Eremiasaurus
Eremiasaurus heterodontus

Eremiasaurus (littéralement « lézard du désert ») est un genre éteint de mosasaures ayant vécu durant la fin du Crétacé supérieur (Maastrichtien) dans ce qui est actuellement l'Afrique du Nord. Une seule espèce est connue, Eremiasaurus heterodontus, décrite en 2012 à partir de deux spécimens fossiles plus ou moins complets, découverts dans le bassin d'Ouled Abdoun, au Maroc. Ce site est connue pour avoir livré un nombre importants d'autres mosasaures apparentés.
Eremiasaurus est un représentant de taille moyenne pour un mosasaure, étant estimé à environ 5 m de long d'après les observations effectué sur les spécimens syntypes. Le crâne d'Eremiasaurus est robuste et est caractérisée de par son hétérodontie très différencié. L'anatomie des vertèbres caudales d'Eremiasaurus suggère qu'il aurait été un prédateur capable de nager à une vitesse élevée.
Eremiasaurus à vécu dans la marge sud de la Téthys méditerranéenne. Ce paléo-océan présentait un diversité importante de vertébrés aquatiques et était de climat océanique tempéré et chaud. Les archives fossiles montrent également qu'il y aurait eu une partition de niche entre Eremiasaurus et les divers autres espèces de mosasaures identifiés au sein du bassin d'Ouled Abdoun.
Historique des recherches
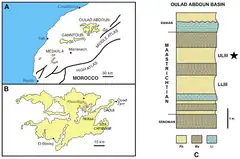
En 2012, les paléontologues Aaron R. H. LeBlanc (d), Michael W. Caldwell (d) et Nathalie Bardet (d) décrivent officiellement Eremiasaurus heterodontus sur la base de deux spécimens découverts au sein des gisements de phosphate du bassin d'Ouled Abdoun au Maroc, et plus précisément dans la zone de Sidi Daoui, situé à proximité de la ville d'Oued Zem. Les syntypes, catalogués respectivement UALVP 51744 et OCP DEK/GE 112, sont deux spécimens fossiles plus ou moins complets représentants presque tout le squelette connu du genre. La justification de l'utilisation des syntypes plutôt qu'un seul holotype provient du fait qu'UALVP 51744, le spécimen le plus complet, est dérivé du champ commercial sans données de localité précises, alors qu'OCP DEK/GE 112 fut exhumé par l'une des paléontologues, Nathalie Bardet, et dont la présence géographique et stratigraphique est connue. La zone précise concernant cette découverte se situe au niveau de la Couche III, datant du Maastrichtien supérieur[1] - [2], zone où d'autres mosasauridés ont été trouvé, notamment les imposantes espèces Mosasaurus beaugei[2] et Thalassotitan atrox[3].
Le nom de genre Eremiasaurus vient du grec ancien ἐρημία / eremia « désert » et σαῦρος / saûros « lézard », le tout voulant littéralement dire « lézard du désert », en référence au climat aride du Maroc actuel d'où le reptile marin a été découvert. L'épithète spécifique heterodontus provient également du grec ancien ἕτερος / heteros « différent » et ὀδόντος / odóntos « dent », les deux réunis voulant dire « dents différentes », faisant référence au changement drastique de la forme des dents sur divers points des mâchoires[1].
Description
Taille
Les syntypes d'Eremiasaurus indique une taille plutôt moyenne pour un mosasauridé. Le spécimen le plus complet, UALVP 51744, mesure 4,5 m de long, le crâne mesurant 63,5 cm, mais l'absence de plusieurs vertèbres dorsales suggère que la longueur réelle du corps aurait été plus proche de 5 m. Le second spécimen, OCP DEK/GE 112, fut d'abord estimé à une longueur atteignant les 6 m, taille basée sur l'observation du crâne qui est plus grand, mesurant 70 cm[1]. Cependant, une révision des mosasauridés provenant du Maroc menée par Bardet et ses collègues en 2015 réduit la taille proposée pour ce spécimen à 5 m[2].
