Facteur de nécrose tumorale
Les facteurs de nécrose tumorale, ou TNF (de l'anglais : tumor necrosis factors), forment une superfamille de protéines dite superfamille des TNF, dont le membre type est le TNF α, également appelé cachectine, voire cachexine. Ce sont des protéines transmembranaires présentant un domaine homologue dit TNF. Le terme « facteur de nécrose tumorale » sans autre précision fait généralement référence au seul TNF α, importante cytokine impliquée dans l'inflammation systémique et dans la réaction de phase aiguë, qui sera développé dans cet article ; d'autres membres de la famille des TNF portent également d'autres noms, comme le TNF β, qui n'est autre que la lymphotoxine α.
| Facteur de nécrose tumorale α | ||
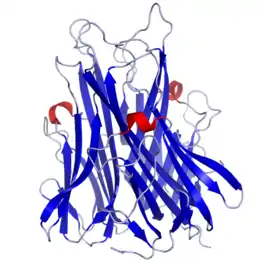 Structure cristallisée d'un TNF α humain (PDB 1TNF[2]). | ||
| Caractéristiques générales | ||
|---|---|---|
| Nom approuvé | Facteur de nécrose tumorale | |
| Symbole | TNF | |
| Synonymes | TNF α, cachectine, TNFSF2 | |
| Homo sapiens | ||
| Locus | 6p21.33 | |
| Masse moléculaire | 25 644 Da[3] | |
| Nombre de résidus | 233 acides aminés[3] | |
| Entrez | 7124 | |
| HUGO | 11892 | |
| OMIM | 191160 | |
| UniProt | P01375 | |
| RefSeq (ARNm) | NM_000594.3 | |
| RefSeq (protéine) | NP_000585.2 | |
| Ensembl | ENSG00000232810 | |
| PDB | 1A8M, 1TNF, 2AZ5, 2E7A, 2TUN, 2ZJC, 2ZPX, 3ALQ, 3IT8, 3L9J, 3WD5, 4G3Y, 4TSV, 4TWT, 4Y6O, 5M2I, 5M2J, 5M2M, 5MU8, 5TSW, 5UUI, 5WUX, 5YOY | |
|
GENATLAS • GeneTests • GoPubmed • HCOP • H-InvDB • Treefam • Vega | ||
| Liens accessibles depuis GeneCards et HUGO. | ||
Le TNF α est produit essentiellement par des macrophages activés, mais il peut également être produit par de nombreux autres types de cellules, comme les lymphocytes T auxiliaires, les lymphocytes NK, les granulocytes neutrophiles, les granulocytes éosinophiles, les mastocytes et les neurones. Il a principalement un rôle régulateur des leucocytes. Son action dans l'organisme déclenche la fièvre, l'apoptose, la cachexie, l'inflammation, en vue de combattre la cancérogenèse et la réplication virale ainsi que de répondre au sepsis en activant les cellules productrices d'interleukine 1 et d'interleukine 6. Le dérèglement de la production du TNF α a été impliqué dans un ensemble de maladies humaines telles que la maladie d'Alzheimer[4], le cancer[5], la dépression[6], le psoriasis[7] et les maladies inflammatoires chroniques de l’intestin[8] (MICI). Bien que controversées, certaines études relatives à la dépression et aux MICI relient ces conditions à des niveaux élevés de TNF[9] - [10].
Histoire et nomenclature
Le TNFα a été isolé en 1975 par Carswell et al. sous forme d'un facteur soluble libéré par les cellules de l'hôte ayant provoqué la nécrose d'une tumeur transplantée, la « sarcome Meth A »[11]. Bien que le TNFα ne provoque pas la nécrose de certaines tumeurs, il peut stimuler la croissance d'autres. Dans ce sens, son nom est quelque peu abusif.
Gène
Le gène du TNF α humain a été cloné en 1985[12]. Il se trouve sur le chromosome 6 en position 6p21.3, s'étend sur environ 3 kilobases et contient quatre exons. Le dernier exon présente des similitudes avec la lymphotoxine α, également appelée TNF β[13]. La région 3'-UTR de l'ARN messager du TNF α contient un élément riche en AU (en) (ARE).
