Théorie du coureur de fond
La théorie du coureur de fond propose que certains caractères évolutifs de l'être humain peuvent s'expliquer par une adaptation spécifique à la course de fond[1] - [2].
Une telle adaptation à la course d'endurance a pu jouer un rôle important dans la stratégie alimentaire des premiers représentants du genre Homo. Elle a pu leur permettre dans un premier temps de couvrir plus d'espace dans leur recherche de nourriture, puis leur aurait permis de pratiquer la chasse à l'épuisement.
Différence entre marche et course


La marche et la course reposent sur des principes biomécaniques différents. La marche fonctionne sur le modèle d'un « pendule inversé », où le centre de masse bascule d'un côté sur l'autre, pendant que les jambes tendues alternent à chaque pas une transformation entre énergie cinétique et énergie potentielle de gravitation[2].
De son côté, la course fonctionne sur le modèle d'un système masse-ressort, au cours duquel l'énergie cinétique est transférée vers des tendons et des ligaments où elle est accumulée sous forme d'énergie potentielle élastique, puis restituée. Quand la vitesse est suffisamment grande, ce système masse-ressort demande un moindre coût énergétique que celui du pendule inversé pour un déplacement donné[2]. Inversement, un tel système élastique ne présente pas d'avantage particulier pour la marche.
Bien que le système masse-ressort soit donc plus favorable à grande vitesse, il implique aussi de plus grandes forces de réaction au sol que ne le fait la marche[2], et est moins stable parce qu'il provoque des mouvements des membres et des oscillations verticales du tronc. À l'inverse, le centre de gravité du corps se déplace beaucoup moins lors de la marche, ce qui en fait une allure intrinsèquement plus stable.
Adaptations anatomiques et physiologiques
Avantages sélectifs pour la course
De nombreuses recherches ont porté sur la manière dont la bipédie a pu être acquise par la lignée des hominines, mais peu se sont penchées sur la manière dont une adaptation spécifique à la course a pu être acquise par le genre Homo, et en quoi ces adaptations ont pu influencer l'évolution humaine.
Le corps humain a acquis un certain nombre de traits adaptatifs permettant d'améliorer la stabilisation lors de la course, et d'une manière générale améliorant la performance du système masse-ressort. Les recherches ayant porté sur la course ont permis de montrer que certaines structures ou fonctions physiologiques ne procurent un avantage sélectif que pour la course, et ne sont pas utilisées lors de la marche. Ceci tend à montrer que la capacité à la course est une adaptation spécifique, et non une simple conséquence de la bipédie.
Ces adaptations, décrites ci-dessous, sont présentées dans la « théorie du coureur de fond » comme autant de preuves en faveur d'une adaptation sélective à la course de fond. Quelques-unes de ces adaptations apparaissent avec Homo habilis, mais elles n'atteignent leur quasi-complétude qu'avec Homo ergaster, apparu en Afrique il y a 2 millions d'années[3] - [4].
Cependant, même si dans l'ensemble ces changements anatomiques pointent vers une adaptation à la course, il convient de rester prudent dans le détail, telle ou telle adaptation ayant pu avoir une autre origine. En particulier, les adaptations présentes chez Homo sapiens mais absentes chez Homo neanderthalensis (taille des canaux semi-circulaires, taille du talon) auraient un lien incertain avec une adaptation à la course.
Adaptations squelettiques
La comparaison entre le squelette des Australopithèques et celui des Homo révèle de nombreuses différences structurelles qui peuvent influer significativement sur la course de fond.
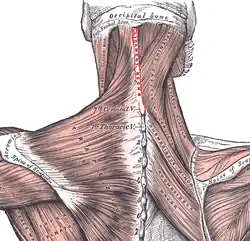
Parce que la tête n'est pas solidaire des épaules, elle doit être stabilisée pendant la course. Le ligament nuchal est un trait évolutif important permettant de stabiliser la tête. Il part du milieu de l'os occipital et se rattache au niveau du muscle trapèze à un processus épineux de la septième vertèbre cervicale. Ce ligament est très important en termes de paléoanthropologie, parce que si le ligament lui-même ne se fossilise pas, son point d'attache sur le crâne est marqué par une petite indentation et une crête osseuse, ce qui permet aux chercheurs de déterminer quelles espèces étaient pourvues d'un tel ligament. Comme ce ligament est absent chez l'Australopithèque et chez le Chimpanzé, on peut en conclure qu'il est spécifique au genre Homo[5].