Crâne
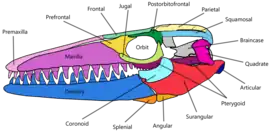
Le crâne d'Eremiasaurus est construit de manière robuste, mais pas autant que ceux de certaines espèces attribuées au genre apparenté Prognathodon[2]. Le rostre du prémaxillaire est court et conique en vue dorsale et s'étend vers l'avant sur moins de la longueur d'un espace alvéolaire. Le contour de la surface dorsale du prémaxillaire est étroite et pointue vers l'avant, semblable à Mosasaurus et à Plesiotylosaurus. Le maxillaire est long et fin. Les foramens nourriciers tapissent la surface latérale du maxillaire et s'agrandissent vers l'arrière. Ces foramens sont situés à environ 2 cm en arrière de la rangée de dents des maxillaires[1]. La partie préorbitaire est légèrement plus grande que la moitié de la longueur totale du crâne et la région frontale est élargie[2]. La surface latérale du préfrontal est concave et possède une surface dorsale large et plate pour le contact avec le frontal et le maxillaire qui se chevauchent. Postérieur à l'extrémité de la suture prémaxillaire-maxillaire, la barre internarial (la longue partie du prémaxillaire s'étendant derrière les dents de cet os) est resserré latéralement et a une quille dorsale médiane proéminente s'étendant le long de la moitié postérieure. Un grand processus supraorbitaire et triangulaire arrondi s'étend de la surface postéro-dorsale du préfrontal, similaire aux autres membres des mosasaurinés. Les processus infrastapédiens et suprastapédiens du carré sont fusionnés, possédant une fosse stapédienne très grande et arrondie, cette dernière étant l'une des autapomorphies de l'espèce[1].
La mâchoire inférieure est mince par rapport à la taille des dents logées le long du dentaire. La marge dorsale du dentaire est légèrement convexe en vue latérale. Cette convexité n'est pas aussi prononcée que chez la plupart des espèces de Prognathodon et ressemble plutôt à celle chez Mosasaurus et P. kianda. Le coronoïde porte un grand processus postérieur orienté verticalement, ce qui donne à la marge dorsale de l'os un angle de près de 90° entre l'extrémité antérieure horizontale et l'aile postérieure verticale. Les os articulaires forment de larges prolongements rectangulaires des mâchoires inférieures derrière les fosses glénoïdes. Cas exceptionnel chez les mosasaures, Eremiasaurus semble avoir un os hyoïde, un élément rarement retrouvé dans les fossiles des représentants de ce groupe. Cet os est légèrement élargi dans sa partie postérieure[1].
Dentition
Eremiasaurus est caractérisée de par son hétérodontie très marqué, les dents se différenciants dans de nombreux points au niveau des mâchoires de par leur forme et de taille[1] - [2]. Les dents des prémaxillaires auraient été pointés de manière droite plutôt que de faire saillie vers l'avant, chose qui est basée d'après les observations d'une petite fosse antérieure dans la première dent des os dentaires, ainsi que la position de la première alvéole du prémaxillaire[1].
Treize positions de dents peuvent être comptés dans les maxillaires gauche et droit chez Eremiasaurus[N 1]. Les trois dents antérieures d'UALVP 51744 ressemblent à celles du prémaxillaire et sont longues, droites et coniques, en forme de larme à rond en vue transversale, et ne possèdent que des carènes antérieures dentelées. Ces dents sont interdigités avec celles du dentaire et se logent dans de petites fosses entre les dents des dentaires adjacentes lorsque les mâchoires sont fermées. Ce type de contact se trouve également le long de la marge dentaire du maxillaire. Les couronnes sont en forme de lame, bicarinées avec un angle intercarinal de 180° et sont beaucoup plus longues d'avant en arrière que dans la dentition antérieure. Les surfaces d'émail sont lisses avec de faibles traces de facettes s'étendant jusqu'aux deux tiers de la hauteur d'une couronne dentaire donnée. De plus, l'expansion antéropostérieure de la couronne dentaire laisse peu de place entre les dents adjacentes et, à la septième dent du maxillaire, les fosses interdentaires sont absentes. À partir de ce point précis, les dents des maxillaires et des dentaires ne s'interdigitent plus, mais se cisaillent les unes contre les autres de sorte que les dents des maxillaires cachent les dents inférieures en vue latérale lorsque les mâchoires sont fermées. Les quatrième à neuvième dents maxillaires deviennent progressivement plus comprimés latéralement et légèrement incurvés. Les dixième à treizième dents des maxillaires sont également bicarinées, mais présentent beaucoup moins de compression latérale et sont également asymétriques et élargi antérieurement. En conséquence, les faces antérieures de ces dents sont convexes en vue latérale. Ces couronnes sont également évasées à leurs bases et ont des surfaces lisses en émail[1].
