Réplication virale
La réplication virale est l'ensemble des processus biochimiques qui se déroulent dans la cellule infectée par un virus et qui ont pour effet de produire de nouvelles unités de ce virus (ou virions). Ce mode de multiplication parasitaire qui exploite le mécanisme de réplication de l'ADN est ce qui définit les virus.
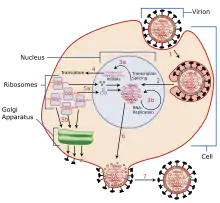
Les virus doivent d'abord pénétrer dans la cellule avant que la réplication virale puisse se produire. Grâce à la génération de copies abondantes de son génome et à l'encapsidation de ces copies, le virus peut continuer à infecter de nouveaux hôtes. La réplication des virus au sein des cellules est très variée et dépend du type de gènes viraux impliqués. La plupart des virus à ADN s'assemblent dans le noyau, tandis que la plupart des virus à ARN se développent uniquement dans le cytoplasme[1].
Cycle de réplication virale classique
Chaque virus possède des modes de réplication bien particuliers, notamment selon qu'il s’agit d’un virus à ADN ou d’un virus à ARN. Les virus ne se multiplient que dans les cellules vivantes. La cellule hôte doit fournir l'énergie et la machinerie de synthèse ainsi que les précurseurs de faible poids moléculaire pour la synthèse des protéines virales et des acides nucléiques[2]. La réplication virale n'est présentée ici que dans ses grandes lignes et comprend sept étapes, qui sont :
- Adsorption ;
- Pénétration dans la cellule-hôte ;
- Décapsidation ;
- Réplication du génome ;
- Synthèse des composants viraux ;
- Assemblage et encapsidation ;
- Libération.
Adsorption
C'est la première étape de la réplication virale. Le virus se fixe à la membrane de la cellule hôte et y injecte ensuite son ADN ou son ARN pour déclencher l'infection. Dans les cellules animales, ces virus entrent par le processus d'endocytose qui fonctionne par fusion du virus (enveloppe virale) avec la membrane cellulaire ; dans les cellules végétales, il entre par le processus de pinocytose qui fonctionne par « coincement » de l'enveloppe virale avec la paroi de la cellule.
Au cours de cette étape et dans les deux cas précédemment cités, il y a liaison d’une protéine de l'enveloppe virale à un récepteur de la membrane cellulaire. Les récepteurs des cellules eucaryotes peuvent être soit des glycoprotéines, soit des glycosphingolipides. Les récepteurs des bactériophages sont des glycoprotéines ou des lipopolysaccharides. Les cellules végétales ne possèdent pas de récepteurs spécifiques aux virus.
Pénétration
Selon les virus, il existe plusieurs mécanismes de pénétration à l’intérieur de la cellule. Chez les bactériophages, seul le génome viral pénètre dans la cellule bactérienne. Chez les virus animaux le virus peut pénétrer par plusieurs mécanismes. Le virus peut pénétrer par pinocytose : la nucléocapside virale, entourée de la membrane plasmique pénètre dans la cellule via une vacuole pinocytotique. Cela protège la cellule contre les anticorps, comme dans le cas du VIH[3]. C’est souvent le cas des virus nus. Dans le cas des virus enveloppés, le virus peut pénétrer soit par fusion (entre l’enveloppe virale et la membrane plasmique cellulaire) soit par endocytose (accumulation de particules virales dans des vésicules cytoplasmiques).
Décapsidation
Des enzymes cellulaires (provenant des lysosomes) éliminent l'enveloppe protéique du virus. Cela libère ou rend accessible l'acide nucléique ou le génome du virus. Selon les virus, la décapsidation peut avoir lieu dans le cytoplasme ou dans le noyau.
Réplication du génome
Selon les types de virus et la nature de leur génome, cette étape peut varier. Pendant ce cycle (transcription puis traduction qui suit), il est impossible d'isoler une particule virale.
Chez certains virus à ARN, l'ARN infectant produit de l'ARN messager (ARNm). Il va servir ensuite à la traduction du génome en produits protéiques (voir étape suivante). Pour d'autres virus, comme ceux à ARN ou ADN à brin négatif, les virus sont produits par intégration du génome dans celui de la cellule-hôte (rétrotranscription de l'ARN en ADN puis insertion dans l'ADN de la cellule-hôte dans le cas des virus à ARN à brin négatif, ou insertion directe de l'ADN viral dans l'ADN de la cellule-hôte dans le cas des virus à ADN à brin négatif), puis transcription de l'ADN de la cellule-hôte par sa machinerie de réplication habituelle. Les ARNm résultants contiendront des ARNm issus du virus, permettant de produire des protéines virales.
Synthèse des composants viraux (protéines et acide nucléique)
Les composants suivants sont fabriqués par le virus par le biais des organites existants dans la cellule-hôte.
- Synthèse des protéines virales : l'ARNm du virus est traduit au niveau des ribosomes des cellules en deux types de protéines virales :
- structurelles : les protéines qui composent la particule virale sont fabriquées et assemblées ;
- non-structurelles : on ne les trouve pas dans la particule virale ; ce sont principalement des enzymes pour la réplication du génome du virus (polymérase par exemple).