Le squelette humain diffère de celui des autres hominidés en ce que l'arrière de la ceinture scapulaire est beaucoup moins fortement relié aux épaules et au haut du dos. Une liaison solide est un avantage pour l'escalade ou la progression par brachiation, mais inversement, elle devient une gêne entravant les mouvements du haut du corps nécessaires pour compenser le balourd des membres inférieurs et ainsi stabiliser le tronc et la tête pendant la course. Une telle stabilisation n'est pas nécessaire pour la marche.
- Longueur et masse des membres :
Par rapport à sa masse corporelle, Homo présente des membres inférieurs plus longs, ce qui permet de diminuer le cout énergétique de la course en augmentant le temps pendant lequel le membre est en contact avec le sol[2]. C'est en particulier le cas à partir d'Homo ergaster[6] D'autre part, la partie distale des membres est proportionnellement moins massive, ce qui diminue le cout énergétique de la course d'endurance, mais n'a guère d'effet sur la masse[2]. Enfin, la masse des membres supérieurs a fortement décru chez Homo par rapport à la masse corporelle d'ensemble, ce qui est un facteur important de stabilisation de ces membres pendant la course.
- Surface des articulations :
L'évolution du squelette des Homo leur a permis de subir les efforts et chocs importants produits par la course. L'accélération induite par les impacts d'une course de fond peut atteindre trois ou quatre fois le poids du corps, ce qui soumet l'ensemble de la structure squelettique à de fortes contraintes. Pour réduire ces contraintes, les squelettes Homo présentent des surfaces articulaires importantes qui répartissent ces forces et diminuent les pressions correspondantes, tout particulièrement vers le bas du corps[2]. Cette adaptation n'est pas présente sur le squelette des Australopithèques.
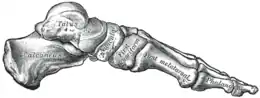
La voûte plantaire humaine a une fonction élastique qui permet d’emmagasiner et de restituer l'énergie pendant la course, mais n'est pas fonctionnelle pour la marche[2]. Les fossiles des Australopithèques ne présentent que des voûtes plantaires incomplètement formées, ce qui tend à montrer qu'elles étaient moins élastiques. De plus, pour que le rôle élastique de la voûte plantaire puisse être pleinement efficace, le pied ne doit pas jouer en rotation entre l'avant et l'arrière du pied. Ce blocage est provoqué par l'os du gros orteil et la structure compacte du squelette du pied, que l'on ne voit apparaître sous cette forme qu'à partir d'Homo habilis[2].

- Tubérosité postérieure du calcanéus et tendon d'Achille :
La longueur de la tubérosité postérieure du calcanéus (le bout du talon) est corrélée avec la longueur du tendon d'Achille. Quand le pied se replie, un bras de levier plus important au niveau du calcanéus impose au tendon d'Achille un déplacement plus grand, et donc permet d'emmagasiner plus d'énergie élastique, ce qui améliore l'économie générale de la course.
La paléoanthropologie montre que cette adaptation n'est pas présente chez Homo neanderthalensis, ce qui suggère qu'elle n'était pas nécessaire auparavant et a pu être une adaptation plus tardive à la course d'endurance[7].
- Raccourcissement des orteils :
Si on les compare aux autres hominidés, les orteils humains sont rectilignes, et très courts par rapport aux proportions générales du corps. Pendant la course, les orteils soutiennent 50 à 75 % du poids de l'homme. L'impulsion et le travail mécanique nécessaires pour une foulée augmentent avec la longueur des orteils, montrant qu'il est énergétiquement favorable d'avoir des orteils courts. Le coût à payer est une moindre performance pour grimper aux arbres, et pour utiliser les orteils en force ; mais ce coût paraît faible en regard du bénéfice, puisque les orteils d'Australopithecus afarensis sont déjà significativement plus courts que ceux des autres hominidés et restent encore 40 % plus longs que ceux des humains modernes, démontrant une tendance générale au raccourcissement des orteils au fur et à mesure que l'on s'éloigne de la vie arboricole. En théorie, cet écart de 40 % par rapport aux humains modernes induirait un moment d'impulsion 2,5 fois plus important chez Afarensis, qui imposerait deux fois plus de travail mécanique consacré à la stabilisation[8].
