Interféron gamma
L'interféron gamma (ou IFNγ) est une cytokine soluble dimérisée qui est le seul membre de la classe des interférons de type II[1]. L'existence de cet interféron, qui au début de son histoire, était connu sous le nom d'"interféron immunitaire", a été décrite par E. F. Wheelock comme produit des leucocytes humains activés par la phytohémagglutinine, ainsi que par d'autres chercheurs comme un produit des lymphocytes activés par des antigènes[2]. Il a également été démontré qu'il était produit dans les lymphocytes humains[3] ou dans les lymphocytes péritonéaux de souris sensibilisés à la tuberculine[4], atteints de DPP ; les surnageants qui en résultent inhibent la croissance du virus de la stomatite vésiculaire. Ces rapports contenaient également l'observation de base qui sous-tend le test de libération d'interféron gamma, désormais largement utilisé pour tester la tuberculose. Chez l'homme, la protéine IFNγ est codée par le gène IFNG[5] - [6].
Fonction
L'IFNγ, ou interféron de type II, est une cytokine essentielle à l'immunité innée et adaptative contre les infections virales, certaines bactéries et certains agents pathogènes unicellulaires. L'IFNγ est un activateur important des macrophages et inducteur de l'expression des molécules du complexe majeur d'histocompatibilité (CMH) de classe II. L'expression anormale d'IFNγ est associée à un certain nombre de maladies auto-inflammatoires et auto-immunes. L'importance de l'IFNγ dans le système immunitaire découle en partie de sa capacité à inhiber directement la réplication virale, et surtout de ses effets immunostimulants et immunomodulateurs. L'IFNγ est produit principalement par les cellules tueuses naturelles (NK) et les lymphocytes T tueurs naturels (NKT) dans le cadre de la réponse immunitaire innée, et par les cellules T effectrices des lymphocytes T cytotoxiques (CTL) CD4 Th1 et CD8 une fois que l'immunité spécifique à l'antigène se développe[7] - [8] dans le cadre de la réponse immunitaire adaptative. L'IFNγ est également produit par des cellules lymphoïdes innées non cytotoxiques (ILC), une famille de cellules immunitaires découvertes pour la première fois au début des années 2010[9].
Structure

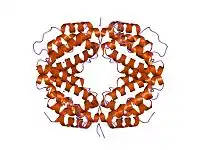
Le monomère d'IFNγ consiste en un cœur de six hélices alpha ainsi qu'une séquence étendue non pliée dans la région C-terminale (voir figure ci-contre à gauche)[10]. Les hélices α au cœur de la structure sont numérotées de 1 à 6.

Le dimère biologiquement actif est formé par l'emboîtement antiparallèle des deux monomères comme indiqué ci-contre à droite. Dans le modèle de dessin animé, un monomère est représenté en rouge, l'autre en bleu.
Liaison au récepteur

Les réponses cellulaires à l'IFNγ sont activées par son interaction avec un récepteur hétérodimérique composé du récepteur d'interféron gamma numéro 1 (IFNGR1) et du récepteur d'interféron gamma numéro 2 (IFNGR2). La liaison de l'IFNγ avec le récepteur active la voie JAK-STAT. L'IFNγ se lie également au glycosaminoglycane sulfate d'héparane (HS) à la surface des cellules. Toutefois, contrairement à de nombreuses autres protéines de liaison au sulfate d'héparane où la liaison favorise l'activité biologique, ici la liaison de l'IFNγ au HS inhibe son activité biologique[11].
Les modèles structurels présentés dans les figures 1-3 pour l'IFNγ[10] sont tous raccourcis à leur extrémité C-terminale par 17 acides aminés. La longueur totale de l'IFNγ est de 143 acides aminés, les modèles sont de 126 acides aminés. L'affinité pour le sulfate d'héparane ne réside que dans la séquence supprimée de 17 acides aminés[12]. Dans cette séquence de 17 acides aminés se trouvent deux groupes d'acides aminés de base appelés respectivement D1 et D2. Le sulfate d'héparane interagit avec ces deux groupes[13]. En l'absence de sulfate d'héparane, la présence de la séquence D1 augmente la vitesse à laquelle les complexes IFNγ-récepteur se forment[11]. Les interactions entre le groupe d'acides aminés D1 et le récepteur peuvent être la première étape de la formation du complexe. En se liant à D1, les HS peuvent entrer en compétition avec le récepteur et empêcher la formation de complexes récepteurs actifs.
