité correspond à des groupements d’animaux résultant de facteurs (signaux chimiques, contacts) émanant de leurs congénères et non de l’environnement. Et se traduit par une synchronisation des activités du groupe (nutrition, sommeil, déplacement, fuite). C’est la première étape vers la socialisation.
- stade subsocial : mise en place de comportements parentaux (manipulation des œufs, défense de la progéniture, voire son nourrissage).
- stade colonial : les soins aux jeunes ont lieu dans un site d'élevage commun à plusieurs femelles, mais où ces dernières ne coopèrent pas.
- stade communal : les femelles coopèrent dans les soins aux jeunes mais il n'y a pas de spécialisation des tâches.
- stade eusocial : mode de vie sociale le plus élaboré.
Edouard O. Wilson (1975) définit trois critères nécessaires pour pouvoir parler d'eusocialité :
- la coopération dans les soins aux jeunes
- le chevauchement d'au moins deux générations d'individus, durant une partie de leur vie, les descendants peuvent assister leurs parents.
- l'existence d'individus spécialisés dans la reproduction. Cette division du travail (ou polyéthisme) conduit à une société divisée en castes ; on distingue des individus féconds et des individus stériles.
Historique

Selon Charles Darwin (1809-1882), célèbre naturaliste anglais, les mots-clés de la socialité sont « Instincts sociaux, affections parentales et sociales, sympathie, lutte et principe d’utilité de groupe. »
Dans son ouvrage, La descendance de l’Homme, il s’attarde notamment sur la socialité d’un point de vue anthropomorphique (Darwinisme social) : « L'impression de plaisir que procure la société est probablement une extension des affections de parenté ou des affections filiales ; on peut attribuer cette extension principalement à la sélection naturelle, et peut être aussi, en partie, à l'habitude. Car, chez les animaux pour lesquels la vie sociale est avantageuse, les individus qui trouvent le plus de plaisir à être réunis peuvent le mieux échapper à divers dangers (…). Il est inutile de spéculer sur l’origine de l’affection des parents pour leurs enfants et de ceux-ci pour leurs parents ; ces affections constituent évidemment la base des affections sociales. »
Aujourd'hui on compare les contributions de William Donald « Bill » Hamilton (1936-2000), biologiste britannique de l’évolution, sur la valeur sélective inclusive (ou fitness inclusive) à celles de Darwin pour la sélection naturelle ou encore à celles de Newton pour la mécanique classique. Cependant, il n’est pas complètement à l’origine de la sélection de parentèle (ou kin selection); en effet avant lui J.B.S. Haldane (1955), Ronald Fisher (1958) et George C. Williams (1957) ont travaillé sur la génétique des populations et forgé l’idée de sélection de parentèle mais aucun d’eux n’a su synthétiser autant de détails que dans la théorie émise par Hamilton. Il s’est également appuyé sur des biologistes du comportement, Vero Copner Wynne-Edwards (1962) et William Emerson Ritter (1960) qui ont émis des hypothèses à propos de la coopération dans la nature.
Plusieurs biologistes du comportement et généticiens des populations avaient en effet déjà émis des hypothèses et théories liées à l’évolution de la socialité, comme Vero Copner Wynne-Edwards à propos de la sélection de groupe ou John Maynard Smith sur la sélection de groupe et de parentèle.
En 1964, Hamilton publie Genetical Evolution of Social Behaviour dans lequel il établit ce qu’est pour lui la sélection de parentèle et la fitness inclusive. Il y écrit : « Le comportement social d'une espèce se développe de telle façon que dans chaque situation évoquant un comportement distinct, l'individu semble comparer la valeur sélective de ses voisins à la sienne selon les coefficients de relation appropriée[1] ». L’article d’Hamilton marque vraiment la mise en place de la sélection de parentèle telle que nous la connaissons encore aujourd’hui. Il a accompagné ce travail de divers articles notamment sur la sélection sexuelle, la théorie de la sénescence, le sex-ratio… Il se penche plus particulièrement sur l’eusocialité et tente d’expliquer son évolution.
Depuis longtemps, les scientifiques ont émis des hypothèses concernant l’eusocialité, ce phénomène ultime et paradoxal de l’évolution de la socialité. Darwin finira par dire dans son ouvrage The Origin of Species qu’une colonie d’insectes eusociaux constitue une unité de sélection. Aujourd’hui encore cette proposition est largement suivie par les chercheurs actuels, en effet l’eusocialité n’est pas un phénomène marginal dans le monde vivant et de nombreux insectes notamment des hyménoptères ont développé les caractéristiques de ce stade social. En 1964, W.D. Hamilton a formulé une théorie qui a permis d’interpréter l’évolution génétique de l’eusocialité, la sélection de parentèle. Il a également mis en place le concept de fitness inclusive. Elle mesure l’effet de la sélection qui agit sur un groupe d’individus au niveau des gènes.