Stabilisation
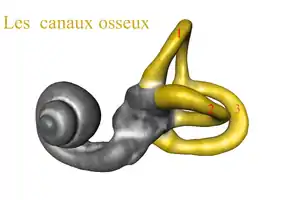
Le canal semi-circulaire est une formation de trois tubes interconnectés dans l'oreille interne, qui interviennent pour détecter les mouvements angulaires de la tête, et par conséquent jouent un rôle critique pour détecter les mouvements, maintenir l'équilibre et assurer la coordination des mouvements. Des études comparatives montrent que les animaux dont le canal semi-circulaire est plus grand ont plus de facilité pour détecter les mouvements de la tête et par conséquent, peuvent se déplacer avec une plus grande agilité même à grande vitesse.
Sur le plan évolutif, on constate que l'Homme de Néandertal a des canaux semi-circulaires très réduits, mais que ceux-ci sont beaucoup plus grands chez Homo sapiens. Il est possible que cette adaptation ait également été une réponse adaptative plus tardive à une plus grande sollicitation envers la course de fond[7].
- Réflexe vestibulo-oculaire :
Le système vestibulaire transmet de manière réflexe les accélérations angulaires de la tête aux muscles qui commandent le mouvement des yeux, ce qui permet à ceux-ci d'ajuster par réflexe leur mouvement pour stabiliser l'image formée sur la rétine. C'est une adaptation importante pour la course, parce qu'elle permet aux humains de bénéficier d'une meilleure vision malgré les mouvements saccadés induits par la course[5].
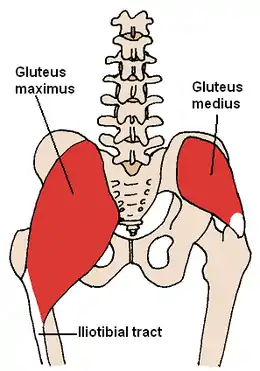
Le gluteus maximus du genre Homo est significativement plus grand que celui des Australopithèques. Cette taille est bien adaptée pour stabiliser le tronc pendant la course, mais des glutéaux de cette taille et de cette force ne sont pas nécessaires pour simplement pouvoir marcher.
- Épine iliaque :
Le genre Homo présente des aires d'attache musculaire plus larges au niveau du sacrum et de l'ilium, pour les muscles utilisés dans la stabilisation du tronc. Ces muscles réduisent le basculement du corps vers l'avant lors des chocs causés par les foulées de la course.
Performance énergétique
Outre des adaptations du squelette et de la stabilisation, des adaptations permettant une meilleure dissipation de la chaleur sont un point de passage obligé pour donner aux humains une capacité de coureur de fond[1]. La durée pendant laquelle un animal peut courir est déterminée par sa capacité à dissiper plus de chaleur que n'en produit l'effort de sa course, de manière à éviter une montée de la température corporelle qui peut être fatale.
La plupart des mammifères, y compris le genre humain, utilisent le refroidissement par évaporation pour réguler la température corporelle. Celui-ci peut prendre deux formes : la transpiration, ou le halètement. La plupart des grands mammifères utilisent la stratégie du halètement, alors que le genre Homo est l'un des rares (avec le cheval par exemple) à utiliser la transpiration. L'avantage du halètement est qu'il implique une moindre surface de peau, une moindre perte de sel, et la thermorégulation fonctionne localement en convection forcée, au lieu de s'appuyer sur une ventilation extérieure (vent, marche, ou éventail). Inversement, l'avantage de la transpiration est qu'elle porte sur toute la peau, donc une surface beaucoup plus grande, et qu'elle est indépendante de la ventilation forcée qu'est la respiration, ce qui en fait un mode de refroidissement beaucoup plus performant pour une activité intense comme celle de la course.
Chez l'homme, les glandes sudoripares sont sous un contrôle neuronal plus important que celui d'autres espèces, conduisant à une production de plus de sueur par unité de surface corporelle que pour n'importe quelle autre espèce. De plus, l'efficacité de la thermorégulation par évaporation de la sueur est augmentée par la réduction de la pilosité de la peau humaine[1]. En s'étant débarrassés de leur manteau de fourrure, les humains en train de courir peuvent dissiper avec une plus grande efficacité la chaleur générée par cet exercice.
Les hominines disposent de moyens de respiration libérés d'une contrainte. D'une manière générale, chez les mammifères, la respiration pendant la course est affectée par les contraintes induites par le squelette et la musculature. Le squelette et les muscles de la cage thoracique jouent leur rôle pour absorber les chocs de la course, mais surtout, ils sont contraints par les cycles de compression et de détente induits par l'allure de course elle-même. À cause de cette contrainte, les quadrupèdes doivent nécessairement coordonner un cycle de respiration et d'expiration au cycle de leur allure, et doivent donc nécessairement coordonner ces deux cycles[1]. Cette nécessaire coordination se traduit par une autre contrainte : pour une anatomie donnée, il n'y a qu'une seule vitesse de course qui soit énergétiquement optimale.
