Trilobita
Les trilobites (Trilobita) constituent une classe d'arthropodes marins fossiles ayant existé durant le Paléozoïque (ère primaire) du Cambrien au Permien. Les derniers trilobites ont disparu lors de l'extinction de masse à la fin du Permien, il y a 250 Ma.

| Règne | Animalia |
|---|---|
| Sous-règne | Eumetazoa |
| Clade | Protostomia |
| Super-embr. | Ecdysozoa |
| Embranchement | Arthropoda |
Ordres de rang inférieur
- † Agnostida Salter, 1864
- † Asaphida Fortey & Chatterton 1988
- † Corynexochida Kobayashi 1935
- † Harpetida Ebach & Mcnamara 2002
- † Lichida Moore 1959
- † Odontopleurida Whittington 1959
- † Phacopida Salter 1864
- † Proetida Fortey & Owens 1975
- † Ptychopariida Swinnerton 1915
- † Redlichiida Richter, 1932[1]
Les trilobites variaient en taille, entre 1 millimètre et environ 70 cm de long[2] - [3] - [4]. Leur taille moyenne se situe cependant entre 2 et 7 centimètres. Bien connus, ils sont très abondants dans les roches datant de l'ère primaire et très appréciés des collectionneurs de fossiles par leur beauté et leur variété de formes. Plus de 18 750 espèces[5] ont été décrites chez cette classe d'arthropodes, ce qui en fait l'un des groupes exclusivement fossiles les plus diversifiés.
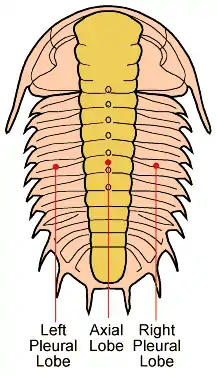
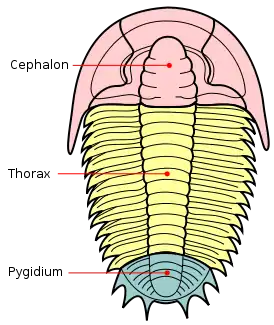
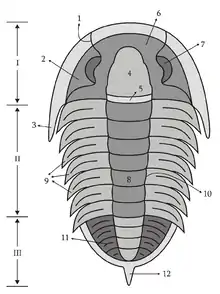
Les trilobites forment leur propre groupe défini par leur anatomie spécifique, caractérisée par la « trilobation » longitudinale (lobe médian ou rachis, et deux lobes latéraux ou plèvres) et métamérique (céphalon ou « tête », thorax et pygidium ou « queue »). C'est l'origine de leur nom. Certains crustacés (cloportes isopodes, triops) ou certains chélicérates (limules) peuvent leur ressembler superficiellement, mais n'en font pas partie, même si ce sont, eux aussi, des arthropodes.
Anatomie
Anatomie dorsale
Le céphalon
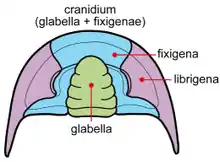
Le céphalon se compose de cinq (peut-être six) segments que l'on rassemble sous le terme de glabelle. Elle peut présenter trois types de morphologie : pointue (rétrécie vers l'avant), arrondie ou sphérique (morphologie archaïque) et élargie vers l'avant. Le segment le plus en avant forme l'anneau occipital, il joue un rôle important dans la détermination des trilobites. Parmi les segments suivants on trouve : un segment portant les yeux composés, un autre portant une paire d'antennes uniramées (i.e. à une seule branche) et trois segments portant chacun une paire de segments biramés (i.e. à deux branches).
La glabelle est bordée par les joues. Elles sont traversées par une ligne de suture faciale qui permet de distinguer les joues fixes (fixigènes - intérieur) des joues libres (librigènes - extérieur). C'est aussi au niveau de cette ligne de suture que l'on trouve les yeux en forme de croissant lesquels pouvaient être particulièrement évolués (ou absent chez les espèces vivant sous la zone euphotique). Ces yeux sont composés de prismes de calcite (jointifs ou non), ces yeux ne pouvaient donc pas réaliser d'accommodation (mais avaient une grande profondeur de champ et une faible aberration). En fait, les trilobites furent les premiers animaux à développer de vrais yeux, il y a de ça 543 Ma.
