Carbone bleu
Le carbone bleu fait référence au dioxyde de carbone retiré de l'atmosphère par les écosystèmes océaniques côtiers du monde, principalement les mangroves, les marais salés, les herbiers marins et potentiellement les macroalgues, par la croissance des plantes et l'accumulation et l'enfouissement de matière organique dans le sol.
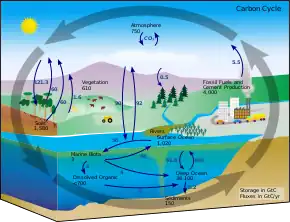
Puits de carbone
Plus de la moitié (55 %) du carbone biologique capturé mondialement l'est par des organismes marins vivants[1] - [2].
Historiquement, l'océan, l'atmosphère, le sol et les écosystèmes forestiers terrestres ont été les plus grands puits de carbone naturels. Les recherches sur le rôle des écosystèmes côtiers végétalisés ont mis en évidence leur efficacité potentielle en tant que puits de carbone[2]. Elles ont conduit à la reconnaissance scientifique du terme «carbone bleu» pour désigner le carbone qui est fixé via les écosystèmes océaniques côtiers, plutôt que par les écosystèmes terrestres traditionnels, comme les forêts[3]. Bien que les habitats végétalisés de l'océan couvrent moins de 0,5% des fonds marins, ils sont responsables de plus de 50%, et potentiellement jusqu'à 70%, du stockage de carbone dans les sédiments océaniques. Les mangroves, les marais salés et les herbiers marins constituent la majorité des habitats végétalisés de l'océan, mais ne représentent que 0,05% de la biomasse végétale terrestre. Ils peuvent stocker une quantité comparable de carbone par an et sont des puits de carbone très efficaces en séquestrant le carbone dans leurs sédiments sous-jacents, dans la biomasse souterraine et dans la biomasse morte[2] - [4].
Le carbone bleu peut être séquestré pendant des millions d'années dans les sédiments végétaux sous-jacents issus des feuilles, des tiges, des branches ou des racines. Les estimations de la capacité d'enfouissement du carbone bleu à long terme sont encore incertaines[4]. Bien que les écosystèmes côtiers végétalisés couvrent moins de surface et aient moins de biomasse aérienne que les plantes terrestres, ils peuvent avoir un impact sur la séquestration à long terme du carbone, en particulier dans les puits de sédiments[3]. L'une des principales préoccupations est que ces importants écosystèmes marins disparaissent plus vite que tout autre écosystème de la planète. Les estimations actuelles suggèrent une perte de 2 à 7% par an, ce qui est non seulement une perte de séquestration de carbone, mais également une perte d'habitat qui est importante pour la gestion du climat, de la protection des côtes et de la santé.
Types d'écosystèmes de carbone bleu
Herbiers

Les herbiers marins sont un groupe d'une soixantaine d'espèces d'angiospermes qui se sont adaptées à la vie aquatique et peuvent pousser dans les prairies le long des côtes de tous les continents sauf de l'Antarctique[5]. Les prairies sous-marines se forment jusqu'à des profondeurs maximales de 50 m, selon la qualité de l'eau et la disponibilité de la lumière, et peuvent inclure jusqu'à 12 espèces différentes par prairie. Ces herbiers marins sont des habitats hautement productifs qui fournissent de nombreux services écosystémiques, notamment la stabilisation des sédiments, l'habitat et la biodiversité, une meilleure qualité de l'eau et la séquestration du carbone et des nutriments[6]. La superficie des herbiers répertoriés est de 177 000 km 2, mais on estime que la superficie réelle se situe entre 300 000 à 600 000 km 2 [7]. Bien que les herbiers ne représentent que 0,1% de la surface du plancher océanique, ils représentent environ 10 à 18% de l'enfouissement total du carbone océanique[8]. Actuellement, les herbiers marins mondiaux stockent jusqu'à 19,9 Pg (gigatonnes ou milliards de tonnes) de carbone organique. Le carbone s'accumule principalement dans les sédiments marins, qui sont anoxiques et préservent donc le carbone organique à des échelles de temps décennales, centennales et millénaires. Des taux d'accumulation élevés, un faible taux d'oxygène, une faible conductivité des sédiments et des taux de décomposition microbienne plus lents encouragent l'enfouissement et l'accumulation de carbone dans ces sédiments côtiers. Alors que les habitats terrestres perdent leur stock de carbone lors de la décomposition ou lors de perturbations comme les incendies ou la déforestation, les puits de carbone marins peuvent conserver le carbone pendant des périodes beaucoup plus longues. Les taux de séquestration du carbone dans les herbiers marins varient en fonction des espèces, des caractéristiques des sédiments et de la profondeur des habitats, mais en moyenne, le taux d'enfouissement du carbone est d'environ 138 g de carbone par m2 et par an[4]. Les habitats des herbiers marins sont menacés par l'eutrophisation côtière, l'augmentation des températures de l'eau de mer, l'augmentation de la sédimentation et du développement côtier, et l'élévation du niveau de la mer qui peuvent diminuer la lumière disponible pour la photosynthèse. La perte d'herbiers marins s'est accélérée au cours des dernières décennies, passant de 0,9% par an avant 1940 à 7% par an en 1990, avec environ 1/3 des pertes mondiales depuis la Seconde Guerre mondiale[9].