Structure
Le TNF α est produit sous forme d'une protéine transmembranaire initialement longue de 233 résidus d'acides aminés formant des homotrimères stables[14] - [15]. La cytokine homotrimérique soluble TNFs est libérée à partir de cette forme membranaire par clivage protéolytique sous l'action d'une métalloprotéinase, l'enzyme de conversion du TNF α, appelée TACE ou ADAM17 (en)[16]. Le TNFs trimérique soluble de 51 kDa tend à se dissocier aux concentrations inférieures à la nanomole par litre, ce qui lui fait perdre en activité biologique. La forme sécrétée du TNF humain adopte une forme pyramidale pour une masse d'environ 17 kDa. Les formes membranaire et sécrétée sont biologiquement actives, mais les rôles respectifs de chacune d'entre elles demeurent débattus ; ce deux formes présentent néanmoins à la fois des fonctions distinctes et des fonctions communes[17].
Le TNF α fait partie d'un groupe de plusieurs cytokines impliquées dans l'inflammation, en phase de réaction aiguë et lors de l'inflammation chronique. C'est une glycoprotéine de 185 acides aminés, obtenue par clivage d'un précurseur de 212 acides aminés se trouvant à la surface de macrophages ou de fibroblastes. Certaines cellules sécrètent des isoformes plus ou moins longs. Le gène du TNFa est situé sur le chromosome 6 humain (en 6p21).
La structure du TNF α est constituée de deux feuillets β antiparallèles. Des ponts disulfure stabilisent la structure, mais ils ne sont pas nécessaires à l’activité biologique.
L'extrémité C-terminale est à l'intérieur du feuillet alors que l'extrémité N-terminale est libre à l'extérieur.
On dénombre deux formes de TNF α : une forme soluble et une forme liée à la membrane. Ces deux formes sont actives mais ont des affinités différentes pour les récepteurs membranaires au TNF (TNFR-1 et TNFR-2). Il existe également des récepteurs solubles, ou circulants, ils ont un rôle de leurre (decoy), entrant en compétition avec les récepteurs membranaires et réduisant ainsi l'activité biologique du TNF α.
Biologie
La fixation des TNF sur des récepteurs (TNFR 1 et 2) peut aboutir à au moins trois évènements :
- activation de la voie NF-κB (inhibition de l'apoptose)[18] ;
- induction de l'apoptose ;
- induction d'une nécrose programmée.
La fixation sur le TFNR1 permet la formation d'un complexe avec le TRADD, le RIP1 et le TRAF2, le tout permettant l'activation du NF-κB[19].
Physiopathologie
Le TNFα est libéré par des leucocytes, l'endothélium et d'autres tissus généralement en réponse à une lésion, et/ou lors d'une infection. Sa libération est stimulée par plusieurs autres médiateurs, comme l'interleukine 1 ou l'endotoxine bactérienne. Il possède plusieurs actions sur divers organes et systèmes, généralement en coopération avec les interleukines 1 et 6 :
- effets sur l'hypothalamus :
- stimulation du complexe hypothalamo-hypophysaire, via la libération de corticolibérine,
- suppression de l'appétence, faim (d'où son nom historique de « cachectine », son action a été rattachée à la perte de poids sévère observée lors de processus pathologiques, cancéreux en particulier). Son rôle dans les vomissements est associé à une catastrophe sanitaire : la thalidomide, molécule anti-TNF mais aussi tératogène, a été utilisée contre les vomissements associés à la grossesse, entraînant une vague de nombreuses malformations chez les nouveau-nés, avant d'être interdite,
- fièvre ;
- sur le foie : stimulation de la phase de réponse aiguë de l'inflammation, conduisant à une augmentation de CRP et d'autres médiateurs ;
- il attire très efficacement les polynucléaires neutrophiles, et les aide à adhérer à la paroi des cellules endothéliales, d'où elles sortiront par diapédèse ;
- sur les macrophages : stimulation de la phagocytose, production d'IL-1, d'oxydants et de lipides pro-inflammatoires, p.e la prostaglandine E2 (PGE2) ;
- autres tissus : augmentation de la résistance à l'insuline , stimulation de la différenciation des ostéoclastes[20].
Une augmentation locale de la concentration en TNFα est une des causes des signes cardinaux de l'inflammation : Rubor (rougeur, érythème), Calor (chaleur, due à la vasodilatation), Tumor (tuméfaction, œdème), Dolor (douleur).