On distingue deux types d'yeux :
- Les yeux de type holochroale se caractérisent par une cornée recouvrant toutes les ommatidies dont le nombre est estimé entre 100 et 15 000. Les images de chaque ommatidie se superposent créant une image en relief mais le champ de vision de chaque œil est réduit.
- Les yeux de type schizochroale diffèrent par une individualisation des ommatidies. Chaque unité est ainsi séparée par du tissu conjonctif et elle possède sa propre cornée. Cette configuration conduit aussi à vue par opposition des images car chaque ommatidie ne voit qu'un fragment mais elle permet aussi au trilobite de couvrir un champ de vision supérieur à 180°. Ce type d'œil ne se rencontre que dans l'ordre des Phacopida, il semble qu'il soit adapté pour des environnements de faible luminosité.
Le thorax
Il a pour caractéristique d'être composé de segments articulés les uns avec les autres (segments mobiles). Le nombre de segments dans cette région du corps varie de 2 à 103 (Balcoraccania dailyi[6]). Un trilobite possédant un nombre inférieur ou égal à 3 segments est qualifié de myomera, s'il possède plus de 3 segments il est qualifié de polymera. Les segments du thorax forment parfois deux ensembles morphologiques distincts chez certaines formes primitives. On parle alors de prothorax, pour la partie antérieure, et d'opisthothorax, pour la partie postérieure. Le nombre élevé de segments offre une grande souplesse au thorax favorisant ainsi une position de défense par enroulement comme on peut l'observer chez d'autres arthropodes.
Le pygidium
Le pygidium se compose chez la plupart des trilobites de segments identiques aux segments thoraciques mais ne s'articulant pas entre eux (segments soudés). L'ensemble rigide ainsi formé semble avoir été utilisé comme palette natatoire et peut être de taille très différente selon les espèces. On parle de forme micropyge, subisopyge, isopyge ou macropyge selon que le pygidium est de taille respectivement très inférieure, un peu inférieure, à peu près identique, ou largement supérieure à celle du céphalon. La plupart des formes primitives de trilobites sont de type micropyge. Des extensions non segmentées (épines pygidiales) sont quelques fois observées aux extrémités du pygidium, on distingue le telson (une seule pointe) et les cerques (pointes multiples).
L'exosquelette
Les trilobites n'étaient protégés qu'en surface, mais ils possédaient un solide exosquelette. Durant la mue, cet exosquelette se scindait en deux au niveau de la jonction tête/thorax, si bien que de nombreux fossiles sont dépourvus de l'une ou l'autre de ces parties du corps. En effet, de nombreux fossiles sont des résidus de la mue plutôt que des trilobites morts. Dans la plupart des groupes, on trouvait deux sutures faciales (articulations sans cartilages et peu mobiles) sur les joues, ce qui facilitait la mue.
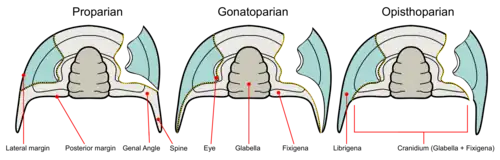
La géométrie de la suture faciale participe à la systématique des trilobites, on distingue :
- Suture proparienne : elle aboutit avant les pointes génales (genre Phacops).
- Suture gonatoparienne : elle aboutit dans les angles (genre Asaphiscus).
- Suture opisthoparienne : elle aboutit à l'arrière du céphalon.
- Suture hypoparienne : elle n'est plus visible en vue dorsale car elle se situe sur la bordure du bouclier céphalique. On la rencontre surtout chez des individus primitifs tels que ceux appartenant au genre Olenellus).