Mangrove

Les mangroves sont des halophytes ligneuses qui forment des forêts intertidales et fournissent de nombreux services écosystémiques comme la protection côtière, les zones d'alevinage pour les poissons et les crustacés côtiers, les produits forestiers, les loisirs, la filtration des nutriments et la séquestration du carbone[10]. On trouve des mangroves dans 105 pays différents, à Hong Kong et Macao, en Martinique, Guyane, Guadeloupe, Mayotte et dans le Somaliland, le long des côtes dans les eaux subtropicales et tropicales, selon la température, les précipitations, les marées, les vagues et le débit d'eau[11]. Elles poussent à l'intersection entre la terre et la mer, ont des composants semi-terrestres et marins, et comprennent des adaptations uniques, notamment des racines aériennes, des embryons vivipares et des mécanismes de rétention des nutriments très efficaces[12]. À l'échelle mondiale, les mangroves ont stocké 4,19 ± 0,62 Pg de carbone en 2012, l'Indonésie, le Brésil, la Malaisie et la Papouasie-Nouvelle-Guinée représentant plus de 50% du stock mondial. 2,96 ± 0,53 Pg de ce stock sont contenus dans le sol et 1,23 ± 0,06 Pg dans la biomasse vivante. De ces 1,23 Pg, environ 0,41 ± 0,02 Pg se trouvent dans la biomasse souterraine dans le système racinaire et environ 0,82 ± 0,04 Pg se trouvent dans la biomasse vivante aérienne[13].
On estime que, au niveau mondial, la canopée des mangroves couvre entre 83 495 km 2 et 167 387 km 2 en 2012, l'Indonésie contenant environ 30% de l'ensemble de la superficie mondiale des forêts de mangroves[14]. Les forêts de mangroves sont responsables d'environ 10% de l'enfouissement mondial du carbone[15] avec un taux d'enfouissement du carbone estimé à 174 g de carbone par m 2 et par an[12]. Les mangroves, comme les herbiers marins, ont un potentiel élevé de séquestration du carbone. Elles représentent 3% de la séquestration mondiale de carbone par les forêts tropicales et 14% de l'enfouissement de carbone de l'océan côtier mondial[11]. Les mangroves sont naturellement perturbées par les inondations, les tsunamis, les tempêtes côtières comme les cyclones tropicaux, la foudre, les maladies et les ravageurs, et les changements dans la qualité ou la température de l'eau. Bien qu'elles résistent à bon nombre de ces perturbations naturelles, elles sont très sensibles aux impacts humains, notamment le développement urbain, l'aquaculture, l'exploitation minière et la surexploitation des mollusques, crustacés, poissons et du bois [16]. Les mangroves fournissent des services écosystémiques et une séquestration du carbone d'importance mondiale et sont donc un habitat important à conserver et à réparer lorsque cela est possible.
Les marais salés

Les marais, écosystèmes intertidaux dominés par la végétation herbacée, se trouvent des côtes de l'Arctique à celles des régions subtropicales tandis que sous les tropiques, les mangroves dominent [17]. Les marais ont une productivité élevée, avec une grande partie de la production primaire de biomasse souterraine. Cette biomasse souterraine peut former des dépôts jusqu'à 8 m de profondeur. Les marais offrent un habitat précieux aux plantes, aux oiseaux et aux alevins, protègent l'habitat côtier des ondes de tempête et des inondations et peuvent réduire la charge en nutriments dans les eaux côtières[18]. À l'instar des mangroves et des herbiers marins, les marais salés constituent également d'importants puits de carbone. Les marais séquestrent le carbone dans la biomasse souterraine en raison des taux élevés de sédimentation organique et de décomposition anaérobie[19]. Les marais salés couvrent environ 22 000 à 400 000 km 2 dans le monde, avec un taux d'enfouissement du carbone estimé à 210 g de carbone par m 2 et par an. Les marais maritimes ont été gérés par les humains pendant des siècles, pour le pâturage, la fenaison, l'agriculture, le développement et les ports, les bassins d'évaporation pour la production de sel, l'aquaculture, la lutte contre les insectes, l'énergie marémotrice et la protection contre les inondations[20]. Les marais sont également sensibles à la pollution par le pétrole, les produits chimiques industriels et, le plus souvent, à l'eutrophisation. Les espèces introduites, l'élévation du niveau de la mer, les barrages et la diminution de la sédimentation sont des changements à long terme qui affectent l'habitat des marais et, à leur tour, peuvent affecter leur potentiel de séquestration du carbone[21].