La signification biologique de l'interaction entre les sulfates d'héparane et l'IFNγ n'est pas claire ; cependant, la liaison du groupe D1 au HS pourrait le protéger du clivage protéolytique[13].
Activité biologique
L'IFNγ est sécrété par les cellules T auxiliaires (plus précisément, les cellules Th1), les lymphocytes T cytotoxiques (cellules CTL), les macrophages, les cellules épithéliales des muqueuses et les cellules NK. L'IFNγ est le seul interféron de type II et il est sérologiquement distinct des interférons de type I ; il est labile en présence d'acides, tandis que les variantes de type I sont stables en présence d'acides.
L'IFNγ a des propriétés antivirales, immunorégulatrices et antitumorales[14]. Il modifie la transcription dans jusqu'à 30 gènes, produisant une variété de réponses physiologiques et cellulaires. Parmi celles-ci, on peut citer :
- une induction de l'activité des cellules NK ;
- une augmentation de la présentation de l'antigène (à la surface des cellules infectées pour qu'elles soient reconnues par les cellules de l'immunité) et de l'activité des lysosomes des macrophages ;
- une activation de l'enzyme oxyde nitrique synthase inductible (iNOS) ;
- une induction de la production d'IgG2a et d'IgG3 à partir de lymphocytes B plasmatiques activés ;
- une augmentation de l'expression des molécules du CMH de classe I et du CMH de classe II dans les cellules présentant l'antigène, plus précisément par l'induction de gènes de traitement de l'antigène, y compris des sous-unités de l'immunoprotéasome (MECL1, LMP2, LMP7), ainsi que du TAP et de l'ERAAP, en plus éventuellement de la régulation positive directe des chaînes lourdes du CMH et de la microglobuline β2 elle-même ;
- une facilitation de l'adhésion et de la liaison nécessaires à la migration des leucocytes ;
- une induction de l'expression de facteurs de défense intrinsèques — par exemple, en ce qui concerne les rétrovirus, les gènes pertinents comprennent TRIM5alpha, APOBEC et Tetherin, représentant directement les effets antiviraux ;
- une priorisation des macrophages alvéolaires contre les infections bactériennes secondaires[15] - [16].
L'IFNγ est la cytokine primaire qui définit les cellules Th1 : Les cellules Th1 sécrètent l'IFNγ, qui à son tour provoque la différenciation d'un plus grand nombre de cellules CD4+ indifférenciées (cellules Th0) en cellules Th1, représentant une boucle de rétroaction positive — tout en supprimant la différenciation des cellules Th2. (Les cytokines définissant les autres cellules comprennent l'IL-4 pour les cellules Th2 et l'IL-17 pour les cellules Th17.)
Les cellules NK et les cellules T cytotoxiques CD8+ produisent également l'IFNγ. L'IFNγ supprime la formation d'ostéoclastes en dégradant rapidement la protéine d'adaptation TRAF6 du RANK dans la voie de signalisation RANK-RANKL, qui en temps normal stimule la production de NF-κB.
Activité dans la formation de granulome
Un granulome résulte de la façon dont l'organisme traite une substance qu'il ne peut pas éliminer ou stériliser. Les causes infectieuses des granulomes (les infections sont généralement la cause la plus courante des granulomes) comprennent la tuberculose, la lèpre, l'histoplasmose, la cryptococcose, la coccidioïdomycose, la blastomycose et la toxoplasmose. Parmi les maladies granulomateuses non infectieuses, on peut citer la sarcoïdose, la maladie de Crohn, la bérylliose, l'artérite à cellules géantes, la granulomatose avec polyangéite, la granulomatose éosinophilique avec polyangéite, les nodules rhumatoïdes pulmonaires et l'aspiration de nourriture et d'autres matières particulaires dans les poumons. La pathophysiologie infectieuse des granulomes est principalement abordée dans cette section.