- Synthèse de l'acide nucléique viral (réplication du génome) : le nouveau génome du virus est synthétisé ; les matrices sont soit le génome parental, soit, dans le cas des génomes d'acide nucléique simple brin, des brins complémentaires nouvellement formés par le virus (appelés polymérats ou réplicats) par une enzyme cellulaire. Cela se produit dans les cellules qui se divisent rapidement.
Assemblage et encapsidation
Les virions sont assemblés et maturés dans les cellules infectées. Il y a encapsidation du génome avec les protéines virales pour former de nouvelles particules virales.
Cela peut se produire dans le noyau de la cellule, le cytoplasme ou la membrane plasmique pour la plupart des virus enveloppés. Les virus enveloppés acquièrent leur enveloppe par bourgeonnement, au détriment de la membrane plasmique ou de la membrane nucléaire de la cellule-hôte.
Libération des virions
Les virions reconstitués sont libérés à l'extérieur de la cellule. Cela se produit soit par rupture soudaine de la cellule, soit par extrusion progressive (bourgeonnement) de virus enveloppés à travers la membrane cellulaire.
Les nouveaux virus peuvent envahir ou attaquer d'autres cellules, ou rester dormants dans la cellule-hôte originelle. Dans le cas des bactériophages, la libération des virions nouvellement formés a lieu par lyse de la bactérie infectée. Cependant, dans le cas des virus animaux, la libération se fait généralement sans lyse des cellules.
Cas particulier des Poxvirus
- Article principal Réplication des Poxvirus
La réplication des Poxviridae est inhabituelle pour des virus à ADN double brin (ADNdb) parce qu'elle se produit dans le cytoplasme.
Cas particulier des virus géants
Le mimivirus a la particularité de se répliquer par phagocytose[4].
Système de classification de Baltimore
- Article principal Classification Baltimore
Les virus sont classés en 7 types de gènes, chacun d'eux ayant ses propres familles de virus, qui à leur tour ont des stratégies de réplication différentes. David Baltimore, biologiste lauréat du prix Nobel, a conçu un système de classification pour trier les différents virus en fonction de leur stratégie de réplication unique. Il existe sept stratégies de réplication différentes basées sur ce système (Classe I, II, III, IV, V, VI, VII). Les sept classes de virus sont énumérées ici de façon succincte et générale[5].
Classe I : virus à ADN double brin
Ce type de virus doit généralement entrer dans le noyau de l'hôte avant de pouvoir se répliquer. Certains de ces virus ont besoin de polymérases de la cellule hôte pour répliquer leur génome, tandis que d'autres comme les adénovirus ou les herpèsvirus codent leurs propres facteurs de réplication. Cependant, dans les deux cas, la réplication du génome viral dépend fortement d'un état cellulaire permettant la réplication de l'ADN, et donc du cycle cellulaire. Le virus peut amener la cellule à subir une division cellulaire forcée, qui peut conduire à la transformation de la cellule et, finalement, au cancer. Un exemple de famille de cette classification est celle des adénovirus.
Il n'existe qu'un seul exemple bien étudié dans lequel une famille de virus de classe I ne se réplique pas dans le noyau : il s'agit de la famille des Poxvirus, qui comprend des virus hautement pathogènes qui infectent les vertébrés (voir plus haut).
Classe II : virus à ADN simple brin
Les virus qui entrent dans cette catégorie comprennent ceux qui ne sont pas aussi bien étudiés que les précédents, mais qui ont tout de même une forte incidence sur les vertébrés. Les Circoviridae et Parvoviridae en sont deux exemples. Ils se répliquent dans le noyau et forment un ADN double brin intermédiaire lors de la réplication. Un Anellovirus humain appelé TTV est inclus dans cette classification et se retrouve chez presque tous les humains, les infectant de manière asymptomatique dans presque tous les organes principaux.
Classe III : virus à ARN double brin
Comme la plupart des virus à ARN, les virus à ARN double brin ne dépendent pas de la polymérase de l'hôte pour leur réplication, contrairement aux virus à ADN. Les virus à ARN double brin ne sont pas aussi bien étudiés que les autres classes. Cette classe comprend deux grandes familles, les Reoviridae et les Birnaviridae. La réplication est monocistronique et comprend des génomes individuels et segmentés, ce qui signifie que chacun des gènes ne code qu’une seule protéine, contrairement aux autres virus qui présentent une traduction plus complexe.