TNF-α : rôle local et effet systémique
Le recrutement et l'activation des macrophages et des lymphocytes par le TNF sur le site d'une inflammation locale contribue à contenir et à éliminer les agents pathogènes ayant pénétré un tissu. Cependant en cas d'infection systémique (sepsis), le TNF-α peut être sécrété de façon excessive à la suite de la présence massive d'agents infectieux dans les tissus et/ou le sang. L'action locale (par exemple vasodilatation de capillaires sanguins proches d'un lieu d'infection) se propage donc à l'ensemble de l'organisme. Cela entraîne un état de choc, avec coagulation intravasculaire disséminée, défaillance de plusieurs organes nobles, pouvant entraîner la mort[21].
Pharmacologie
Depuis le début des années 1990 et grâce aux travaux de Jan Vilcek et de son groupe à la New York University School of Medicine, l'inhibition du TNFα par un anticorps monoclonal, comme l'infliximab[22] (Remicade) ou l'adalimumab (Humira), ou une protéine chimérisée avec le récepteur soluble au TNF (aussi appelée immunoadhésine), comme l'etanercept (Enbrel), fait partie de l'arsenal thérapeutique actuellement disponible dans de nombreuses maladies autoimmunes ou inflammatoires chroniques comme la polyarthrite rhumatoïde (PR), la spondylarthrite ankylosante (SA), la maladie de Crohn, la rectocolite hémorragique et le psoriasis. En inhibant le TNFα, ces drogues inhibent la réponse inflammatoire qui est la cause principale des manifestations cliniques de ces pathologies. Des essais cliniques quant à l'efficacité de ces drogues sur l'hidradenitis suppurativa (maladie de Verneuil) sont en cours. En 2010, un quatrième inhibiteur du TNF, le certolizumab pegol, a été approuvé pour utilisation humaine (Cimzia). De nouveaux essais cliniques sont également lancés par l'OMS pour mesurer leur efficacité contre le COVID-19.
Ces traitements sont immunosuppresseurs et peuvent augmenter le risque de contracter la tuberculose, ou la réactivation d'infections latentes.
Le TNF, ou ses effets, peuvent également être inhibés de façon plus modeste par certains composés naturels, dont la curcumine (présente dans la curcuma) et les catéchines (présentes dans le thé vert).
Médicaments anti-TNF
- Thalidomide : diminution de la sécrétion de TNF ;
- Étanercept (PR, psoriasis, SA) : neutralisation du TNF par des récepteurs solubles
- Infliximab (PR, Crohn, SA)
- Adalimumab : neutralisation du TNFα par des anticorps
- Certolizumab pegol (PR, Crohn, psoriasis, SA) : premier anti-TNF pegylé
Notes et références
- (en) M. J. Eck et S. R. Sprang, « The structure of tumor necrosis factor-alpha at 2.6 A resolution. Implications for receptor binding. », Journal of Biological Chemistry, vol. 264, no 29, , p. 17595-17605 (PMID 2551905, DOI 10.2210/pdb1tnf/pdb, lire en ligne)
- (en) M. J. Eck et S. R. Sprang, « The structure of tumor necrosis factor-alpha at 2.6 A resolution. Implications for receptor binding. », Journal of Biological Chemistry, vol. 264, no 29, , p. 17595-17605 (PMID 2551905, DOI 10.2210/pdb1tnf/pdb, lire en ligne)
- Les valeurs de la masse et du nombre de résidus indiquées ici sont celles du précurseur protéique issu de la traduction du gène, avant modifications post-traductionnelles, et peuvent différer significativement des valeurs correspondantes pour la protéine fonctionnelle.