La principale association entre l'IFNγ et les granulomes est que l'IFNγ active les macrophages afin qu'ils deviennent plus puissants pour tuer les organismes intracellulaires. L'activation des macrophages par l'IFNγ à partir des cellules auxiliaires Th1 dans les infections mycobactériennes permet aux macrophages de surmonter l'inhibition de la maturation des phagolysosomes causée par les mycobactéries (pour rester en vie à l'intérieur des macrophages)[17]. Les premières étapes de la formation de granulomes induite par l'IFNγ sont l'activation des cellules auxiliaires Th1 par les macrophages libérant l'IL-1 et l'IL-12 en présence de pathogènes intracellulaires, et la présentation des antigènes de ces pathogènes. Ensuite, les cellules auxiliaires Th1 s'agrègent autour des macrophages et libèrent l'IFNγ, qui active les macrophages. Une activation supplémentaire des macrophages entraîne un cycle de destruction supplémentaire des bactéries intracellulaires et une présentation supplémentaire des antigènes aux cellules auxiliaires Th1 avec une libération supplémentaire de l'IFNγ. Enfin, les macrophages entourent les cellules auxiliaires Th1 et se transforment en cellules semblables à des fibroblastes, qui deviennent des barrières contre l'infection.
Activité pendant la grossesse
Les cellules tueuses naturelles (NK) utérines sécrètent des niveaux élevés de chimioattracteurs, comme l'IFNγ. L'IFNγ dilate et amincit les parois des artères spirales maternelles pour améliorer le flux sanguin vers le site d'implantation. Ce remodelage favorise le développement du placenta qui envahit l'utérus dans sa quête de nutriments. Les souris knockout pour l'IFNγ ne parviennent pas à initier la modification normale des artères déciduales induite par la grossesse. Ces modèles présentent des quantités anormalement faibles de cellules ou une nécrose de la decidua[18].
Production
L'interféron gamma humain recombinant, en tant que produit biopharmaceutique coûteux, a été exprimé dans différents systèmes d'expression, notamment dans des cellules procaryotes, protozoaires, fongiques (levures), végétales, d'insectes et de mammifères. L'interféron gamma humain est couramment exprimé et produit chez Escherichia coli, commercialisé sous le nom d'Actimmune. Cependant, le produit résultant du système d'expression procaryote n'est pas glycosylé et possède une courte demi-vie dans la circulation sanguine après injection ; le processus de purification du système d'expression bactérien est également très coûteux. D'autres systèmes d'expression comme Pichia pastoris n'ont pas donné de résultats satisfaisants en termes de rendement[19] - [20].
Usage thérapeutique
L'interféron-γ 1b est approuvé par la Food and Drug Administration américaine pour traiter la maladie granulomateuse chronique[21] et l'ostéopétrose[22].
Il n'a pas été approuvé pour traiter la fibrose pulmonaire idiopathique (FPI). En 2002, le fabricant InterMune a publié un communiqué de presse indiquant que les données de la phase III ont démontré un bénéfice en termes de survie dans la FPI et ont réduit la mortalité de 70% chez les patients atteints d'une maladie légère à modérée. Le ministère américain de la justice a accusé le communiqué de contenir des déclarations fausses et trompeuses. Le directeur général d'InterMune, Scott Harkonen, a été accusé d'avoir manipulé les données du procès, a été reconnu coupable en 2009 de fraude électronique et a été condamné à des amendes et à des travaux d'intérêt général. Harkonen a fait appel de sa condamnation devant la Cour d'appel américaine pour le neuvième circuit, et a perdu[23].
La molécule est étudiée à l'hôpital des enfants de Philadelphie pour le traitement de l'ataxie de Friedreich[24].