Hamilton explique dans sa théorie que la coopération est favorisée par la sélection naturelle si la relation entre les deux individus est supérieure au ratio coût/bénéfice. Il reviendra plus tard sur cette formule en s’apercevant que la fitness inclusive semble avoir des limites. La sélection de parentèle a été acceptée comme un élément indispensable de la théorie sociobiologique. Cependant Hamilton a permis cela en émettant une autre hypothèse, celle de l’haplodiploïdie. Il montre que les colonies d’individus altruistes évoluent plus fréquemment vers une situation haplodiploïde que diplodiploïde permettant ensuite une sélection de parentèle et peu à peu la mise en place de socialité et plus particulièrement d’eusocialité.
Dans les années 1960 et 70, toutes les espèces eusociales appartenaient aux hyménoptères, cela confortait donc Hamilton dans sa théorie et renforçait le rôle clé de la sélection de parentèle. Mais les années 1990 viennent mettre un point noir dans cette théorie, de nouvelles espèces eusociales ont montré qu’elles ne suivaient pas ce modèle. L’hypothèse haplodiploïde fut en partie abandonnée. Cette perception de l’évolution de l’eusocialité est ici une vue de l’article de Nowak[2] qui a donné lieu à un débat relativement virulent entre son auteur et d’autres chercheurs qui n’étaient pas d’accord avec sa démarche.
Au travers de la biologie moléculaire et de la génétique, les chercheurs tentent aujourd’hui de comprendre l’effet des gènes dans l’évolution de la socialité. Ils ont su définir une composante moléculaire appelée « taux d’évolution moléculaire » liée aux taux de mutations dans le génome. D’après les études actuelles, ce sont les comportements sociaux de base, les interactions sociales qui ont permis :
- Une réduction « efficace » de la taille des populations, on parle ici en termes de population de gènes, l’apparentement génétique est d’autant plus fort que le degré de socialité est élevé.
- Il semblerait que les gènes mitochondriaux aient plus d’effet sur la socialité que les gènes nucléaires.
- Le degré de socialité (stade) est corrélé à ce taux. Il correspond au taux d’apparition de nouvelles protéines dans le génome d’un individu.
Pour résumer ces quelques points : il existerait des gènes mitochondriaux qui favoriseraient certains comportements sociaux, ces derniers conduiraient à des « plus petites populations génétiques » qui seraient affectées de manière plus importante par les mutations du génome. Ce phénomène aurait donc tendance à fixer la socialité comme une caractéristique génétique et donc héritable.
Les grandes forces motrices de l'évolution de la socialité
La coopération mutuelle (mutualisme)
Pour comprendre l’évolution de la socialité, il faut se demander pourquoi les individus s’associent. La coopération a une action positive sur la fitness, c’est-à-dire la capacité à survivre et à se reproduire des individus, résultant d’un bénéfice net pour chacun. Clutton-Brock publie en 2002 son étude sur la reproduction chez les Vertébrés. Dans certaines sociétés aviaires, par exemple, la femelle qui élève sa progéniture est un partenaire potentiel pour les mâles, car ces derniers en l'aidant à se nourrir, par exemple, elle et sa progéniture augmentent leurs chances de se reproduire avec cette même femelle dans le futur[3].
Pour expliquer l’apparition de la coopération entre individus non apparentés, John Maynard Smith et George R. Price (1973) ont appliqué la théorie des jeux en écologie comportementale et évolutive. Celle-ci a pour but de résoudre les conflits d’intérêts entre les joueurs en trouvant la solution la moins mauvaise pour chacun. Dans un jeu à deux joueurs, dans quelles conditions doivent-ils coopérer et dans quels cas ne le doivent-ils pas ?
| Joueur 1 / joueur 2 | Coopération | Défection |
|---|---|---|
| Coopération | (B ; B) | (C ; T) |
| Défection | (T ; C) | (P ; P) |
|
B : bénéfice de la coopération mutuelle | ||
- Dans le cas où B > T et C > P, la coopération mutuelle sera la seule stratégie évolutivement stable (ESS). Une stratégie évolutivement stable (cas particulier de l’équilibre de Nash) permet aux individus de ne pas être envahis (par un mutant par exemple).
- Dans le cas où B > T et P > C, cette fois-ci la coopération mutuelle ET la défection mutuelle sont deux ESS (synergisme).
- Dans le cas où T > B et C > P ni la coopération mutuelle ni la défection mutuelle ne sont des équilibres (Cruel Bind).
- Dans le cas où T > B et P > S la défection mutuelle est une solution stable mais sous optimale. (dilemme du prisonnier.)