Chez certains trilobites, comme ceux de l'ordre Lichida, l'exosquelette a évolué vers des formes élaborées en épines, de l'Ordovicien jusqu'à la fin de la période du Dévonien. On a trouvé de tels fossiles dans la formation du Hamar Laghad d'Alnif, au Maroc. De nombreux faux de ce type de fossiles ont été produits. D'autres spécimens ont été retrouvés dans l'est de la Russie et en Ontario, au Canada. D'après le New Scientist magazine (), « certains [...] trilobites [...] avaient des cornes sur la tête, similaires à celles des scarabées actuels. » En se fondant sur leur localisation, leur forme et leur taille, le biologiste londonien Rob Knell et son collègue Richard Fortey, du Muséum d'histoire naturelle de Londres, ont avancé que ces cornes servaient probablement de dispositif d'attaque aux trilobites, lors de combats visant à départager des prétendants à un accouplement. Cela ferait des trilobites l'un des tout premiers groupes à avoir manifesté ce comportement, et bien que l'étude n'ait couvert que la famille des raphiophoridae, les conclusions peuvent certainement s'appliquer à d'autres groupes.
Anatomie ventrale

Les trilobites étaient pourvus d'une paire d'antennes préorales et de membres biramés indifférenciés. Ces derniers se rencontraient uniquement sur le thorax et le pygidum. Seul un segment (précoxa) relie l'appendice au reste du corps. La partie locomotrice du membre (l'endopodite) est constitué de sept segments, elle est reliée au précoxa par l'intermédiaire du coxa. La partie branchiale est représentée par le préépipodite, il est directement relié au précoxa. Les membres étaient tous protégés sur leurs côtés par les lobes pleuraux, disposés symétriquement autour d'un lobe axial.
Anatomie interne
- Les principaux organes de l’appareil digestif étaient contenus dans le rachis ; un hypostome vers l’avant et une petite pièce médiane ou épistome encadrent la bouche. L’œsophage, l’estomac et les glandes secrétant des sucs digestifs logent dans la glabelle où l’on trouve leurs empreintes. Les coecums ou ceca présentent une disposition métamérique, mais le segment oviparial en est dépourvu.
- Le système musculaire des trilobites est mal connu mais il est probable que les muscles étaient striés comme chez les autres arthropodes.
- Le système nerveux, proche des autres arthropodes, se compose presque toujours d’un gros ganglion cérébral, de nerfs optiques conduisant aux yeux composés, d’un collier péri-œsophagien, d’un ganglion sous-œsophagien et d’une chaîne nerveuse ventrale.
- Comme chez les autres arthropodes, l’appareil circulatoire s’articule autour d’un épais vaisseau dorsal composé de « cœurs » emboîtés pompant d’arrière en avant un sang que l’on suppose incolore et à base de cuivre (hémocyanique) comme celui des autres arthropodes.
Développement
Le développement des trilobites est particulièrement connu, et ce grâce à une minéralisation précoce de leur carapace dorsale ayant permis la fossilisation de très jeunes individus. Il est généralement subdivisé en trois périodes :
- La période protaspis (« larvaire ») ;
- La période meraspis (« juvénile ») ;
- La période holaspis (« adulte »).
Écologie
Modes de vie

Les trilobites étaient exclusivement marins. La très grande majorité d'entre eux étaient benthiques, ils vivaient sur (épibenthique) ou dans (endobenthique) le substrat constituant le fond des mers. Leurs activités ont laissé d'importantes traces. Les Bilobites (deux bourrelets subparallèles et striés) ont longtemps été considérés comme le déplacement du trilobite sur le sédiment, chaque lobe correspondant à un rang d'appendice, mais ils sont désormais interprétés « en paléo-ichnologie comme les empreintes de déplacement de gros vers marins à soies rigides[7] ». Les Rhizophicus ou "Pas de bœuf" résultent de l'enfouissement, du fouissage voire de traces de repos de trilobite dans le sédiment.