Bien qu'il ne soit pas officiellement approuvé, l'interféron-γ s'est également révélé efficace dans le traitement des patients atteints de dermatite atopique modérée à sévère[25] - [26] - [27].
Utilisation potentielle en immunothérapie
L'interféron gamma n'est pas encore approuvé pour le traitement dans le cadre d'une immunothérapie contre le cancer. Toutefois, une amélioration de la survie a été observée lorsque l'interféron gamma a été administré à des patients atteints de carcinomes de la vessie et de mélanomes. Le résultat le plus prometteur a été obtenu chez les patientes atteintes d'un carcinome ovarien de stade 2 ou 3. Au contraire, il a été souligné : « L'interféron-γ sécrété par les lymphocytes CD8-positifs régule la PD-L1 sur les cellules cancéreuses de l'ovaire et favorise la croissance de la tumeur »[28]. L'étude in vitro de l'IFNγ dans les cellules cancéreuses est plus étendue et les résultats indiquent une activité anti-proliférative de l'IFNγ conduisant à l'inhibition de la croissance ou à la mort cellulaire, généralement induite par apoptose mais parfois par autophagie. En outre, il a été signalé que la glycosylation de l'IFNγ humain recombinant, exprimé dans HEK293, chez les mammifères, améliore son efficacité thérapeutique par rapport à la forme non glycosylée qui est exprimée chez E. coli[29].
Interactions
On a montré que l'IFNγ interagissait avec le récepteur à l'interféron gamma numéro 1[10] - [30].
Maladies
On a montré que l'IFNγ a un rôle crucial dans la réponse immunitaire contre certains pathogènes intracellulaires, tels que celui qui cause la maladie de Chagas[31]. Il a également été identifié comme ayant un rôle dans la dermatite séborrhéique[32].
Régulation
Il est prouvé que l'expression de l'IFNγ est régulée par un élément pseudo-codé dans son UTR 5'[33]. Il est également prouvé que l'IFNγ est régulé directement ou indirectement par les microARN miR-29[34]. En outre, il est prouvé que l'expression de l'IFNγ est régulée par la GAPDH dans les lymphocytes T. Cette interaction a lieu dans l'UTR 3', où la liaison de la GAPDH empêche la traduction de la séquence de l'ARNm[35].
Voir aussi
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Interferon gamma » (voir la liste des auteurs).
Références
- P. W. Gray et D. V. Goeddel, « Structure of the human immune interferon gene », Nature, vol. 298, no 5877, , p. 859–863 (ISSN 0028-0836, PMID 6180322, DOI 10.1038/298859a0, lire en ligne, consulté le )
- E. F. Wheelock, « Interferon-Like Virus-Inhibitor Induced in Human Leukocytes by Phytohemagglutinin », Science (New York, N.Y.), vol. 149, no 3681, , p. 310–311 (ISSN 0036-8075, PMID 17838106, DOI 10.1126/science.149.3681.310, lire en ligne, consulté le )
- J. A. Green, S. R. Cooperband et S. Kibrick, « Immune specific induction of interferon production in cultures of human blood lymphocytes », Science (New York, N.Y.), vol. 164, no 3886, , p. 1415–1417 (ISSN 0036-8075, PMID 5783715, DOI 10.1126/science.164.3886.1415, lire en ligne, consulté le )
- L. M. Milstone et B. H. Waksman, « Release of virus inhibitor from tuberculin-sensitized peritoneal cells stimulated by antigen », Journal of Immunology (Baltimore, Md.: 1950), vol. 105, no 5, , p. 1068–1071 (ISSN 0022-1767, PMID 4321289, lire en ligne, consulté le )