Algues
Les macroalgues et les microalgues sont à l'étude comme moyens possibles de séquestration du carbone[22]. Parce que les algues n'ont pas la lignine complexe des plantes terrestres, le carbone des algues est libéré dans l'atmosphère plus rapidement que le carbone capturé sur terre[23] - [24]. En raison de leur teneur élevée en lipides, les microalgues ont été proposées comme réservoir de stockage à court terme de carbone pour être utilisées comme matière première pour la production de divers biocarburants et en particulier pour la production de biodiesel et de biométhane[25]. Les macroalgues, en revanche, n'ont pas une teneur élevée en lipides et ont un potentiel limité en tant que matière première de biodiesel, bien qu'elles puissent être utilisées comme matière première pour la génération d'autres biocarburants. Les macroalgues ont également été étudiées comme matière première pour la production de biochar car le biochar produit à partir de macroalgues est plus concentré en nutriments pour l'agriculture que le biochar produit à partir de sources terrestres[26].Une autre approche novatrice de la capture du carbone à partir des algues est le système intégré de capture et de production d'algues à base de bicarbonate (BICCAPS) issu d'une collaboration entre la Washington State University aux États-Unis et la Dalian Ocean University en Chine. De nombreuses espèces de cyanobactéries, de microalgues et de macroalgues peuvent utiliser le carbonate comme source de carbone pour la photosynthèse. Dans le BICCAPS, les microalgues alcaliphiles utilisent du carbone capturé à partir de gaz de combustion sous forme de bicarbonate[27] - [28]. En Corée du Sud, les macroalgues ont été utilisées dans le cadre d'un programme d'atténuation du réchauffement climatique. Le pays a mis en place une ceinture côtière d'élimination du CO 2 (CCRB) composée d'écosystèmes naturels et artificiels. L'objectif est de capturer le carbone à l'aide de vastes zones de forêt de kelp[29].
Restauration de l'écosystème
La restauration des mangroves, des herbiers marins, des marais et des forêts de varech est mise en œuvre dans de nombreux pays. Les herbiers marins restaurés séquestrent le carbone dans les sédiments au bout de quatre ans lorsque le pré atteint une densité de pousses suffisante pour provoquer le dépôt de sédiments[6]. Les plantations de mangroves en Chine montrent des taux de sédimentation plus élevés que les terres stériles et plus faibles que les forêts de mangroves établies. Ce schéma de vitesse de sédimentation semble être dû au jeune âge de la plantation et à une densité de végétation plus faible [30].
Stœchiométrie nutritive des herbiers
Les principaux nutriments qui déterminent la croissance des herbiers sont le carbone (C), l'azote (N), le phosphore (P) et la lumière nécessaire à la photosynthèse. L'azote et le phosphore peuvent être obtenus à partir de l'eau interstitielle des sédiments ou de la colonne d'eau, et les herbes marines peuvent absorber l'azote sous forme d'ammonium (NH 4+ ) et de nitrate (NO 3− )[23].
Un certain nombre d'études du monde entier ont révélé qu'il existe une large gamme de concentrations de C, N et P dans les herbiers, en fonction de leurs espèces et des facteurs environnementaux. Par exemple, les plantes collectées dans des environnements riches en nutriments avaient des rapports C/ N et C/P inférieurs à ceux des plantes collectées dans des environnements pauvres en nutriments. La stœchiométrie des herbiers marins ne suit pas le rapport de Redfield couramment utilisé comme indicateur de la disponibilité des nutriments pour la croissance du phytoplancton. En fait, un certain nombre d'études du monde entier ont montré que les rapports C/ N et C/P dans les herbiers marins peuvent varier considérablement en fonction de leurs espèces, de la disponibilité des nutriments ou d'autres facteurs environnementaux. Selon les conditions environnementales, les herbiers peuvent être limités en P ou en N[31].
Une première étude de la stœchiométrie des herbiers marins a suggéré que le ratio de Redfield entre N et P pour les herbiers marins était d'environ 30[24]. Cependant, les concentrations de N et de P ne sont pas strictement corrélées, ce qui suggère que les herbiers marins peuvent adapter leur absorption de nutriments en fonction de ce qui est disponible dans l'environnement. Par exemple, les herbiers marins proches des prairies fertilisées avec des excréments d'oiseaux ont montré une proportion plus élevée de phosphate que près des prairies non fertilisées. Alternativement, les herbiers marins dans des environnements avec des taux de charge plus élevés et une diagenèse de la matière organique fournissent plus de P, conduisant à une limitation en N. La disponibilité du P dans T. testudinum est le nutriment limitant. La distribution des nutriments dans T. testudinum varie entre 29,4-43,3% C, 0,88-3,96% N et 0,048-0,243% P. Cela équivaut à un rapport moyen de 24,6 C/ N, 937,4 C/ P et 40,2 N/P. Ces informations peuvent également être utilisées pour caractériser la disponibilité en nutriments d'une baie ou d'un autre plan d'eau (qui est difficile à mesurer directement) en échantillonnant les herbiers marins qui y vivent[21].