Les os dentaires possèdent quinze dents de chaque cotés. Le modèle d'hétérodontie dans la dentition inférieure reflète celui du prémaxillaire et des maxillaires. Les cinq premiers dents sont droites et coniques et chacune n'a qu'une seule dent dentée et une carène antérieure. Les sixième à neuvième dents sont plus comprimées latéralement et en forme de lame, et chacune a une carène dentelée sur les bords antérieur et postérieur. Ces couronnes dentaires sont également plus convexes le long de leurs surfaces linguaux (le côté tourné vers la langue) que le long de leurs côtés labiaux (le côté tourné vers l'extérieur). Les dixième à quinzième dents sont plus bulbeuses et courbes, et possèdent encore deux carènes avec un angle intercarinal de 180°. Il y a de petites dépressions le long de la marge dentaire entre les dents adjacentes, qui disparaissent en arrière de la huitième position des dents des os dentaires[1].
Une rangée provenant d'un os ptérygoïde se compose de cinq dents et d'une dent isolée mais associée. La dent isolée appartient à la même rangée de dents des ptérygoïdes, sur la base d'une courbure identique des couronnes. Les dents des ptérygoïdes sont longues, approchant les tailles des dents marginales postérieures, mais sont nettement plus petits que les couronnes marginales médianes. Les deux dents antérieures sont fines, droites et coniques, tandis que les trois dernières deviennent progressivement plus petit et plus courbé. La dent isolée est plus petite que la denture marginale et a une base élargie de la couronne. Le degré marqué d'hétérodontie dans la dentition des os ptérygoïdes reflète des changements similaires dans les dents marginales, où les couronnes deviennent plus petites, plus bulbeuses et incurvée plus en arrière. De plus, la section transversale des zones des bases des dents ptérygoïdiens chez Eremiasaurus ne s'agrandissent pas vers l'avant, contrairement à ce à quoi on pourrait s'attendre pour toutes les espèces provisoirement classées au sein du genre Prognathodon[1].
Squelette postcrânien
Le nombre exact de vertèbres chez Eremiasaurus est inconnue, même le spécimen UALVP 51744, le fossile le plus complet connu, étant dépourvue de nombreuses parties des vertèbres dorsales et caudales. Mais en excluant ceci, certaines vertèbres et autres parties postcrâniens de l'animal sont bien préservée et permettent de le visualiser dans son ensemble. Les côtes semble devenirs de plus en plus petites en avançant vers l'arrière des vertèbres. Au niveau de la deuxième vertèbre dorsale, les côtes sont nombreuse et larges, tandis que certaines de la troisième sont courtes et fines. Plusieurs grandes côtes thoraciques cachent la marge dorsale de la lame scapulaire en vue latérale, masquant l'arc formé par le bord dorsal de l'omoplate[1].
Eremiasaurus possède plusieurs caractéristiques semblable à Plotosaurus au niveau des vertèbres caudales. Dans les caractéristiques les plus notables figurent la présence d'une déviation ventrale de la queue, un éventail des épines neurales caudales et une série inhabituellement longue de vertèbres pygales. Le grand nombre de vertèbres pygales, accompagné d'un nombre réduit de caudales intermédiaires est unique chez Eremiasaurus. La présence d'une déviation ventrale de la queue, similaire à celle de Plotosaurus, et un large éventail d'épines neurales dans la même région de la série vertébrale caudale suggèrent un développement similaire d'une lobe de la nageoire dorsale naissante chez Eremiasaurus, quoique moins développé. Ces changements dans les proportions, telles qu'une augmentation du nombre de vertèbres pygales, suggèrent un prédateur de poursuite à grande vitesse, convergente avec les proportions vertébrales de Plotosaurus[1].