- S. L. Naylor, A. Y. Sakaguchi, T. B. Shows et M. L. Law, « Human immune interferon gene is located on chromosome 12 », The Journal of Experimental Medicine, vol. 157, no 3, , p. 1020–1027 (ISSN 0022-1007, PMID 6403645, PMCID 2186972, DOI 10.1084/jem.157.3.1020, lire en ligne, consulté le )
- « IFNGR2 interferon gamma receptor 2 [Homo sapiens (human)] - Gene - NCBI », sur www.ncbi.nlm.nih.gov (consulté le )
- « IFNG interferon gamma [Homo sapiens (human)] - Gene - NCBI », sur www.ncbi.nlm.nih.gov (consulté le )
- Jamie R. Schoenborn et Christopher B. Wilson, « Regulation of interferon-gamma during innate and adaptive immune responses », Advances in Immunology, vol. 96, , p. 41–101 (ISSN 0065-2776, PMID 17981204, DOI 10.1016/S0065-2776(07)96002-2, lire en ligne, consulté le )
- David Artis et Hergen Spits, « The biology of innate lymphoid cells », Nature, vol. 517, no 7534, , p. 293–301 (ISSN 1476-4687, PMID 25592534, DOI 10.1038/nature14189, lire en ligne, consulté le )
- D. J. Thiel, M. H. le Du, R. L. Walter et A. D'Arcy, « Observation of an unexpected third receptor molecule in the crystal structure of human interferon-gamma receptor complex », Structure (London, England: 1993), vol. 8, no 9, , p. 927–936 (ISSN 0969-2126, PMID 10986460, DOI 10.1016/s0969-2126(00)00184-2, lire en ligne, consulté le )
- R. Sadir, E. Forest et H. Lortat-Jacob, « The heparan sulfate binding sequence of interferon-gamma increased the on rate of the interferon-gamma-interferon-gamma receptor complex formation », The Journal of Biological Chemistry, vol. 273, no 18, , p. 10919–10925 (ISSN 0021-9258, PMID 9556569, DOI 10.1074/jbc.273.18.10919, lire en ligne, consulté le )
- Cécile Vanhaverbeke, Jean-Pierre Simorre, Rabia Sadir et Pierre Gans, « NMR characterization of the interaction between the C-terminal domain of interferon-gamma and heparin-derived oligosaccharides », The Biochemical Journal, vol. 384, no Pt 1, , p. 93–99 (ISSN 1470-8728, PMID 15270718, PMCID 1134092, DOI 10.1042/BJ20040757, lire en ligne, consulté le )
- H. Lortat-Jacob et J. A. Grimaud, « Interferon-gamma binds to heparan sulfate by a cluster of amino acids located in the C-terminal part of the molecule », FEBS letters, vol. 280, no 1, , p. 152–154 (ISSN 0014-5793, PMID 1901275, DOI 10.1016/0014-5793(91)80225-r, lire en ligne, consulté le )
- Kate Schroder, Paul J. Hertzog, Timothy Ravasi et David A. Hume, « Interferon-gamma: an overview of signals, mechanisms and functions », Journal of Leukocyte Biology, vol. 75, no 2, , p. 163–189 (ISSN 0741-5400, PMID 14525967, DOI 10.1189/jlb.0603252, lire en ligne, consulté le )
- Friedrich Felix Hoyer, Kamila Naxerova, Maximilian J. Schloss et Maarten Hulsmans, « Tissue-Specific Macrophage Responses to Remote Injury Impact the Outcome of Subsequent Local Immune Challenge », Immunity, vol. 51, no 5, 11 19, 2019, p. 899–914.e7 (ISSN 1097-4180, PMID 31732166, PMCID 6892583, DOI 10.1016/j.immuni.2019.10.010, lire en ligne, consulté le )
- Yushi Yao, Mangalakumari Jeyanathan, Siamak Haddadi et Nicole G. Barra, « Induction of Autonomous Memory Alveolar Macrophages Requires T Cell Help and Is Critical to Trained Immunity », Cell, vol. 175, no 6, 11 29, 2018, p. 1634–1650.e17 (ISSN 1097-4172, PMID 30433869, DOI 10.1016/j.cell.2018.09.042, lire en ligne, consulté le )