La disponibilité de la lumière est un autre facteur qui peut affecter la stœchiométrie des nutriments des herbiers marins. La limitation des nutriments ne peut se produire que lorsque l'énergie photosynthétique entraîne une croissance plus rapide des herbes que l'afflux de nouveaux nutriments. Par exemple, les environnements à faible luminosité ont tendance à avoir un rapport C/ N inférieur[21]. Alternativement, les environnements riches en N peuvent avoir un effet négatif indirect sur la croissance des herbiers marins en favorisant la croissance des algues qui réduisent la quantité totale de lumière disponible[17].
La variabilité des nutriments dans les herbiers marins peut avoir des implications potentielles pour la gestion des eaux usées dans les environnements côtiers. Des quantités élevées de rejets anthropiques d'azote pourraient provoquer l'eutrophisation dans des environnements auparavant limités en N, conduisant à des conditions hypoxiques dans les herbiers marins et affectant la capacité de charge de cet écosystème[21].
Une étude des dépôts annuels de C, N et P des herbiers marins de P. oceanica dans le nord-est de l'Espagne a révélé que la prairie séquestrait 198 g C par m2 par an, 13,4 g N par m2 par an et 2,01 g P par m2 par an dans les sédiments. La reminéralisation ultérieure du carbone des sédiments due à la respiration a libéré environ 8% du carbone séquestré, soit 15,6 g C par m2 par an[28].
Distribution et déclin des écosystèmes de carbone bleu
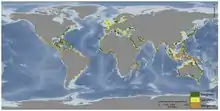
Les herbiers marins, les mangroves et les marais sont des habitats côtiers végétalisés qui couvrent environ 49 millions d'hectares dans le monde[32]. Les écosystèmes d'herbes marines se situent entre les régions polaires et les régions tropicales, les mangroves se situent dans les écosystèmes tropicaux et subtropicaux et les marais se situent principalement dans les régions tempérées comme sur la côte est des États-Unis. À mesure que les habitats qui séquestrent le carbone sont modifiés et disparaissent, cette quantité stockée de C est libérée dans l'atmosphère. Les impacts sur ces habitats à l'échelle mondiale libèrent directement et indirectement le carbone précédemment stocké, qui avait été séquestré dans les sédiments de ces habitats. Des déclins d'habitats côtiers végétalisés sont observés dans le monde entier; les exemples observés dans les mangroves sont dus au défrichement des étangs à crevettes comme c'est le cas en Indonésie, tandis que dans les herbiers marins il y a des causes naturelles dues aux agents pathogènes qui peuvent être exacerbées par des effets anthropiques. Il est difficile de quantifier cette disparition, mais les chercheurs estiment que si les écosystèmes de carbone bleu continuent de décliner, 30 à 40% des marais et des herbiers marins et la totalité des mangroves pourraient disparaître au siècle prochain.
Le déclin des herbiers marins est dû à un certain nombre de facteurs, notamment la sécheresse, la qualité de l'eau, les pratiques agricoles, les espèces envahissantes, les agents pathogènes, la pêche et le réchauffement climatique[33]. Il reste plus de 35% de l'habitat mondial des mangroves et leur perte est due aux barrages, au défrichement pour l'aquaculture, au développement,à la surpêche et au réchauffement climatique, selon le WWF[34]. Près de 16% des mangroves évaluées par l'UICN figurent sur la liste rouge de l'UICN et 1 mangrove sur 6 est menacée d'extinction[35]. Les barrages diminuent la quantité d'eau douce atteignant les mangroves. La destruction des récifs coralliens qui normalement ralentissent l'énergie des vagues à un niveau auquel les mangroves sont plus tolérantes joue aussi un rôle. Les marais salés ne sont peut-être pas très étendus, mais ils ont un taux d'enfouissement de C estimé à 87,2 ± 9,6 Tg C par an, ce qui est plus de 50 fois supérieur à celui des forêts tropicales humides, 53 ± 9,6 Tg C par an[36]. Depuis les années 1800, les marais salés dont l'importance n'était pas comprise ont décliné de 25% ce qui a entraîné une diminution de ce puits de C, la libération de C dans l'atmosphère et une accélération des processus d'érosion due au manque de biomasse végétale.