Classes IV et V : virus à ARN simple brin
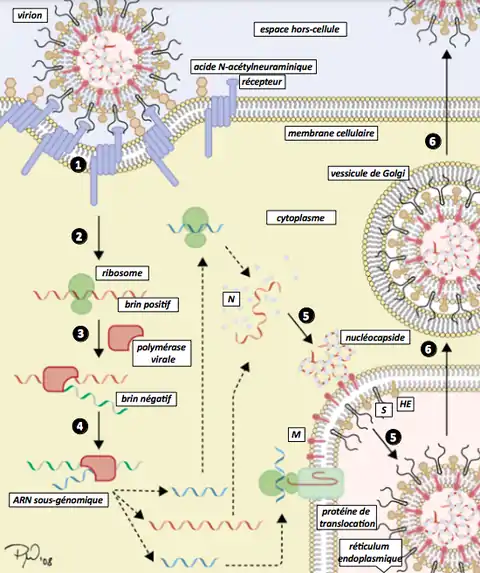
Ces virus consistent en deux types différents, mais tous deux partagent le fait que leur réplication se déroule principalement dans le cytoplasme, et que la réplication ne dépend pas autant du cycle cellulaire de la cellule-hôte que celle des virus à ADN. Ces classes de virus comprennent également parmi les types de virus les plus étudiés, avec les virus à ADN double brin.
Classe IV : virus à ARN simple brin à sens positif
Les virus à ARN à sens positif (et en fait tous les gènes définis comme à sens positif) peuvent être directement accessibles par les ribosomes de l'hôte pour former immédiatement des protéines. Ils peuvent être divisés en deux groupes, qui se répliquent tous deux dans le cytoplasme :
- les virus à ARNm polycistronique où l'ARN du génome forme l'ARNm puis est traduit en un produit polyprotéique qui est ensuite clivé pour former les protéines matures. Cela signifie que le gène peut utiliser plusieurs méthodes pour produire des protéines à partir du même brin d'ARN, réduisant ainsi la taille de son génome ;
- les virus à transcription complexe, qui peuvent utiliser des ARNm sous-génomiques, le déphasage ribosomique et le traitement protéolytique des polyprotéines. Tous ces mécanismes sont différents et permettent de produire des protéines à partir d'un même brin d'ARN.
Les familles des Coronaviridae, Flaviviridae et Picornaviridae sont des exemples de virus appartenant à cette classe.
Classe V : virus à ARN simple brin à sens négatif
Les virus à ARN à sens négatif (et en fait tous les gènes définis comme à sens négatif) ne peuvent pas être directement accessibles par les ribosomes de l'hôte pour former immédiatement des protéines. Ils doivent alors être transcrits par des polymérases virales dans le sens positif complémentaire "lisible". On peut également les diviser en deux groupes :
- les virus contenant des génomes non segmentés pour lesquels la première étape de la réplication est la transcription à partir du génome à brin négatif par l'ARN polymérase virale dépendante de l'ARN pour produire des ARNm monocistroniques qui codent les différentes protéines virales. Une copie du génome à sens positif qui sert de modèle pour la production du génome de brin négatif est ensuite produite. La réplication se fait dans le cytoplasme ;
- les virus à génomes segmentés pour lesquels la réplication se produit dans le cytoplasme et pour lesquels l'ARN polymérase virale dépendante de l'ARN produit des ARNm monocistroniques à partir de chaque segment de génome.
Les familles des Orthomyxoviridae, Paramyxoviridae, Bunyaviridae, Filoviridae et Rhabdoviridae (qui comprend la rage) sont des exemples de virus appartenant à cette classe.
Classe VI : virus à ARN simple brin à sens positif utilisant un ADN intermédiaire pour la réplication
Une famille bien étudiée de cette classe de virus comprend les rétrovirus. L'une de leurs caractéristiques est l'utilisation de la transcriptase inverse pour convertir l'ARN à sens positif en ADN. Au lieu d'utiliser l'ARN pour créer les modèles de traduction des protéines, ils utilisent l'ADN pour créer ces modèles, qui sont épissés dans le génome de l'hôte à l'aide d'une intégrase. La réplication peut alors commencer à l'aide des polymérases de la cellule hôte.
Classe VII : virus à ADN double brin utilisant un ARN simple brin intermédiaire pour la réplication
Ce petit groupe de virus, dont le virus de l'hépatite B est un exemple, possède un génome double-brin à lacunes qui sont ensuite remplies pour former un ADN circulaire fermé de manière covalente (ADNcfc ou cccDNA en anglais) qui sert de modèle pour la production d'ARNm viraux et d'un ARN sous-génomique. L'ARN pré-génome sert de modèle pour la transcriptase inverse virale et pour la production du génome de l'ADN.
Voir aussi
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Viral replication » (voir la liste des auteurs).
Références
- (en) R.J. Roberts, Fish pathology, Elsevier Health Sciences, , 3e éd.
- Geo F. Brooks et al., Jawetz Melnick & Adelberg's Medical Microbiology, McGraw Hill, , 26e éd., 880 p. (ISBN 978-0-07-181578-9)
- Rosangela G. Lima, Johan Van Weyenbergh, Elvira M. B. Saraiva et Manoel Barral-Netto, « The Replication of Human Immunodeficiency Virus Type 1 in Macrophages Is Enhanced after Phagocytosis of Apoptotic Cells », The Journal of Infectious Diseases, vol. 185, no 11, , p. 1561–1566 (ISSN 0022-1899, lire en ligne, consulté le )
- (en) Ameobal Pathogen Mimivirus Infects Macrophages through Phagocytosis
- (en) N.J. Dimmock et al., Introduction to Modern Virology, Blackwell Publishing,