- « A Controlled Trial of Interferon Gamma to Prevent Infection in Chronic Granulomatous Disease », New England Journal of Medicine, vol. 324, no 8, , p. 509–516 (ISSN 0028-4793, PMID 1846940, DOI 10.1056/NEJM199102213240801, lire en ligne, consulté le )
- A. A. Ashkar, J. P. Di Santo et B. A. Croy, « Interferon gamma contributes to initiation of uterine vascular modification, decidual integrity, and uterine natural killer cell maturation during normal murine pregnancy », The Journal of Experimental Medicine, vol. 192, no 2, , p. 259–270 (ISSN 0022-1007, PMID 10899912, PMCID 2193246, DOI 10.1084/jem.192.2.259, lire en ligne, consulté le )
- Ali Razaghi, Leigh Owens et Kirsten Heimann, « Review of the recombinant human interferon gamma as an immunotherapeutic: Impacts of production platforms and glycosylation », Journal of Biotechnology, vol. 240, , p. 48–60 (ISSN 1873-4863, PMID 27794496, DOI 10.1016/j.jbiotec.2016.10.022, lire en ligne, consulté le )
- Ali Razaghi, Emilyn Tan, Linda H. L. Lua et Leigh Owens, « Is Pichia pastoris a realistic platform for industrial production of recombinant human interferon gamma? », Biologicals: Journal of the International Association of Biological Standardization, vol. 45, , p. 52–60 (ISSN 1095-8320, PMID 27810255, DOI 10.1016/j.biologicals.2016.09.015, lire en ligne, consulté le )
- P. A. Todd et K. L. Goa, « Interferon gamma-1b. A review of its pharmacology and therapeutic potential in chronic granulomatous disease », Drugs, vol. 43, no 1, , p. 111–122 (ISSN 0012-6667, PMID 1372855, DOI 10.2165/00003495-199243010-00008, lire en ligne, consulté le )
- L. L. Key, W. L. Ries, R. M. Rodriguiz et H. C. Hatcher, « Recombinant human interferon gamma therapy for osteopetrosis », The Journal of Pediatrics, vol. 121, no 1, , p. 119–124 (ISSN 0022-3476, PMID 1320672, DOI 10.1016/s0022-3476(05)82557-0, lire en ligne, consulté le )
- Ed Silverman, « Drug Marketing. The line between scientific uncertainty and promotion of snake oil », BMJ (Clinical research ed.), vol. 347, , f5687 (ISSN 1756-1833, PMID 24055923, DOI 10.1136/bmj.f5687, lire en ligne, consulté le )
- L. Seyer, N. Greeley, D. Foerster et C. Strawser, « Open-label pilot study of interferon gamma-1b in Friedreich ataxia », Acta Neurologica Scandinavica, vol. 132, no 1, , p. 7–15 (ISSN 1600-0404, PMID 25335475, DOI 10.1111/ane.12337, lire en ligne, consulté le )
- Arash Akhavan et Donald Rudikoff, « Atopic dermatitis: systemic immunosuppressive therapy », Seminars in Cutaneous Medicine and Surgery, vol. 27, no 2, , p. 151–155 (ISSN 1085-5629, PMID 18620137, DOI 10.1016/j.sder.2008.04.004, lire en ligne, consulté le )
- L. C. Schneider, Z. Baz, C. Zarcone et D. Zurakowski, « Long-term therapy with recombinant interferon-gamma (rIFN-gamma) for atopic dermatitis », Annals of Allergy, Asthma & Immunology: Official Publication of the American College of Allergy, Asthma, & Immunology, vol. 80, no 3, , p. 263–268 (ISSN 1081-1206, PMID 9532976, DOI 10.1016/S1081-1206(10)62968-7, lire en ligne, consulté le )