Les raisons du déclin des mangroves, des herbiers marins et des marais sont les changements d'utilisation des terres, les effets du climat et de la sécheresse, les barrages construits dans le bassin versant, l'aquaculture et l'agriculture, l'aménagement du territoire et l'élévation du niveau de la mer. Cela peut entraîner une diminution importante de l'habitat disponible et donc une augmentation du C rejeté par les sédiments. À mesure que les effets anthropiques et le réchauffement climatique s'intensifieront, l'efficacité des puits de carbone bleu diminuera et les émissions de CO 2 augmenteront. Les données sur les taux de rejet de CO 2 dans l'atmosphère ne sont pas fiables actuellement; cependant, des recherches sont en cours pour recueillir de meilleures informations afin d'analyser les tendances. La perte de biomasse souterraine (racines et rhizomes) pourrait transformer ces habitats en sources plutôt qu'en puits de carbone[36].
Sédimentation et enfouissement du carbone bleu
Le carbone organique n'est séquestré dans le système océanique que s'il atteint le fond marin et est recouvert d'une couche de sédiments. La matière organique qui n'est pas enfouie par une couche de sédiments suffisamment profonde est sujette à une remise en suspension par les courants océaniques changeants, à la bioturbation par des organismes qui vivent dans la couche supérieure des sédiments marins et à la décomposition par des bactéries hétérotrophes. La séquestration du carbone n'a lieu que si les taux d'enfouissement par les sédiments sont supérieurs aux taux à long terme d'érosion, de bioturbation et de décomposition[17] - [37].
Variabilité spatiale de la sédimentation
La sédimentation est la vitesse à laquelle les particules flottantes ou en suspension s'enfoncent et s'accumulent au fond de l'océan. Plus le courant est rapide, plus il peut capter de sédiments. À mesure que les courants chargés de sédiments ralentissent, les particules tombent en suspension et se déposent sur le fond marin. En d'autres termes, un courant rapide peut emmener beaucoup de grains lourds, alors qu'un courant lent ne peut emmener que de minuscules morceaux ce qui accentue la variabilité de la quantité de sédiments en suspension et le taux de dépôt[37].
Haute mer
La zone pélagique a des taux de sédimentation très faibles car la plupart des sédiments y arrivant sont transportés par le vent, ce qui ne représente qu'une petite fraction de l'apport total de sédiments aux océans. De plus, il y a beaucoup moins de vie végétale et animale vivant en pleine mer. Par conséquent, les taux d'enfouissement du carbone sont faibles[38].
Marges côtières
Les marges côtières ont des taux de sédimentation élevés en raison de l'apport de sédiments par les fleuves, qui représentent la grande majorité de l'apport de sédiments dans l'océan. Dans la plupart des cas, les sédiments se déposent près de l'embouchure ou sont transportés vers le littoral par les vagues. À certains endroits, les sédiments tombent dans les canyons sous-marins et sont transportés hors du plateau, si le canyon est suffisamment grand ou le plateau est étroit. Les marges côtières contiennent également des espèces marines diverses et abondantes, en particulier dans les endroits qui subissent des remontées d'eau périodiques. Une vie marine plus abondante combinée à des taux de sédimentation plus élevés sur les marges côtières crée des points chauds pour l'enfouissement du carbone[17] - [39].
Canyons sous-marins
Les courants transportent les sédiments sur le plateau dans la direction côtière et traversent les canyons marins perpendiculairement. Au dessus de ces eaux beaucoup plus profondes, le courant ralentit et dépose des sédiments. Les taux d'enfouissement du carbone dans le canyon de Nazaré près du Portugal sont 30 fois supérieurs à la pente continentale adjacente. Ce canyon représente à lui seul environ 0,03% de l'enfouissement mondial de carbone organique terrestre dans les sédiments marins alors qu'il ne représente que 0,0001% de la superficie du plancher océanique du monde[38].
Changements opérés par l'homme dans les systèmes sédimentaires mondiaux
Les humains modifient les cycles sédimentaires à grande échelle depuis des milliers d'années grâce à un certain nombre de mécanismes.
Agriculture / défrichage
Le premier changement majeur du cycle sédimentaire mondial s'est produit lorsque les humains ont commencé à défricher les terres pour faire pousser des cultures. Dans un écosystème naturel, les racines des plantes retiennent les sédiments en place lorsqu'il pleut. Les arbres et les arbustes réduisent la quantité de pluie qui affecte les sols et crée des obstacles autour desquels les cours d'eau forestiers doivent circuler. Lorsque toute la végétation disparait, les précipitations ont un impact direct et lessivent les sols. Pour cette raison, le défrichement entraîne une augmentation des taux d'érosion par rapport à un système naturel.