Brown JS et Vincent TL ont publié 2008 des travaux selon lesquels la fitness des individus devrait augmenter en fonction du nombre d’individus dans le groupe. De ce fait, cela apporte inévitablement de la compétition dans le groupe. La taille de ce dernier doit être suffisamment grande pour surpasser les variations de l’environnement et ainsi permettre la colonisation d’environnements ou de niches écologiques que ne seraient pas capable d'occuper des populations composées entièrement d'individus solitaires[4].
Certaines contraintes comme une intense prédation, une forte compétition interspécifique ou intraspécifique, des environnements imprévisibles ou des ressources difficiles à acquérir, pourraient être à l'origine de l’apparition de la coopération. Il est donc possible que la socialité ait évolué chez des espèces durablement installées ayant des croissances satisfaisantes, mais qui en coopérant, ont pu avoir accès à d’autres ressources plus abondantes. D'autre part, la fitness d'un individu serait uniquement basée sur le profit associé à son comportement individuel et ne serait pas influencée par la fitness d’autres individus du même groupe[5].
L'évolution de l'altruisme par la sélection de parentèle
La sélection de parentèle est une théorie formulée par William Donald Hamilton en 1964 [6]. Elle apparaît comme un mécanisme majeur pour expliquer l’évolution de la socialité dans le règne animal, plus précisément l’apparition de comportements altruistes, c’est-à-dire des actes qui apportent des bénéfices pour l’individu qui les reçoit et un coût pour celui qui les procure. Celle-ci s'appuie sur le principe selon lequel la fitness d'un individu ne s'appuie pas seulement sur son succès reproducteur, mais également sur celui des individus qui lui sont apparentés ; le bénéfice est alors indirect pour cet individu. La somme des valeurs sélectives directe et indirecte est appelée fitness inclusive ; c'est-à-dire qu'elle combine à la fois les descendants produits par l'animal, mais également le fait que ses gènes soient transmis par les autres.
La sélection de parentèle s’exerce donc au niveau des gènes : les individus peuvent privilégier des comportements de coopération avec leurs apparentés afin de maximiser la propagation de leur propre génome. Cependant Hamilton énonce une limite à ce principe : en réalité la sélection va favoriser un acte altruiste uniquement si les bénéfices de l'acte pour le donneur (augmentation indirecte de la fréquence de ses gènes) dépassent les coûts. De ce fait il établit une relation mathématique qui prédit qu’un gène altruiste sera favorisé par la sélection naturelle si :
- r B > C
- r est le coefficient d’apparentement, il représente la proportion de gènes qui est partagée par les individus
- C le coût de l’acte pour l’individu altruiste
- B le bénéfice pour le receveur
Plus les individus sont proches, plus leur coefficient d'apparentement est grand, donc la probabilité d’observer un acte altruiste sera d’autant plus importante. Cependant il faut remarquer que même avec un faible coefficient d’apparentement, le produit obtenu avec le bénéfice peut surpasser le coût, notamment quand celui-ci est très faible. De plus, le bénéfice doit être pris sur l’ensemble des apparentés: même un coût élevé pour un individu peut être compensé s’il confère un bénéfice à un grand nombre d’apparentés. Ce modèle permet ainsi d’expliquer le paradoxe de l’abeille qui meurt lorsqu’elle pique : en piquant un potentiel intrus son acte altruiste lui coûte la vie mais celle-ci défend sa ruche et augmente de ce fait les chances de survie de ses congénères.
Le degré de parenté chez des individus diploïdes, où chaque parent transmet la moitié de ses gènes à sa descendance, est facilement déterminable en retraçant le nombre de connexions généalogiques. Le coefficient de parenté vaut donc par exemple 0,5 entre parents et enfants ; 0,5 entre frères et sœurs ; 0,25 entre les grands parents et les petits enfants ou encore 0,125 entre cousins. Ainsi chez de nombreux oiseaux, on constate que les jeunes aident leurs parents à élever une nouvelle génération plutôt que de partir se reproduire eux-mêmes.
Le modèle de la sélection de parentèle d’Hamilton explique également l’apparition de castes de type reine/ouvrières avec division du travail chez des espèces d’hyménoptères haplodiploïdes comme les abeilles, guêpes ou encore fourmis comme l'ont montré les biologistes Robert L.Trivers et Hope Hare[7].