En ce qui concerne les « singes debout » que sont les hominines, la station bipède les libère de cette liaison physiologiquement nécessaire entre allure et respiration. Parce que leur cage thoracique n'est plus directement comprimée, ni même impliquée, dans les gestes nécessaires à la course, les hominines peuvent respirer indépendamment de leur allure[1]. Cette versatilité dans le rythme de respiration permet aux hominines de disposer d'un spectre beaucoup plus large de vitesses possibles dans la course.
- Stockage et utilisation de l'énergie :
Pendant une activité de longue durée, les animaux dépendent d'une combinaison de deux sources d'énergie : le glycogène, qui est conservé dans les muscles et le foie, et la graisse. Parce que le glycogène est plus facilement oxydé que la graisse et est disponible localement, c'est lui qui est consommé en premier. Mais quand l'effort doit se prolonger sur de longues durées, la demande énergétique impose de puiser dans les ressources de graisse. C'est le cas de tous les mammifères, mais les humains peuvent adapter leur alimentation pour mieux répondre à ces besoins énergétiques de longue durée.
Outre cette flexibilité alimentaire, les humains disposent d'une thyroïde et de glandes surrénales, qui leur permettent d'utiliser l'énergie des glucides et des acides gras d'une manière à la fois plus rapide et plus efficace. Ces organes relâchent des hormones diverses, dont notamment l'adrénaline, la noradrénaline, l'hormone corticotrope (ACTH) et la thyroxine. Une taille plus grande de ces glandes permet donc une plus grande production de ces hormones et en fin de compte, un meilleur métabolisme de ces réserves énergétiques[1].
L'un dans l'autre, cette adaptabilité alimentaire et l'utilisation plus efficiente des réserves énergétiques renforce cette capacité qu'ont les humains à ne pas être limités à une seule allure de course optimale comme le sont généralement les quadrupèdes. Pour la plupart des quadrupèdes, une course plus rapide impose une plus grande consommation d'oxygène et de réserves énergétiques. Avec leur bipédie et leur structure squelettique, les humains peuvent courir sur un grand éventail de vitesses et d'allures, tout en maintenant un taux de consommation d'énergie de l'ordre de 4,1 MJ pour 15 km. Ils améliorent ainsi grandement leur bilan énergétique[1].
Avantage sélectif de la course de fond
Recherche de nourriture

Toutes les adaptations mentionnées ci-dessus ont permis aux premiers représentants du genre Homo de rechercher leur nourriture avec une plus grande efficacité. Se déplacer en courant permet d'accéder à des carcasses distantes, ou toute autre source de nourriture, plus rapidement que d'autres charognards ou d'autres concurrents.
La stratégie du charognard peut être opportuniste ou stratégique. Les premiers humains pratiquaient certainement un charognage opportuniste. Un charognage peut être qualifié d'opportuniste quand « ils tombent sur une carcasse au cours de leur activité quotidienne de recherche de nourriture »[5].
Un charognage stratégique implique au contraire une recherche délibérée de carcasses. C'est pour ce type de charognage que la course de fond peut avoir été un avantage adaptatif de nos premiers ancêtres. Le charognage stratégique s'appuie sur des indices visibles à grande distance, comme des oiseaux charognards en train de tourner au-dessus d'une carcasse. Maîtriser la course de fond est alors un avantage, parce qu'elle permet aux humains d'atteindre plus rapidement la carcasse. La pression sélective aurait dû être très forte dans ce contexte, parce que les humains sont des animaux diurnes, alors que leurs principaux concurrents (hyènes, lions, etc.) ne le sont pas. Ils doivent donc atteindre rapidement les carcasses pour en tirer le plus possible avant la tombée de la nuit. En outre, une pression sélective supplémentaire résulte de la faiblesse naturelle des humains : parce qu'ils n'ont pas d'arme naturelle, ils n'ont pas la capacité de chasser d'une carcasse un groupe de gros carnivores qui décide d'en profiter. De ce fait, il est d'autant plus urgent qu'ils puissent atteindre la carcasse avant ces concurrents[5].