Barrages
Les premiers barrages remontent à 3000 avant JC et ont été construits pour contrôler les crues et pour irriguer les champs. Lorsque la rivière chargée de sédiments atteint le réservoir d'un barrage, l'eau ralentit à mesure qu'elle s'accumule. Les sédiments sont déposés avant que l'eau ne traverse le barrage. L'utilisation de barrages pour contrôler les inondations réduit la capacité des canaux en aval à produire des sédiments. Étant donné que la grande majorité de la sédimentation se produit pendant les inondations, la fréquence et l'intensité réduites des inondations peuvent modifier radicalement les dépôts de sédiments. Pendant des milliers d'années, il y avait trop peu de barrages pour avoir un impact significatif sur les cycles sédimentaires mondiaux, à l'exception des impacts locaux sur quelques deltas fluviaux tels que le Nil. Cependant, la popularisation de l'énergie hydroélectrique au siècle dernier a provoqué un énorme boom dans la construction de barrages et actuellement, seulement un tiers des plus grands fleuves du monde coulent sans entrave vers l'océan[40].
Canalisation
Dans un système naturel, les rives d'une rivière serpentent lorsque différents canaux s'érodent, s'accumulent, s'ouvrent ou se ferment. Les inondations saisonnières submergent régulièrement les berges des rivières et déposent des nutriments dans les plaines inondables adjacentes. Ces services sont essentiels aux écosystèmes naturels, mais peuvent perturber les infrastructures humaines à proximité des rivières. De ce fait, les rivières dans les zones peuplées sont souvent canalisées, ce qui signifie que leurs berges et parfois leurs lits sont fixés avec des roches ou du béton, ce qui empêche l'érosion et empêche le déplacement du cours d'eau. Cela inhibe également la sédimentation.
Impacts
Actuellement, l'effet net de l'homme sur le cycle sédimentaire mondial est une réduction drastique de la quantité de sédiments déversés dans l'océan. Ainsi les deltas et les plages rétrécissent et les marais salés disparaissent. La capacité des marges côtières à enterrer le carbone bleu est amoindrie. La perte de séquestration du carbone dans les sédiments marins côtiers pourrait accélérer du réchauffement climatique mondial[41].
Autres facteurs influençant les taux d'enfouissement du carbone bleu
Densité de végétation
La densité de la végétation dans les mangroves, les herbiers marins et les marais est un facteur important dans les taux d'enfouissement du carbone. Elle doit être suffisante pour modifier suffisamment les débits d'eau et réduire l'érosion et augmenter le dépôt de sédiments[42].
Charge nutritive
Des augmentations de la capture et de la séquestration du carbone ont été observées dans les écosystèmes de mangroves et d'herbes marines qui ont été soumis à des charges nutritives élevées, intentionnellement ou en raison de sous-produits d'activités humaines[22]. La fertilisation intentionnelle a été utilisée dans la restauration des herbiers marins en installant des perchoirs pour oiseaux marins dans le pré voisin afin que les déjections d'oiseaux jouent le rôle d'engrais. La fertilisation permet aux variétés d'herbes marines à croissance rapide de croître. La composition en espèces de ces prairies est nettement différente de celle des prairies sous-marines d'origine, bien qu'une fois la prairie rétablie et la fertilisation terminée, les prairies reprennent une composition en espèces plus proche d'une prairie non perturbée[43]. Les recherches effectuées sur les sols de mangroves de la mer Rouge ont montré que l'augmentation des charges de nutriments dans ces sols n'augmente pas la minéralisation du carbone et la libération subséquente de CO 2[44]. Cet effet neutre de la fertilisation ne s'est pas révélé vrai dans tous les types de forêts de mangroves. Les taux de capture de carbone ont également augmenté dans ces forêts en raison de l'augmentation des taux de croissance des mangroves. Dans les forêts où la respiration augmente, la croissance des mangroves a également augmenté jusqu'à six fois le taux normal [24].
Approches techniques du carbone bleu
Une étude du département américain de l'Énergie de 2001 a proposé de reproduire un processus naturel de séquestration du carbone dans l'océan en combinant de l'eau riche en gaz CO 2 avec du carbonate [CO
3 ] pour produire un lisier de bicarbonate [HCO
3 ]. En pratique, le processus d'ingénierie pourrait impliquer l'hydratation du CO 2 des gaz de combustion de la centrale électrique et son passage à travers un lit poreux de calcaire pour «fixer» le carbone dans une solution de bicarbonate saturée. Cette solution pourrait ensuite être déposée en mer pour sombrer dans l'océan profond. Le coût de ce processus, de la capture à l'enfouissement de l'océan, était estimé entre 90 $ et 180 $ par tonne de CO 2 selon la distance de transport du calcaire, de l'eau de mer et de la solution de bicarbonate résultante.
Les avantages escomptés de la production de bicarbonate par rapport au CO 2 dissous dans l'eau de mer seraient une moindre acidité des océans et un enfouissement prolongé avant que le carbone capturé ne soit relâché dans l'atmosphère[45].