L'omoplate et le coracoïde semble être étroitement suturé en avant de la fosse glénoïde. Cette même fosse est d'ailleurs légèrement bombé, une autre caractéristique distinctive du taxon. En arrière de cette fosse, le bord postérieur de l'omoplate s'étend dorsalement avant de s'étendre jusqu'à former la marge postérieure de la lame scapulaire, semblable à Clidastes. La face latérale de l'omoplate est large, lisse et plate. L'humérus est de hauteur et de largeur subégales, contrairement à Mosasaurus et à Plotosaurus, où ils sont plus larges que haut. Comme tous les mosasauridés, la crête iliaque est réduite à un processus cylindrique incliné vers l'avant. L'extrémité distale de l'ilium est élargie et porte des facettes pour l'articulation avec le pubis et l'ischion. Le tibia est un élément rectangulaire, plus long proximodistalement qu'antéropostérieurement. Le fibula est en forme de cloche, avec l'extrémité distale étant beaucoup plus large que l'extrémité proximale, contrairement à Mosasaurus et à Plotosaurus, où les deux extrémités du fibula sont de largeur inférieure. Cet os fait aussi environ les trois quarts de la longueur du tibia[1], contrairement à Tylosaurus et à Platecarpus, dans lequel le péroné est de la même longueur que le tibia[4]. Le plus large des éléments connus du tarse est interprété comme étant un astragal. Cet os est en forme de rein, avec une facette fibulaire pédonculée sur la face dorsale. Les phalanges sont allongés en forme de fuseau avec des épiphyses modérément élargies, différents des proportions robustes et en forme de bloc observées chez Mosasaurus et Plotosaurus[1].
Classification


Dans l'un des nombreux cladogrammes présents dans la description formelle du taxon menée par LeBlanc et ses collègues en 2012, Eremiasaurus est placée en tant que groupe frère de la tribu des Plotosaurini[1]. Plotosaurini est un taxon qui fut érigée pour la première fois en 1967 par Dale Russell dans son ouvrage scientifique intitulée Systematics and morphology of American mosasaurs, d'où il révise la plupart des mosasauridés nord-américains[4]. La validité de cette tribu commence à remise en question à partir de 1997, année d'où la révision phylogénétique des Mosasauroidea menée par Gorden L. Bell Jr. est officiellement publiée, la considérant comme polyphylétique et donc obsolète[5]. L'étude de LeBlanc et al. (2012) fait valoir que, bien qu'il ne soit pas nécessairement invalide, l'abandon de Mosasaurini ne suivrait pas le principe général du genre type reporté à tous les rangs dans une hiérarchie de classification, et que les diagnostics originaux des Plotosaurini sont aujourd'hui obsolètes. Les auteurs proposent donc de synonymiser le taxon Plotosaurini vers Mosasaurini, et placent Eremiasaurus dans un clade contenant Mosasaurus et Plotosaurus sur la base de divers caractéristiques crâniennes. Les caractéristiques justifiant cette classification incluent : la présence d'une quille dans la barre internarial ; l'exclusion des préfrontaux des bordures nariales ; des nariales enfoncé dans le frontal ; et la présence d'une rainure dans l'os carré[N 2] - [1]. La validité des Mosasaurini sera immédiatement accepté, et le placement d'Eremiasaurus tel que proposée par LeBlanc et ses collègues en 2012 sera maintenue dans divers études ultérieurs[6] - [7] - [8] - [9].