La sélection de groupe
En 1962, Wynne-Edwards formule la théorie de la sélection de groupe selon laquelle on peut considérer le groupe comme unité de sélection, indépendamment de l'apparentement entre les individus. C'est alors les caractéristiques du groupes (et leurs modifications) qui régissent leur fitness. Bien qu'on ne trouve aujourd'hui plus vraiment d'exemple de sélection de groupe que la sélection de parentèle ne puisse pas expliquer, cette théorie reste fréquemment citée car elle reste la première à postuler le groupe comme étant une unité de sélection. En effet, elle est antérieure à la sélection de parentèle. Le rejet de la sélection de groupe par une partie de la communauté scientifique est basé sur la focalisation des gènes capable de se reproduire à l'identique (réplicateur) et ne répond pas à la problématique de l'organisation fonctionnelle des groupes par rapport aux individus. Pour d'autres, la sélection de groupe apparaît comme une force importante dans la nature et la sélection de parentèle devient un cas particulier de la sélection de groupe[8].
Chez les invertébrés
La socialité évolue à différents niveaux et indépendamment chez chaque classe d'invertébrés en réponse aux différentes pressions de sélections qu'ils rencontrent dans leurs milieux. Le cas le plus cité reste cependant l'évolution de l'eusocialité chez certains hyménoptères et isoptères.
Le grégarisme

La communication intraspécifique (facteurs chimiques…) joue un rôle majeur dans la mise en place de ce type de socialité: elle permet l’attraction mutuelle entre individus de la même espèce. Cette interattraction profite à l’ensemble de la population, que ce soit au niveau de la dispersion des individus, de l'acquisition des ressources ou de la résistance à la prédation. Ces caractéristiques grégaires sont maintenues car elles diminuent le coût énergétique alloué à la reproduction ou à leur survie.
Si l'on prend l’exemple du criquet pèlerin Schistocerca gregaria, celui-ci est inoffensif à sa naissance, dans sa forme solitaire, et peut être dévastateur sous sa forme grégaire. Certaines conditions environnementales affectent l'aire de répartition du criquet. Le changement de comportement de solitaire à grégaire est ensuite dû à un neuromédiateur, la sérotonine. Le bain de foule entraîne l’augmentation de la production de cette hormone, provoquant des changements majeurs chez l’individu solitaire: des changements comportementaux (nuées de criquets), mais aussi physiologiques (adaptation au long vol) et morphologiques (couleur de la cuticule).
En quelques heures, deux individus solitaires en contact peuvent passer à un stade social primitif. On trouve du grégarisme chez de nombreuses autres espèces d’insectes telles que les araignées, certaines espèces de fourmis, etc.
Certaines formes de grégarisme existaient déjà à l'Ordovicien : il y a environ 480 Ma, des trilobites mâles de l'espèce Ampyx priscus se déplaçaient en file indienne, espacées presque régulièrement[9].
Les stades subsociaux, coloniaux et communaux
Les stades subsociaux, coloniaux et communaux semblent avoir une plasticité importante chez les invertébrés : de nombreux cas de réversions brutales entre chacun de ces stades sont référencés dans la littérature scientifique. Ces différents stades sociaux sont similaires dans le comportement parental notamment via la formation d’un nid commun avec une complexité croissante dans l’aide aux jeunes.
Par exemple, le terme d’araignées sociales est employé pour différents comportements sociaux, il regroupe:
- des espèces au comportement parental, les jeunes vivent pendant un temps limité dans la structure élaborée par la mère (stade subsocial),
- des espèces qui forment des associations de toiles individuelles (sociales territoriales : forme de grégarisme). Il y a chez ces espèces une cohésion du groupe qui semble relativement lâche, avec des comportements coopératifs et solitaires dans ces associations de toiles,
- des espèces qui exploitent collectivement une toile commune. Ce dernier groupe renferme ce qu’on appelle habituellement les araignées sociales non territoriales. Celles-ci coopèrent dans la construction du piège, la capture des proies, les soins aux jeunes et la défense de la colonie (stade colonial et communal).
Ces comportements coopératifs et toiles communes permettent l'économie de soie, mais aussi la capture de proies plus importantes. Les bénéfices sont nets pour chaque protagoniste de ces interactions sociales. Chez certaines araignées comme Anelosimus eximius, certaines phéromones dont la fonction est établie comme essentielle dans les comportements reproductifs, semblent également être le facteur principal de la cohésion du groupe.
Le rôle de la soie dans la cohésion de groupe est souligné par le fait qu’il n’existe pas d’espèce sociale dépourvue de structures soyeuses. Certains auteurs ont souligné l’exception de Diaea socialis et Delena cancerides dont les individus vivent dans un nid soyeux mais chassent de manière solitaire à l’extérieur et sans piège. Ces deux exemples ne font que renforcer l’idée que la soie est essentielle car la vie des araignées sociales ne se déroule que dans le nid commun.
L’organisation sociale chez ces araignées communales, est liée à un processus stigmergique dans la mise en place de la toile : le comportement d’un individu est piloté par le produit du comportement d’un congénère. De plus, le passage d’un stade solitaire à sociale semble être directement lié à l’utilisation de la soie chez les araignées.