Références
- Nellemann, Christian et al. (2009): Blue Carbon. The Role of Healthy Oceans in Binding Carbon. A Rapid Response Assessment. Arendal, Norway: UNEP/GRID-Arendal
- (en) Negative Emissions Technologies and Reliable Sequestration: A Research Agenda, Washington, D.C., National Academies of Sciences, Engineering, and Medicine, , 45 p. (ISBN 978-0-309-48452-7, DOI 10.17226/25259, lire en ligne)
- Nelleman, « Blue carbon: the role of healthy oceans in binding carbon. » [archive du ]
- McLeod, « A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2 »
- Duarte, « Assessing the capacity of seagrass meadows for carbon burial: current limitations and future strategies. », Ocean Coastal Management,
- Greiner, McGlathery, Gunnell et McKee, « Seagrass restoration enhances "blue carbon" sequestration in coastal waters », PLOS ONE, vol. 8, no 8, , e72469 (PMID 23967303, PMCID 3743776, DOI 10.1371/journal.pone.0072469)
- Gattuso, « Light availability in the coastal ocean: impact on the distribution of benthic photosynthetic organisms and their contribution to primary production », Biogeosciences, vol. 3, no 4, , p. 489–513 (DOI 10.5194/bg-3-489-2006)
- Fourqurean, « Seagrass ecosystems as a globally significant carbon stock », Nature Geoscience, vol. 5, no 7, , p. 505–509 (DOI 10.1038/ngeo1477)
- Waycott, « Accelerating loss of seagrasses across the globe threatens coastal ecosystems », Proceedings of the National Academy of Sciences of the USA, vol. 106, no 30, , p. 12377–12381 (PMID 19587236, PMCID 2707273, DOI 10.1073/pnas.0905620106)
- Bouillon, « Mangrove production and carbon sinks: a revision of global budget estimates », Global Biogeochemical Cycles, vol. 22, no 2, , n/a (DOI 10.1029/2007gb003052, Bibcode 2008GBioC..22.2013B)
- Alongi, « Carbon sequestration in mangrove forests », Future Science, (lire en ligne)
- Alongi, « Present state and future of the world's mangrove forests », Environmental Conservation, (lire en ligne)
- (en) Hamilton et Friess, « Global carbon stocks and potential emissions due to mangrove deforestation from 2000 to 2012 », Nature Climate Change, vol. 8, no 3, , p. 240–244 (ISSN 1758-678X, DOI 10.1038/s41558-018-0090-4, arXiv 1611.00307)
- (en) Hamilton et Casey, « Creation of a high spatio-temporal resolution global database of continuous mangrove forest cover for the 21st century (CGMFC-21) », Global Ecology and Biogeography, vol. 25, no 6, , p. 729–738 (ISSN 1466-822X, DOI 10.1111/geb.12449, arXiv 1412.0722)
- Duarte, « Major rule of marine vegetation on the oceanic carbon cycle », Biogeosciences, vol. 2, , p. 1–8 (DOI 10.5194/bg-2-1-2005, lire en ligne)
- Spaulding, « World atlas of mangroves »,
- Chmura et Anisfield, « Global carbon sequestration in tidal, saline wetland soils », Global Biogeochemical Cycles, vol. 17, no 4, , n/a (DOI 10.1029/2002GB001917, Bibcode 2003GBioC..17.1111C)
- Chmura, « What do we need to assess the sustainability of the tidal salt marsh carbon sink? », Ocean and Coastal Management, vol. 83, , p. 25–31 (DOI 10.1016/j.ocecoaman.2011.09.006)
- Mudd, « Impact of dynamic feedbacks between sedimentation, sea-level rise, and biomass production on near-surface marsh stratigraphy and carbon accumulation », Estuarine, Coastal and Shelf Science, vol. 82, no 3, , p. 377–389 (DOI 10.1016/j.ecss.2009.01.028)
- Adam, « Saltmarshes in a time of change », Environmental Conservation,
- Fourqurean et Zieman, « Nutrient Content of the Seagrass Thalassia Testudinum Reveals Regional Patterns of Relative Availability of Nitrogen and Phosphorus in the Florida Keys USA », Biogeochemistry, vol. 61, no 3, , p. 229–45 (DOI 10.1023/A:1020293503405)
- Kumar, Banerjee et Das, « Carbon dioxide sequestration from industrial flue gas by Chlorella sorokiniana », Bioresource Technology, vol. 152, , p. 225–233 (DOI 10.1016/j.biortech.2013.10.098)
- Chung, Beardall, Mehta et Sahoo, « Using marine macroalgae for carbon sequestration: a critical appraisal », Journal of Applied Phycology, vol. 23, no 5, , p. 877–886 (DOI 10.1007/s10811-010-9604-9)
- Mcleod, Chmura, Bouillon et Salm, « A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2 », Frontiers in Ecology and the Environment, vol. 9, no 10, , p. 552–560 (DOI 10.1890/110004, lire en ligne)
- Kumar, Dasgupta, Nayak et Lindblad, « Development of suitable photobioreactors for CO 2 sequestration addressing global warming using green algae and cyanobacteria », Bioresource Technology, vol. 102, no 8, , p. 4945–4953 (PMID 21334885, DOI 10.1016/j.biortech.2011.01.054)
- Bird, Wurster, de Paula Silva et Bass, « Algal biochar–production and properties », Bioresource Technology, vol. 102, no 2, , p. 1886–1891 (DOI 10.1016/j.biortech.2010.07.106)
- Chi, Z., O’Fallon, J. V., & Chen, S. (2011). Bicarbonate produced from carbon capture for algae culture. Trends in biotechnology, 29(11), 537-541.