Il semble toutefois que certaines formes aient été pélagiques, c'est-à-dire vivant dans la masse d'eau en se déplaçant de façon active (i.e. nectonique; e.g. Cyclopyge) ou passive (i.e.planctonique), peut-être le cas des Agnostida). L'adaptation au mode de vie pélagique s'est accompagnée de modifications de l'exosquelette : les pointes génales atteignent de grandes tailles, permettant la stabilisation du corps et les yeux sont très développés. Certaines formes étaient nectobenthiques : elles ne vivaient pas sur le fond mais à proximité du fond. Elles se caractérisaient par l'absence de pointes genales et un corps plat et très grand limitant ainsi les turbulence hydrodynamiques.
Les fossiles de trilobites Ampyx priscus trouvés au Maroc, dans des couches sédimentaires âgées d'environ 480 Ma, témoignent d'un comportement collectif. Ils sont en file linéaire tout en gardant entre eux des contacts étroits via leurs très longues épines[8]
Régimes alimentaires
La majorité des premiers trilobites tels que l'Olenoïdes étaient probablement les prédateurs des invertébrés benthiques, tels que des vers (Ottoia). Leurs pattes étaient munies de gnathobases épineuses. Le ver était extrait de sa galerie puis écrasé ou déchiré par les épines des pattes et les puissantes gnathobases, puis vraisemblablement amené entre les pattes vers la bouche où il était broyé au niveau de l'hypostome avant l'ingestion. Dans les crustacés et les insectes, toutes ces fonctions sont remplies par les trompes antérieures spécialisées dans la transformation de la nourriture avant l'ingestion. Cependant, la majeure partie du traitement s'effectuait dans la cannelure médiale longitudinale entre les membres avec leurs successions de paires de gnathobases.
Les trilobites prédateurs devaient avoir les hypostomes conterminants fermement attachés à la doublure frontale, stabilisant essentiellement l’hypostome contre l'exosquelette céphalique afin d’améliorer le traitement de la proie. Il y a une variation considérable de la taille et de la forme de l’hypostome conterminant, suggérant les nombreuses spécialisations trophiques.
Reproduction
Ce n'est que depuis 2017 qu'on sait de façon certaine que les trilobites pondaient des œufs, après la découverte d’œufs fossiles près de trilobites très bien conservées, dans des roches de l'État de New York vieilles de 450 Ma[9]. Ces œufs ont un diamètre de presque 200 µm. Les trilobites pourraient avoir déposé leurs œufs et leur sperme via des pores génitaux situés à l'arrière de leur tête.
Évolution

Les trilobites apparaissent véritablement en grand nombre au Cambrien inférieur, il y a environ 525 millions d'années[10]. Ces premiers trilobites appartiennent tous à l'ordre des Redlichiida, mais très vite les ordres des Agnostida et des Ptychopariida apparaissent. Le Cambrien est une période importante de diversification pour les trilobites. Ils atteindront d'ailleurs leur maximum de diversité au Cambrien supérieur. L'Ordovicien est marqué par un renouvellement très net des faunes à trilobites dû à l'apparition de nouveaux groupes, en particulier les Asaphida et les Corynexochida. Durant cette période, les trilobites présentent encore une très forte diversité, mais celle-ci chute brutalement lors de l'extinction de masse fini-ordovicienne vraisemblablement engendrée par une importante glaciation.
À la suite de cette crise, les trilobites se diversifient à nouveau, lentement au Silurien, puis de façon plus importante au Dévonien inférieur. Le Dévonien supérieur est néanmoins marqué par une suite d'événements d'extinctions parmi lesquels le 'Kellwasser supérieur', au passage Frasnien/Famennien, et le Hangenberg, au Famennien terminal, constituent des crises majeures de l'histoire de la vie. Des cinq ordres présents au début du Dévonien supérieur, seuls les Phacopida et les Proetida survivent à l'événement 'Kellwasser supérieur'. Puis les Phacopida s'éteignent lors de l'événement 'Hangenberg', ce qui n'empêchera pas les Proetida de survivre encore pendant près de 90 millions d'années. Après une phase importante de diversification au Carbonifère inférieur, la diversité des trilobites chute de façon importante au début du Carbonifère supérieur. Les trilobites ne seront plus alors qu'une composante mineure des faunes marines benthiques, jusqu'à leur disparition à fin du Changhsingien, l'étage géologique le plus récent du Permien, lors de la plus importante crise biologique de l'histoire de la vie.