L’eusocialité
L’analyse de l’origine évolutive de l’eusocialité est plus aisée dans des taxons ayant des niveaux de socialité varié (solitaire et eusociale) ainsi que dans les taxons où l’eusocialité à une origine relativement récente.
Bien que les termites, les fourmis et les abeilles soient des organismes modèles pour la compréhension de l’organisation eusociale des colonies d’insectes et son maintien, ces groupes donnent peu d’informations quant à l’origine évolutive de l’eusocialité car ils sont très anciens et leurs comportements sociaux sont très bien fixés et donc peu variables.
La abeilles de la famille halictidae sont de bons modèles étudier l'eusocialité en raison de la diversité des comportements sociaux selon les espèces. Certaines montrent des pontes solitaires, des pontes communes (stade coloniaux, communaux) et de l’eusocialité primitive. Les variations de structure des colonies annuelles qui vont d’une reine et 5 travailleurs, à plus de 500 travailleurs sont également un bon outil pour mettre en évidence les différentes caractéristiques de l'eusocialité.
Les abeilles ont une vie communautaire dans un nid, leurs caractéristiques sociales sont les suivantes :
- Communication visuelle importante et complexe
- Polymorphisme lié au sexe et à la caste
- Polyéthisme d’âge (division du travail en fonction de l’âge) chez les ouvrières
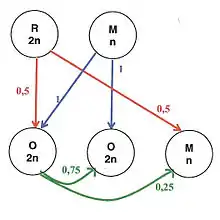
L’activité des ouvrières en fonction de leur âge, se déplace graduellement de l’intérieur de la ruche vers l’extérieur, depuis le nettoyage des alvéoles jusqu’au butinage qu’elles effectueront jusqu’à leur mort. La reine est diploïde et ne transmet qu'une partie de ses chromosomes aux ouvrières comme aux mâles. Le mâle lui est haploïde et ne transmet ses chromosomes qu’aux ouvrières. Les reines peuvent pondre des œufs non fécondés qui donneront naissance à des mâles (parthénogénèse).
Cette asymétrie de parenté peut générer des conflits. La femelle reproductrice est apparentée de façon équivalente à ses filles et ses fils. Elle peut optimiser son succès reproducteur en favorisant un sex-ratio équilibré. Tandis que les ouvrières qui sont beaucoup plus apparentées à leurs sœurs qu’à leurs frères, auront tendance optimiser leur succès reproducteur en biaisant le sex-ratio en faveur du sexe femelle. Il y aura donc un sex-ratio primaire, dans la proportion d’œufs haploïde et diploïde produit par la reine. Mais aussi un sex-ratio secondaire dans les proportions d’adultes de chaque sexe. L’existence même de ces conflits « d’intérêt génétique » confirme le rôle majeur de l’apparentement dans l’évolution de ces sociétés.
Une étude sur l’évolution de la socialité chez les abeilles halictides montre que les caractéristiques importantes de la complexité sociale[10] (ex : taille des colonies, dimorphisme reine/ouvriers…) ne correspondent pas à une phylogénie spécifique (bien qu’elle permet de diminuer les origines de l’eusocialité ainsi que d’identifier les reversions dans le comportement social) mais plutôt à des facteurs environnementaux. En effet des inversions vers des pontes solitaires sont mises en relation à des changements de plantes hôtes. Aussi, des espèces largement répandues montrent des variations intraspécifiques dans la socialité associée à des différences d’altitude et de latitude. D’après ces facteurs environnementaux, il semble que l’eusocialité est très instable chez les abeilles halictide. De plus leur diversité sociale est attribuable à des origines eusociales nombreuses et indépendantes dont une douzaine de réversions ont déjà été référencées.
Les soins aux jeunes par les ouvrières est souvent décrit comme un grand paradoxe évolutif. Des milliers d'ouvrières stériles vont allouer toute leur énergie aux soins des descendants de leur mère. De nombreuses théories ont été formulées quant au maintien de ce comportement pour le moins inhabituel dans le monde du vivant où la sélection de parentèle semble avoir été poussée à son paroxysme (l'haplo-dipoïdie était d'abord avancée pour justifier cet état), et certaines d'entre elles sont aujourd'hui toujours débattues (théorie du despotisme parental).
De nombreux mécanismes et pressions (sélection de parentèle, conditions environnementales) ont permis l'évolution d'un stade social particulièrement complexe et très étudié ou différents protagonistes peuvent allouer l'intégralité de leurs ressources au service d'autres individus, au détriment complet de leur fitness propre, et ne considérant ainsi que leur fitness inclusive. L'origine de l'eusocialité et les théories relatives à celle-ci restent aujourd'hui encore sujet à débats parmi la communauté scientifique.