- Chi, Xie, Elloy et Zheng, « Bicarbonate-based integrated carbon capture and algae production system with alkalihalophilic cyanobacterium », Bioresource Technology, vol. 133, , p. 513–521 (DOI 10.1016/j.biortech.2013.01.150)
- Chung, I. K., Oak, J. H., Lee, J. A., Shin, J. A., Kim, J. G., & Park, K. S. (2013). Installing kelp forests/seaweed beds for mitigation and adaptation against global warming: Korean Project Overview. ICES Journal of Marine Science: Journal du Conseil, fss206.
- Zhang, Cheng-De, Hai et Jun, « Estimating change in sedimentary organic carbon content during mangrove restoration in southern China using carbon isotopic measurements », Pedosphere, vol. 22, no 1, , p. 58–66 (DOI 10.1016/s1002-0160(11)60191-4)
- Fourqurean, Zieman et Powell, « Phosphorus Limitation of Primary Production in Florida Bay: Evidence from C:N:P Ratios of the Dominant Seagrass Thalassia Testudinum », Limnology and Oceanography, vol. 37, no 1, , p. 162–71 (DOI 10.4319/lo.1992.37.1.0162)
- Pendleton, Donato, Murray et Crooks, « Estimating Global "Blue Carbon" Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems », PLoS ONE, vol. 7, no 9, , e43542 (PMID 22962585, PMCID 3433453, DOI 10.1371/journal.pone.0043542)
- (en) Orth, Carruthers, Dennison et Duarte, « A Global Crisis for Seagrass Ecosystems », BioScience, vol. 56, no 12, , p. 987–996 (ISSN 0006-3568, DOI 10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2, lire en ligne)
- http://wwf.panda.org/about_our_earth/blue_planet/coasts/mangroves/mangrove_threats/
- « IUCN - Mangrove forests in worldwide decline », www.iucn.org (consulté le )
- Macreadie, Hughes et Kimbro, « Loss of 'Blue Carbon' from Coastal Salt Marshes Following Habitat Disturbance », PLoS ONE, vol. 8, no 7, , e69244 (PMID 23861964, PMCID 3704532, DOI 10.1371/journal.pone.0069244)
- (en-US) H., « A terrestrial organic matter depocenter on a high-energy margin adjacent to a low-sediment-yield river : the Umpqua River margin, Oregon », ir.library.oregonstate.edu (consulté le )
- Masson, Huvenne, Stigter et Wolff, « Efficient burial of carbon in a submarine canyon », Geology, vol. 38, no 9, , p. 831–834 (DOI 10.1130/g30895.1)
- Nittrouer, C. A. (2007). Continental margin sedimentation: From sediment transport to sequence stratigraphy. Malden, MA: Blackwell Pub. for the International Association of Sedimentologists.
- Dandekar, P. (2012). Where Rivers Run Free. Retrieved February 24, 2016, from https://www.internationalrivers.org/resources/where-rivers-run-free-1670
- « World's large river deltas continue to degrade from human activity » [archive du ], News Center (consulté le )
- Hendriks, I. E., Sintes, T., Bouma, T. J., & Duarte, C. M. (2008). Experimental assessment and modeling evaluation of the effects of the seagrass Posidonia oceanica on flow and particle trapping.
- Herbert et Fourqurean, « Ecosystem structure and function still altered two decades after short-term fertilization of a seagrass meadow », Ecosystems, vol. 11, no 5, , p. 688–700 (DOI 10.1007/s10021-008-9151-2)
- Keuskamp, Schmitt, Laanbroek et Verhoeven, « Nutrient amendment does not increase mineralisation of sequestered carbon during incubation of a nitrogen limited mangrove soil », Soil Biology and Biochemistry, vol. 57, , p. 822–829 (DOI 10.1016/j.soilbio.2012.08.007)
- Rau, G., K. Caldeira, K.G. Knauss, B. Downs, and H. Sarv, 2001. Enhanced Carbonate Dissolution as a Means of Capturing and Sequestering Carbon Dioxide. First National Conference on Carbon Sequestration Washington D.C., May 14–17, 2001.