Chasse à l'épuisement
La chasse à l'épuisement est « une forme de chasse au cours de laquelle l'homme utilise la course d'endurance pendant la grande chaleur de midi pour amener sa proie dans un état d'hyperthermie et d'épuisement tel qu'elle peut alors être facilement rejointe et abattue »[1] - [5].
La chasse à l'épuisement se pratique vers le milieu du jour, sous le soleil plombant, au moment le plus chaud de la journée. Le chasseur se choisit une proie et la chasse à une vitesse intermédiaire entre son trot et son galop, ce qui pour un quadrupède est la plus inefficace des allures. Il continue cette poursuite des heures durant. Il peut perdre de vue sa proie, et dans ce cas, il lui faut suivre la piste et connaitre les mœurs de l'animal pour poursuivre la chasse. En fin de compte, la proie forcée est aux abois, en hyperthermie et incapable de fuir plus avant. Le chasseur, qui a une meilleure thermorégulation et une meilleure capacité à varier l'allure, peut alors l'approcher et la mettre à mort pendant qu'elle est sans ressource et ne peut plus contre-attaquer.
- La chasse à l'épuisement doit être menée pendant la chaleur du jour, pour que l'animal puisse être forcé à l'hyperthermie.
- Le chasseur doit être capable de suivre son gibier à la trace, parce qu'il va vraisemblablement le perdre de vue pendant qu'il le force.
- Une telle course demande de grandes ressources en eau, en sel, et en glycogène.
- Bien que le taux de succès soit élevé (de l'ordre de 50 %[5]), des chasses infructueuses sont extrêmement couteuses pour le chasseur. La chasse à l'épuisement suppose donc un système social dans lequel les individus partagent la nourriture, de manière que les chasseurs bredouilles puissent en emprunter aux autres quand nécessaire.
Au Paléolithique inférieur, les Homo commençaient à peine à utiliser des outils de pierre sous forme de galet aménagé, et les lances n'étaient tout au plus que des bâtons mal aiguisés. Faute d'arme de jet, ils ne pouvaient chasser qu'à très faible distance — au plus de six à dix mètres[9].
Étant donné la difficulté que représente la poursuite d'un animal en fuite, la capacité à suivre une piste a dû être un préalable à la chasse à l'épuisement.
Les restes de squelettes suggèrent que pendant le Paléolithique moyen, les Homo ont eu recours au « pistage systémique » pour chasser des antilopes dans une prairie plus ouverte. Ce « pistage systémique » se limite dans ce cas à suivre les traces de pas de l'animal et peut facilement être utilisé pour suivre des herbivores sur un terrain où leur passage laisse des traces.
Par la suite, durant le Paléolithique moyen et supérieur, le pistage aurait évolué vers un « pistage spéculatif » : lorsque les traces ne peuvent pas facilement être suivies, l'Homme anticipe la réaction de son gibier et interprète d'autres indices pour déterminer où il a le plus de chances de se diriger, et localiser ainsi à nouveau sa proie[10].
Cette méthode avancée permet de retrouver une proie dans une plus grande variété de terrains, rendant encore plus plausible l'hypothèse d'une utilisation de la course de fond pour une chasse à l'épuisement.
Critiques de la théorie
Au-delà des éléments de preuve en faveur d'une adaptation à la course de fond, l'idée que la chasse à l'épuisement et la recherche de nourriture aient réellement pu constituer un avantage écologique a été critiquée.
Contre la chasse à l'épuisement
La principale critique contre la chasse à l'épuisement est que ce que l'on sait du paléohabitat et de la paléoécologie des premiers Homo ne portait pas particulièrement à un tel comportement.
On considère généralement que les premiers Homo vivaient dans des savanes arborées[11], un environnement formé de prairies ouvertes et d'ilots de végétation plus dense, intermédiaire entre la savane proprement dite et la forêt.
On sait d'autre part, d'après les témoignages archéologiques, que les ongulés ont été la principale proie des premiers Homo. Étant donné leur grande vitesse de course, il leur aurait été facile de distancer des Homo primitifs.
La présence d'arbres dans cet environnement aurait réduit la visibilité et donc la capacité de suivre à vue le gibier, ce qui implique qu'une telle chasse ne peut se pratiquer que si l'homme dispose au préalable de la capacité à suivre des pistes. Pour Pickering et Bunn, cette capacité est complexe et demande un niveau cognitif élevé, que les premiers Homo n'auraient pas eu. Leur capacité aurait pu leur permettre de suivre une piste sanglante laissée par un animal blessé au préalable, mais ils n'avaient pas le niveau technologique requis pour fabriquer les armes de jet permettant de blesser de loin un herbivore[11].