Chez les Vertébrés
Caractéristiques de la socialité chez les vertébrés :
- L'importance de la communication selon différents modes tels que des signaux sonores, chimiques ou visuels.
- Les comportements territoriaux comme la défense de ressources ou de territoires de reproduction.
- Des stratégies reproductrices bien déterminées : polygamie la plus répandue ; monogamie stricte quand le mâle pourvoit à la survie des jeunes (soins parentaux).
- Hiérarchie sociale
Le stade subsocial : exemple chez un lézard

Chez les reptiles, seules des formes rudimentaires de la socialité sont représentées. Les soins parentaux sont rarement observés ou se limitent essentiellement à la défense des œufs. Le genre Egernia présente la forme la plus complexe de socialité chez les lézards, avec la formation de groupes sociaux généralement constitués d'un couple monogame et de juvéniles descendants directs du couple monogame ou de copulations hors couple. Le groupe possède un territoire dont un domaine est alloué au mâle et l’autre à la femelle. La distribution spatiale des juvéniles dans le territoire mâle est fonction du taux d’apparentement avec celui-ci, les juvéniles eu par copulation hors couples se limitent au territoire de la mère. Ce comportement est un argument probant en faveur de la théorie de la sélection de parentèle.
Les soins parentaux sont soumis à la sélection car coûteux pour les parents, ils augmentent entre autres la compétition parents-enfants pour les ressources. Les mâles ont ainsi développé une capacité de reconnaissance des apparentés auxquels ils procurent préférentiellement les soins parentaux. Prolongés sur plus d’un an, ils entraînent un retard de dispersion bénéfique en terme fitness pour les juvéniles (accès aux ressources et protection contre les prédateurs). À l'état ancestral, les lézards sociaux devaient présenter une forte promiscuité, causant des paternités multiples très coûteuses pour les femelles car entraînant :
- Une augmentation de la prédation par le père social et donc une augmentation de l'investissement femelle dans la protection.
- Une réduction de la surface du domaine accessible à la progéniture hors-couple et donc des avantages du retard de dispersion pour la descendance.
- Une augmentation du coût pour les femelles du retard de dispersion en amplifiant la concurrence pour les ressources entre les mères et leur progéniture.
L'établissement de conflits parentaux sur la progéniture représente donc une pression de sélection importante pour le développement et le maintien de la socialité chez Egernia et probablement chez d'autres taxons aux systèmes sociaux semblables[11].
Le stade communal : exemple des marmottes

Il existe 14 espèces de marmottes dispersées à travers l’hémisphère nord qui présentent une grande variété de systèmes sociaux.
- Marmota monax mène une existence solitaire.
- Marmota flaviventris forme des groupes de femelles apparentées, avec leurs juvéniles (moins d’un an) et des jeunes d’un an (=yearling).
- Marmota olympus et Marmota caligata présentent un système social constitué d’un mâle avec deux femelles et d’une portée de juvéniles ou de yearlings.
- Toutes les autres espèces vivent en groupes familiaux constitués d’un couple reproducteur territorial, d’adultes subordonnés, de yearlings et de juvéniles.
La socialité chez les marmottes évolue avec deux caractéristiques cruciales de leur biologie: la taille du corps et l’hibernation. L’interaction entre ces deux composantes et les contraintes écologiques rencontrées par les différents individus peuvent avoir pour conséquences le retard de la maturité sexuelle, de l’âge à la première reproduction ou de la dispersion selon les espèces.
Si ces retards favorisent la formation de groupes sociaux d’apparentés, ils sont aussi à l’origine de coûts en termes de reproduction et donc de perte de fitness par suppression de la reproduction des individus matures les plus jeunes par les plus âgés. Certains jeunes n’ont ainsi aucune possibilité de se reproduire. Ces comportements sont néanmoins maintenus en vertu des bénéfices récoltés :
- la survie augmente chez les espèces ayant un retard de dispersion.
- les soins parentaux durant l’hibernation sont prolongés aussi longtemps que le groupe est formé.
Le coût des soins parentaux (temps et énergie alloués aux juvéniles) est compensé par la présence d’adultes subordonnés apparentés aux juvéniles qui aident leurs parents, avec pour bénéfice individuel une augmentation de la fitness inclusive. L’exemple des marmottes met ainsi en évidence le panel de comportements coopératifs et altruistes (par sélection de parentèle) qui ont évolué chez les mammifères[12].
L'eusocialité : le rat-taupe
L’eusocialité chez les vertébrés est rarement observée, il n'existe que deux espèces connues de mammifères eusociaux. Il s'agit du rat-taupe nu Heterocephalus glaber, et du rat-taupe de Damara Cryptomys damarensis.