Intérêts
Stratigraphie
Ces fossiles ont eu une vaste répartition géographique, avec un grand nombre d'espèces évoluant rapidement, ce qui permet de dater les formations géologiques de façon précise. C'est pourquoi les trilobites sont de bons fossiles stratigraphiques, dont on se sert pour créer les divisions chronologiques dans le Paléozoïque (surtout du Cambrien au Dévonien).

En France, on trouve des trilobites en Languedoc, dans les Pyrénées, en Bretagne/Normandie et dans les Ardennes[11]. Les plus anciens ont été découverts dans les départements de l'Hérault, l'Aude et l'Aveyron avec des spécimens datés du Cambrien inférieur. Du Cambrien, à l'Ordovicien et au Dévonien, le Pays de Galles, la Bretagne, la Bohême (République tchèque), la Russie, les Appalaches aux États-Unis et le Maroc (en particulier la région d'Erfoud) sont quelques-unes des régions où les gisements de trilobites sont parmi les plus importants et les plus diversifiés au monde.
À partir du Carbonifère, leur effectif et leur diversité tend à diminuer jusqu'à leur disparition à la fin du Permien, définissant la limite supérieure du Paléozoïque (ère primaire).
Paléogéographie

Au cours du Paléozoïque, les terres émergées se concentraient en trois grandes masses : Gondwana, Laurussia et Sibéria, les trilobites, essentiellement benthiques, se répartissaient alors sur les plateaux continentaux. Toutefois le caractère benthiques de leur mode de vie ne leur permet pas une forte propagation géographique. Ce manque de mobilité entraîne un fort provincialisme avec la création de trois provinces à trilobites. Ces dernières ne sont toutefois pas corrélées avec les masses continentales car l'état larvaire favorise leur dissémination et permet ainsi leur implantation sur d'autres rives. Mais la convergence des masses continentales pour former la Pangée favorise le rapprochement et le mélange des faunes et par là même le développement de la concurrence, ce qui induira la disparition de plusieurs espèces.
Paléoenvironnement
L'environnement des trilobites était constitué d'algues et de coraux primitifs qui ont disparu lors de la crise Permien/Trias. Ils étaient les proies de divers prédateurs tel Anomalocaris[12] ; on a retrouvé des traces de morsure.
Collections

Leur abondance dans certaines séries sédimentaires ainsi que leur diversité de formes et de tailles ont fortement contribué à populariser les trilobites auprès des collectionneurs. Cet intérêt a aussi contribué à favoriser leur étude par les paléontologues au point qu'ils ont fait l'objet de plus de travaux que l'ensemble des autres groupes d'invertébrés.
Cependant leur intérêt ne se cantonne pas à l'histoire récente. Une amulette sculptée à partir d'un trilobite datant du Silurien a été retrouvée dans un refuge rocheux, par la suite nommé Grotte du Trilobite en référence à l'objet, à Arcy-sur-Cure (Bourgogne). Les sédiments ont permis de dater cet objet du Magdalénien (15 000 ans av. J.C.). En Australie, un ustensile a été confectionné par des aborigènes à partir d'un trilobite du Cambrien.
Dans l'ouest de l'Utah, les Indiens de la tribu Utes extraient des trilobites Elrathia kingi qu'ils trouvent en abondance dans les séries du Cambrien moyen à House Range et les façonnent en amulettes. Dans leur langue, cette espèce est nommée timpe khanitza pachavee, ce qui signifie "petite punaise d'eau dans une maison de pierre".