De nos jours, les chasseurs n'ont pratiquement jamais recours à la chasse à l'épuisement, et quand ils le font, ils abandonnent le plus souvent quand la piste qu'ils suivent se perd dans la végétation[11].
Les rares groupes de chasseurs qui la pratiquent occasionnellement peuvent le faire parce qu'ils vivent dans un environnement extrêmement chaud et ouvert. Pour ces chasseurs, une journée entière est nécessaire pour récupérer après une chasse, ce qui montre à quel point la chasse à l'épuisement est couteuse en énergie, et ce qui explique qu'elle ne soit que rarement pratiquée[11].
Enfin, une des critiques de l'étude par Liebenberg de la chasse à l'épuisement moderne a été que la plupart des chasses ayant fait l'objet de l'étude n'étaient pas spontanées mais ont été initiées pour les besoins du tournage. De plus, peu ont été couronnées de succès ; et les chasses fructueuses l'ont été à cause de facteurs externes, notamment parce que les chasseurs ont pu s'arrêter et remplir leurs bouteilles d'eau[11].
Une réponse à ces critiques a été donnée par Lieberman[12].
Contre la recherche de nourriture
L'avantage invoqué pour une course d'endurance est la capacité d'arriver sur une ressource de nourriture avant les concurrents[11]. Cependant, les études de paléoanthropologie montrent que l'habitat de savane arborée présente en réalité peu de concurrence : à cause de la faible visibilité à longue distance, les carcasses ne sont pas facilement localisées par les carnivores.
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Endurance running hypothesis » (voir la liste des auteurs).
- (en) D.R. Carrier et al., « The Energetic Paradox of Human Running and Hominid Evolution », Current Anthropology, vol. 25, no 4, , p. 483–495 (DOI 10.1086/203165, JSTOR 2742907)
- (en) Dennis Bramble et Daniel Lieberman, « Endurance running and the evolution of Homo », Nature, vol. 432, , p. 345–52 (PMID 15549097, DOI 10.1038/nature03052).
- Nature2004_EnduranceRunningandtheEvolutionofHomo.pdf Endurance running and the evolution of Homo, Dennis M. Bramble & Daniel E. Lieberman, Nature, vol. 432, 18 novembre 2004.
- How running made us human : Endurance running let us evolve to look the way we do, University Of Utah, ScienceDaily, 24 novembre 2004.
- (en) Daniel Lieberman, Bramble, Dennis, Raichlen, David et Shea, John, « Brains, Brawn, and the Evolution of Human Endurance Running Capabilities », Contributions from the Third Stony Brook Human Evolution Symposium and Workshop, , p. 77–92
- Humans Were Born to Run, Fossil Study Suggests
- (en) D.A. Raichlen, Armstrong, H et Lieberman, D.E., « Calcaneus length determines running economy: Implications for endurance running performance in modern humans and Neandertals. », Journal of Human Evolution, vol. 60, , p. 299–308 (DOI 10.1016/j.jhevol.2010.11.002, lire en ligne)
- (en) Campbell Rolian, Lieberman, Daniel, Hamill, Joseph, Scott, John et Werbel, William, « Walking, running and the evolution of short toes in humans », The Journal of Experimental Biology, , p. 212, 713–721
- (en) S.E. Churchill, « Weapon technology, prey size selection and hunting methods in modern hunter gatherers: implications for hunting in the Paleolithic and Mesolithic », Archeological Papers of the American Anthropological Association, no 4, , p. 11–24.
- (en) Louis Liebenberg, « Persistence Hunting by Modern Hunter-Gatherers », Current Anthropology (en), vol. 47, no 6, , p. 1017–1026 (DOI 10.1086/508695, lire en ligne).
- (en) Travis Rayne Pickering et Bunn, Henry, « The endurance running hypothesis and hunting and scavenging in savanna-woodlands », Journal of Human Evolution, vol. 53, no 4, , p. 434–438 (DOI 10.1016/j.jhevol.2007.01.012)
- (en) « The evolution of endurance running and the tyranny of ethnography : A reply to Pickering and Bunn (2007) »
Voir aussi
Articles connexes
Liens externes
- Notice dans un dictionnaire ou une encyclopédie généraliste :
- The endurance running hypothesis, the idea that humans evolved as long-distance runners, may have legs thanks to a new study on toes - SEED Magazine