Cas particuliers de socialité
À la suite des travaux de Darwin, la socialité a été décrite chez de nombreuses espèces, pas seulement animales. Aujourd'hui, les théories d'Hamilton et les recherches qui se poursuivent depuis se concentrent principalement sur des espèces animales et ne considèrent pas certains types d'échanges et de mécanismes coopératifs comme des évolutions sociales. Cependant, certaines publications notoires restent sujet à débats quant à l'emploi du terme de socialité à d'autres forme du vivant.
Le cas des bactéries

Chez les bactéries, plusieurs cas de coopération ont été étudiés et bien établis. Les concepts relevant de la socialité (au sens large) à savoir les bénéfices directs ou indirects entre les différents protagonistes sont applicables à ces organismes.
Ce n'est qu'à l'approche des années 2000 que sont présentés les premiers travaux sur la coopération et l'altruisme chez les microorganismes. En 2007, Pamela Lyon publie un historique des recherches sur l'évolution de la socialité chez les bactéries[13]. Loin de se limiter au système reproductif clonal, l'article discute des différentes théories de l'évolution sociale et du maintien des mécanismes coopératifs en s'appuyant sur les modèles bactériens, avançant un intérêt largement reconnu dans la communauté scientifique: ces modèles sont peu coûteux, facilement manipulable et intéressant pour tester des théories évolutives de par leur temps de génération extrêmement court.
Par exemple, Myxococcus xanthus a permis la mise en évidence d'une preuve empirique de socialité bactérienne. Ces Eubactéries se distinguent par la formation d'un "corps de fructification pluricellulaire", une sorte d'essaim de bactérie pour maximiser leur prise alimentaire. Ainsi, elles font preuve d'un mécanisme coopératif dans un environnement qui ne le requiert pas nécessairement, avec pour résultat un bénéfice net en termes de fitness pour chaque individu. Certaines expériences ont aussi montré qu'un tricheur dans ce système coopératif maximiserait sa fitness (relativement au groupe coopératif) sans envahir nécessairement le groupe. Cet exemple est très démonstratif quant à l'application des modèles sociaux précédemment abordés aux bactéries et reste prometteur pour la vérification des nombreuses théories émergentes ou revisitées en écologie évolutive.
Le cas particulier des plantes
Bien que de nombreux mécanismes coopératifs aient été décrits chez les plantes. L'emploi de socialité chez ces organismes semble plus problématique. Quels sont les bénéfices de ces mécanismes ? à qui sont-ils adressés ?
M.Heil et R.Karban publient en 2010 un article synthétique des connaissances actuelles sur un de ces mécanismes les plus observés chez les végétaux et plus particulièrement les arbres[14] : la communication moléculaire. En réponse à une pression de prédation ou à un parasitisme prononcé, certains arbres, comme le saule nord-américain Salix sitchensis, expriment un niveau de résistance nettement supérieur à leurs semblables. Un certain nombre de composés volatils organiques synthétisés par des arbres subissant ou ayant également subi une pression ont été identifiés comme inducteurs de ces niveaux de résistance. Si de nombreux modèles théoriques s'intéressent aux liens entre émetteurs et receveurs de ses signaux, aucun bénéfice net n'a été isolé pour les premiers. La sélection de parentèle a bien entendu été évoquée mais, fait intéressant, les émetteurs de ces signaux moléculaires ne sont pas nécessairement de la même espèce que les individus avec lesquels ils "coopèrent", autrement dit, la nature de ces signaux n'est pas spécifique. Ces recherches sont néanmoins freinées par la nature des données collectées jusqu'à aujourd'hui : celles-ci sont principalement centrées sur la cascade de réaction du receveur et les bénéfices tirés.
La socialité n'a donc toujours pas été mise en évidence chez ces organismes, mais le champ de recherche reste ouvert pour résoudre cette problématique : pourquoi une réaction qui ne semble pas bénéfique en termes de fitness chez les émetteurs est maintenue ?
Débats et controverses témoins d'une science émergente
Les théories sur l'évolution de la socialité s'affinent continuellement. Bien que les 3 concepts évoqués ici restent des modèles de base, il existe aujourd'hui de nombreuses variations sur ces thèmes, tant du point de vue théorique qu'empirique.