En 2017, une étude révisant la phylogénie des mosasauroïdes utilisent plusieurs arbres afin de savoir si le regroupement est valide. Il s'avère que la plupart de leurs arbres gardaient cette position, mais d'autres analyses de parcimonie pondérée et l'analyse bayésienne ne supportent pas une telle topologie[10]. Une autre étude publié la même année par Daniel Madzia et Andrea Cau utilise également l'analyse bayésienne et montre un soutien extrêmement faible quand à cette réunification. Eremiasaurus est donc un taxon qui est toujours classé dans les Mosasaurini, mais pas dans la réunification tel que proposée par LeBlanc et al. (2012)[11].
Le cladogramme suivant est modifié à partir d'un arbre de crédibilité de clade maximum déduit par une analyse bayésienne dans la plus récente analyse phylogénétique majeure de la sous-famille des Mosasaurinae par Madzia & Cau (2017)[11], qui est auto-décrit comme un raffinement d'une étude plus large de Simões et al. (2017)[10] :
Paléoécologie
Les gisements du bassin d'Ouled Abdoun au Maroc constituait durant le Maastrichtien une grande partie de ce qui fut autrefois la marge sud de la Téthys méditerranéenne[2]. Situé le long de l'équateur, entre 20°N et 20°S, les climats tempérés était plus chaudes que la marge nord du paléo-océan, qui lui fut situé entre 30 à 40°N[12]. Ce lieu est représenté par une vaste diversité de vertébrés aquatiques, incluant divers poissons osseux comme Enchodus et Stratodus aux poissons cartilagineux comme Cretalamna, Squalicorax et Rhombodus (en)[3]. La localité précise d'où Eremiasaurus fut découverte était déjà connu pour avoir un nombre élevé d'autres mosasaures, avec plus de dix genres identifiés au sein du bassin d'Ouled Abdoun. L'anatomie dentaire grandement différenciés au sein des taxons suggère qu'une partition des niches aurait eu lieu, dans lequel les prédateurs occupaient différentes niches pour éviter la concurrence les uns avec les autres. Par exemple, certains mosasaures identifiés au sein de cette zone, tels que Globidens et Carinodens (en), ont des dents émoussées pour écraser les coquillages, alors que des mosasaures comme Mosasaurus, Thalassotitan et Prognathodon ont des dentitions spécialisées pour chasser des proies plus grosses[2] - [3]. Bien que le régime alimentaire particulier d'Eremiasaurus soit incertain, sa divergence avec d'autres mosasaurinés aux dents robustes suggérerait qu'il se spécialisait dans des sources de nourritures non exploitées par ses plus grands contemporains du Maroc[1] - [2]. D'autres squamates marins y sont connus, notamment le varan marin Pachyvaranus et possiblement le serpent de mer Palaeophis (en). De nombreux tortues marines de la famille des Bothremydidae ont été identifiés. Les plésiosaures, à l'exception de Zarafasaura, sont rarement présents au sein de la localité[12].
Notes et références
Notes
- Dans le spécimen OCP DEK/GE 112, douze dents peuvent être comptés de chaque côtés du crâne, mais une possible treizième peut-être également présent postérieurement, car la matrice et les fragments d'os obscurcissent cette zone[1].
- L'étude de LeBlanc et al. (2012) décrit cette rainure sous le nom rainure ala[1].