Systématique
La classe des Trilobita a été décrite par le naturaliste allemand Johann Ernst Immanuel Walch en 1771[13].
Liste des ordres
D'après la synthèse du site du Dr. Sam Gon III[1] :
- Agnostida Salter, 1864
- Asaphida Fortey & Chatterton 1988
- Corynexochida Kobayashi 1935
- Harpetida Ebach & Mcnamara 2002
- Lichida Moore 1959
- Odontopleurida Whittington 1959
- Phacopida Salter 1864
- Proetida Fortey & Owens 1975
- Ptychopariida Swinnerton 1915
- Redlichiida Richter, 1932
_-_6%252C8_mm_1.JPG.webp)
 Asaphus lepidurus (Asaphida).
Asaphus lepidurus (Asaphida).
.jpg.webp) Harpes perradiatus (Harpetida).
Harpes perradiatus (Harpetida). Hoplolichas (Lichida).
Hoplolichas (Lichida). Radiaspis (Odontopleurida).
Radiaspis (Odontopleurida). Phacops rana (Phacopida).
Phacops rana (Phacopida). Monodechenella macrocephala (Proetida).
Monodechenella macrocephala (Proetida). Elrathia kingii (Ptychopariida).
Elrathia kingii (Ptychopariida). Acadoparadoxides briareus (Redlichiida).
Acadoparadoxides briareus (Redlichiida).
Notes et références
- (en) Dr. Sam Gon III, « Systematic Listing of Trilobite Families », sur trilobites.info (consulté le ).
- Rapport d'activités – voir les images d’un trilobite géant sur GIG (centre géologique de Canelas, Portugal), Mars 2007
- Article sur le journal Público, le 2 juin 2007 (pt)
- Sea Anelid Fossils, pages 10 et suivantes sur Penichefossil
- J.M. Adrain 2006. Higher taxa as proxies for species diversity: a global species database of Trilobita. Geological Society of America, Abstracts with Programs, 38(7): 207.
- J.R. Paterson & G.D. Edgecombre 2006. The Early Cambrian trilobite family Emuellidae Pocock, 1970 : systematic position and revision of Australian species. Journal of Paleontology, 80(3), pp.496-513.
- Claude Augris, Atlas thématique de l'environnement marin de la baie de Douarnenez (Finistère), éditions Quae, , 135 p. (lire en ligne).
- Futura, « Des trilobites avaient déjà des comportements collectifs il y a 480 millions d’années », sur Futura (consulté le )
- (en) Thomas A. Hegna et al., « Pyritized in situ trilobite eggs from the Ordovician of New York (Lorraine Group) : Implications for trilobite reproductive biology », Geology, , article no G38773.1 (DOI 10.1130/G38773.1).
- (en) Dr. Sam Gon III, « Trilobite Systematic Relationships », sur trilobites.info (consulté le )
- La Réserve naturelle de Vireux-Molhain, plus connue sous le nom de Mur des Douaniers, est un affleurement de schistes d'où ont été extraits de très nombreux spécimens du Dévonien moyen.
- Cette hypothèse est cependant toujours discutée. Voir, par exemple, cet article.
- Walch, Die Naturgeschichte der Versteinerungen zur Erläuterung der Knorrischen Sammlung von Merkwürdigkeiten der Natur. Nürnberg 1771
Voir aussi
Articles connexes
Références taxonomiques
- (en) Référence World Register of Marine Species : taxon Trilobita Walch, 1771 † (+ liste ordres + liste familles)
- (en) Référence Paleobiology Database : Trilobita Walch 1771 †
- (en) Référence Arthropoda Species Files : Trilobita †
- (en) Référence Tree of Life Web Project : Trilobita
- (en) Référence BioLib : Trilobita Walch, 1771 †
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023
- (en) Référence Catalogue of Life : Trilobita † (consulté le )
Liens externes
- A Guide to the Orders of Trilobites (en): site très complet sur tous les aspects des trilobites.
- Luc Van Bellingen, « Les trilobites »