- J. M. Hanifin, L. C. Schneider, D. Y. Leung et C. N. Ellis, « Recombinant interferon gamma therapy for atopic dermatitis », Journal of the American Academy of Dermatology, vol. 28, no 2 Pt 1, , p. 189–197 (ISSN 0190-9622, PMID 8432915, DOI 10.1016/0190-9622(93)70026-p, lire en ligne, consulté le )
- K. Abiko, N. Matsumura, J. Hamanishi et N. Horikawa, « IFN-γ from lymphocytes induces PD-L1 expression and promotes progression of ovarian cancer », British Journal of Cancer, vol. 112, no 9, , p. 1501–1509 (ISSN 1532-1827, PMID 25867264, PMCID 4453666, DOI 10.1038/bjc.2015.101, lire en ligne, consulté le )
- Ali Razaghi, Carina Villacrés, Vincent Jung et Narges Mashkour, « Improved therapeutic efficacy of mammalian expressed-recombinant interferon gamma against ovarian cancer cells », Experimental Cell Research, vol. 359, no 1, 10 01, 2017, p. 20–29 (ISSN 1090-2422, PMID 28803068, DOI 10.1016/j.yexcr.2017.08.014, lire en ligne, consulté le )
- S. V. Kotenko, L. S. Izotova, B. P. Pollack et T. M. Mariano, « Interaction between the components of the interferon gamma receptor complex », The Journal of Biological Chemistry, vol. 270, no 36, , p. 20915–20921 (ISSN 0021-9258, PMID 7673114, DOI 10.1074/jbc.270.36.20915, lire en ligne, consulté le )
- Daniel A. Leon Rodriguez, F. David Carmona, Luis Eduardo Echeverría et Clara Isabel González, « IL18 Gene Variants Influence the Susceptibility to Chagas Disease », PLoS neglected tropical diseases, vol. 10, no 3, , e0004583 (ISSN 1935-2735, PMID 27027876, PMCID 4814063, DOI 10.1371/journal.pntd.0004583, lire en ligne, consulté le )
- Ewa Trznadel-Grodzka, Marcin Błaszkowski et Helena Rotsztejn, « Investigations of seborrheic dermatitis. Part I. The role of selected cytokines in the pathogenesis of seborrheic dermatitis », Postepy Higieny I Medycyny Doswiadczalnej (Online), vol. 66, , p. 843–847 (ISSN 1732-2693, PMID 23175340, DOI 10.5604/17322693.1019642, lire en ligne, consulté le )
- Yitzhak Ben-Asouli, Yona Banai, Yehuda Pel-Or et Alexei Shir, « Human interferon-gamma mRNA autoregulates its translation through a pseudoknot that activates the interferon-inducible protein kinase PKR », Cell, vol. 108, no 2, , p. 221–232 (ISSN 0092-8674, PMID 11832212, DOI 10.1016/s0092-8674(02)00616-5, lire en ligne, consulté le )
- Ananthi J. Asirvatham, Christopher J. Gregorie, Zihua Hu et William J. Magner, « MicroRNA targets in immune genes and the Dicer/Argonaute and ARE machinery components », Molecular Immunology, vol. 45, no 7, , p. 1995–2006 (ISSN 0161-5890, PMID 18061676, PMCID 2678893, DOI 10.1016/j.molimm.2007.10.035, lire en ligne, consulté le )
- Chih-Hao Chang, Jonathan D. Curtis, Leonard B. Maggi et Brandon Faubert, « Posttranscriptional control of T cell effector function by aerobic glycolysis », Cell, vol. 153, no 6, , p. 1239–1251 (ISSN 1097-4172, PMID 23746840, PMCID 3804311, DOI 10.1016/j.cell.2013.05.016, lire en ligne, consulté le )
Articles connexes
Bibliographie
- Jeff G. Leid, Carey J. Willson, Mark E. Shirtliff, Daniel J. Hassett, Matthew R. Parsek and Alyssa K. Jeffers (2005) The Exopolysaccharide Alginate Protects Pseudomonas aeruginosa Biofilm Bacteria from IFN- γ-Mediated Macrophage Killing ; J Immunol 2005; 175:7512-7518 ;doi: 10.4049/jimmunol.175.11.7512 http://www.jimmunol.org/content/175/11/7512