Références
- (en) Aaron R. H. Leblanc, Michael W. Caldwell et Nathalie Bardet, « A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics », Journal of Vertebrate Paleontology, vol. 32, no 1, , p. 82-104 (DOI 10.1080/02724634.2012.624145, JSTOR 41407709, S2CID 130559113, lire en ligne)
- (en) Nathalie Bardet, Alexandra Houssaye, Peggy Vincent, Xabier Pereda Suberbiola, M'barek Amaghzaz, Essaid Jourani et Saïd Meslouh, « Mosasaurids (Squamata) from the Maastrichtian Phosphates of Morocco: Biodiversity, palaeobiogeography and palaeoecology based on tooth morphoguilds », Gondwana Research, vol. 27, no 3, , p. 1068-1078 (DOI 10.1016/j.gr.2014.08.014, Bibcode 2015GondR..27.1068B, S2CID 140596842, lire en ligne)
- (en) Nicholas R. Longrich, Nour-Eddine Jalil, Fatima Khaldoune, Oussama Khadiri Yazami, Xabier Pereda-Suberbiola et Nathalie Bardet, « Thalassotitan atrox, a giant predatory mosasaurid (Squamata) from the Upper Maastrichtian Phosphates of Morocco », Cretaceous Research, vol. 140, , p. 105315 (ISSN 0195-6671, DOI 10.1016/j.cretres.2022.105315, S2CID 251821884)
- (en) Dale A. Russell, Systematics and morphology of American mosasaurs, vol. 23, New Haven, Bulletin of the Peabody Museum of Natural History, , 240 p. (OCLC 205385, lire en ligne)
- (en) Gorden L. Bell Jr., « A Phylogenetic Revision of North American and Adriatic Mosasauroidea », dans Jack M. Callaway et Elizabeth L. Nicholls, Ancient Marine Reptiles, San Diego, Academic Press, , 501 p. (ISBN 978-0-12-155210-7, DOI 10.1016/b978-012155210-7/50017-x, S2CID 88638962), p. 293-332
- (en + ru) Dimitry V. Grigoriev, « Redescription of Prognathodon lutugini (Squamata, Mosasauridae) », Proceedings of the Zoological Institute RAS, vol. 317, no 3, , p. 246-261 (DOI 10.31610/trudyzin/2013.317.3.246
 , S2CID 189800203, lire en ligne [PDF])
, S2CID 189800203, lire en ligne [PDF]) - (en) Alessandro Palci, Michael W. Caldwell et Cesare A. Papazzoni, « A new genus and subfamily of mosasaurs from the Upper Cretaceous of northern Italy », Journal of Vertebrate Paleontology, vol. 33, no 3, , p. 599-612 (DOI 10.1080/02724634.2013.731024, JSTOR 42568543, S2CID 86646993)
- (en) Fedrico Fanti, Andrea Cau et Alessandra Negri, « A giant mosasaur (Reptilia, Squamata) with an unusually twisted dentition from the Argille Scagliose Complex (late Campanian) of Northern Italy », Cretaceous Research, vol. 49, no 2014, , p. 91-104 (DOI 10.1016/j.cretres.2014.01.003, S2CID 129797507, lire en ligne [PDF])
- (en) Paulina Jimenez-Huidobro et Michael W. Caldwell, « Reassessment and reassignment of the early Maastrichtian mosasaur Hainosaurus bernardi Dollo, 1885, to Tylosaurus Marsh, 1872 », Journal of Vertebrate Paleontology, vol. 36, no 3, , e1096275 (DOI 10.1080/02724634.2016.1096275, JSTOR 24740320, S2CID 87315531)
- (en) Tiago R. Simões, Oksana Vernygora, Ilaria Paparella, Paulina Jimenez-Huidobro et Michael W. Caldwell, « Mosasauroid phylogeny under multiple phylogenetic methods provides new insights on the evolution of aquatic adaptations in the group », PLOS ONE, vol. 12, no 5, , e0176773 (PMID 28467456, PMCID 5415187, DOI 10.1371/journal.pone.0176773 , Bibcode 2017PLoSO..1276773S)
- (en) Daniel Madzia et Andrea Cau, « Inferring 'weak spots' in phylogenetic trees: application to mosasauroid nomenclature », PeerJ, vol. 5, , e3782 (PMID 28929018, PMCID 5602675, DOI 10.7717/peerj.3782 )
- (en + fr) Nathalie Bardet, « Les reptiles marins du Maastrichtien de la Tethys méditerranéenne: une approche paléobiogéographique », Bulletin de la Société Géologique de France, vol. 183, no 6, , p. 573–596 (DOI 10.2113/gssgfbull.183.6.573, S2CID 140553167)
Voir aussi
Articles connexes
Liens externes
- (en) Paulina Jiménez-Huidobro, Tiago R. Simões et Michael W. Caldwell, « Mosasauroids from Gondwanan Continents », Journal of Herpetology, vol. 51, no 3, , p. 355-364 (DOI 10.1670/16-017, S2CID 89780058, lire en ligne)
- Ressources relatives au vivant :