En ce qui concerne le mutualisme, plusieurs modèles expérimentaux montrent aujourd'hui que la coopération peut facilement évoluer au niveau inter-spécifique. La sélection de parentèle et la théorie de la fitness inclusive forment un modèle fondamental pour justifier l’altruisme. Cette théorie et son influence sur l’évolution de l’altruisme, considérée par beaucoup comme une découverte majeure de l’écologie évolutive reconnue par la grande majorité de la communauté scientifique, bien qu’elle fasse aujourd’hui encore l’objet de débats quant à son interprétation et la modélisation des données [15] - [16]. Celui initié par Nowak et al[15] - [16]. illustre bien ces différences d’interprétation. Selon Nowak, la théorie de la fitness inclusive (et par extension la portée de la sélection de parentèle) est limitée par le « conditionnement strict » de son modèle mathématique, trop simplificateur et qui laisserai de côté des facteurs important comme la compétition. Ses détracteurs (Patrick Abbot and al.) lui reprochent sa distinction brutale de la « théorie de la sélection naturelle classique » et de celle de la fitness inclusive ainsi qu’une confusion dans la compréhension du modèle mathématique[16].
La théorie d’Hamilton a souvent été argumentée sur l’appui de l’haplodiploïdie, présentée comme étant à l’origine de comportements coopératifs chez les insectes eusociaux. Cependant, de nombreux contre-exemples s’opposent à cet argument : d’abord parce que l’haplodiploïdie reste un système reproductif rare même parmi les espèces sociales, et surtout parce que la majorité des espèces haplodiploïdes (comme la cochenille par exemple) sont et restent solitaires.
La théorie de la manipulation parentale (Alexander 1974) revient de manière périodique comme alternative à l’haplodiploïdie. D’après lui, s’appuyant sur l’exemple des fourmis, la reine sous alimente ses filles dans le but de réduire leur potentiel reproducteur et les domine (agressions fréquentes) pour rendre impossible une fondation indépendante. Un éventuel gène rebelle serait contre-sélectionné car il deviendrait désavantageux quand le rebelle deviendrait lui-même parent. Une partie de la communauté scientifique (parmi lesquels S. Aron) la décrient en raison de sa faible valeur prédictive. Selon eux, l’évolution aura favorisé la sélection de contre-stratégies dans le chef des filles, leur permettant d’échapper à l’emprise royale. La stérilité des ouvrières – totale ou partielle – s’explique aujourd'hui très bien par la sélection de parentèle.
Ces quelques exemples sont témoins des problématiques actuelles rencontrées lorsqu'est abordée l'évolution de la socialité. Les théories explicatives sont toutes relativement récentes, différents modèles sont proposés et encore aujourd'hui discutés.
Notes et références
- D. C. Queller, 2001, W. D. Hamilton and the evolution of sociality, Behaviour Ecology Vol. 12 No. 3: 261-268
- M. Nowak & al., 2010, The evolution of eusociality, Nature 466; 1057–1062
- Tim Clutton-Brock, 2002, Breeding Together: Kin Selection and Mutualism in Cooperative Vertebrates, Science n°296, 69
- Joel S. Brown et Thomas L. Vincent, 2008, Evolution of cooperation with shared costs and benefits, Proceedings: Biological Sciences, Vol. 275, No. 1646, pp. 1985-1994
- Leticia Avilés, 1999, Cooperation and non-linear dynamics: An ecological perspective on the evolution of sociality, Evolutionary Ecology Research, 1: 459–477
- Hamilton W. D., 1964, The genetical evolution of social behaviour, Journal of Theorical Biology, no. 7
- RL Trivers, H Hare, 1976, Haplodiploidy and the Evolution of Social Insects, Science, New Series, Vol. 191 no. 4224 p. 249-263
- D.S. Wilson & E. Sober, 1994, Reintroducing group selection to the human behavioral sciences. Behavioral and Brain Sciences 17 (4): 585-654.
- (en) Jean Vannier, Muriel Vidal, Robin Marchant, Khadija El Hariri, Khaoula Kouraiss et al., « Collective behaviour in 480-million-year-old trilobite arthropods from Morocco », Scientific Reports, vol. 9, , article no 14941 (DOI 10.1038/s41598-019-51012-3).
- Bryan N. Danforth, 2002,Evolution of sociality in a primitively eusocial lineage of bees, 286–290.
- Geoffrey M. While, Tobias Uller et Erik Wapstra, 2009, Family conflict and the evolution of sociality in Reptiles Behavior Ecology, 20:245-250
- Kenneth B. Armitage, 1999, Evolution of sociality in marmots, Journal of Mammalogy, Vol. 80, No. 1, pp. 1-10
- Pamela Lyon, 2007, From quorum to cooperation: lessons from bacterial sociality for evolutionary theory, Biol. & Biomed. Sci. 38 820–833, doi:10.1016/j.shpsc.2007.09.008
- M.Heil & R.Karban, 2010, Explaining evolution of plant communication by airborne signals, Trends in Ecology and Evolution, vol 25 n°3, 137-144
- Martin A. Nowak, Corina E. Tarnita & Edward O. Wilson, 2010, The evolution of eusociality, Nature 466; 1057–1062
- Patrick Abbot and al., 2011, Inclusive fitness theory and eusociality, Nature 471, E1-E4;