- (en) Walter Swardfager, Krista Lanctôt, Lana Rothenburg, Amy Wong, Jaclyn Cappell et Nathan Herrmann, « A Meta-Analysis of Cytokines in Alzheimer's Disease », Biological Psychiatry, vol. 68, no 10, , p. 930-941 (PMID 20692646, DOI 10.1016/j.biopsych.2010.06.012, lire en ligne)
- (en) Richard M. Locksley, Nigel Killeen et Michael J. Lenardo, « The TNF and TNF Receptor Superfamilies », Cell, vol. 104, no 4, , p. 487-501 (PMID 11239407, DOI 10.1016/s0092-8674(01)00237-9, lire en ligne)
- (en) Yekta Dowlati, Nathan Herrmann, Walter Swardfager, Helena Liu, Lauren Sham, Elyse K. Reim et Krista L. Lanctôt, « A Meta-Analysis of Cytokines in Major Depression », Biological Psychiatry, vol. 67, no 5, , p. 446-457 (PMID 20015486, DOI 10.1016/j.biopsych.2009.09.033, lire en ligne)
- (en) Frank C. Victor et Alice B. Gottlieb, « TNF-alpha and Apoptosis: Implications for the Pathogenesis and Treatment of Psoriasis », Journal of Drugs in Dermatology, vol. 1, no 3, , p. 264-275 (PMID 12851985)
- (en) J. Brynskov, P. Foegh, G. Pedersen, C. Ellervik, T. Kirkegaard, A. Bingham et T. Saermark, « Tumour necrosis factor α converting enzyme (TACE) activity in the colonic mucosa of patients with inflammatory bowel disease », Gut, vol. 51, no 1, , p. 37-43 (PMID 12077089, PMCID 1773288, DOI 10.1136/gut.51.1.37, lire en ligne)
- (en) Antonina A. Mikocka-Walus, Deborah A. Turnbull, Nicole T. Moulding, Ian G. Wilson, Jane M. Andrews et Gerald J. Holtmann, « Controversies surrounding the comorbidity of depression and anxiety in inflammatory bowel disease patients: A literature review », Inflammatory Bowel Diseases, vol. 13, no 2, , p. 225-234 (PMID 17206706, DOI 10.1002/ibd.20062, lire en ligne)
- (en) Kinga Bobińska, Elżbieta Gałecka, Janusz Szemraj, Piotr Gałecki et Monika Talarowska, « Is There a Link Between TNF Gene Expression and Cognitive Deficits in Depression? », Acta Biochimica Polonica, vol. 64, no 1, , p. 65-73 (PMID 27991935, DOI 10.18388/abp.2016_1276, lire en ligne)
- (en) Carswell EA, Old LJ, Kassel RL, Green S, Fiore N, Williamson B, « An endotoxin-induced serum factor that causes necrosis of tumors », PNAS, vol. 72, , p. 3666-70 (PMID 1103152, lire en ligne)
- (en) Lloyd J. Old, « Tumor Necrosis Factor (TNF) », Science, vol. 230, no 4726, , p. 630-632 (PMID 2413547, DOI 10.1126/science.2413547, Bibcode 1985Sci...230..630O, lire en ligne)
- (en) Glenn E. Nedwin, Susan L. Naylor, Alan Y. Sakaguchi, Douglas Smith, Julie Jarrett-Nedwin, Diane Pennica, David V. Goeddel et Patrick W. Gray, « Human Lymphotoxin and tumor necrosis factor genes: structure, homology and chromosomal localization », Nucleic Acids Research, vol. 13, no 17, , p. 6361-6373 (PMID 2995927, DOI 10.1093/nar/13.17.6361, Bibcode 321958, lire en ligne)
- (en) M. Kriegler, C. Perez, K. DeFay, I. Albert et S. D. Lu, « A novel form of TNF/cachectin is a cell surface cytotoxic transmembrane protein: Ramifications for the complex physiology of TNF », Cell, vol. 53, no 1, , p. 45-53 (PMID 3349526, DOI 10.1016/0092-8674(88)90486-2, lire en ligne)
- (en) Ping Tang, Mien-Chie Hung et Jim Klostergaard, « Human pro-Tumor Necrosis Factor Is a Homotrimer », Biochemistry, vol. 35, no 25, , p. 8216-8225 (PMID 8679576, DOI 10.1021/bi952182t, lire en ligne)
- (en) Roy A. Black, Charles T. Rauch, Carl J. Kozlosky, Jacques J. Peschon, Jennifer L. Slack, Martin F. Wolfson, Beverly J. Castner, Kim L. Stocking, Pranitha Reddy, Subhashini Srinivasan, Nicole Nelson, Norman Boiani, Kenneth A. Schooley, Mary Gerhart, Raymond Davis, Jeffrey N. Fitzner, Richard S. Johnson, Raymond J. Paxton, Carl J. March et Douglas Pat Cerretti, « A metalloproteinase disintegrin that releases tumour-necrosis factor-α from cells », Nature, vol. 385, no 6618, , p. 729-733 (PMID 9034190, DOI 10.1038/385729a0, Bibcode 1997Natur.385..729B, lire en ligne)
- (en) Michael A. Palladino, Frances Rena Bahjat, Emmanuel A. Theodorakis et Lyle L. Moldawer, « Anti-TNF-α therapies: the next generation », Nature Reviews: Drug Discovery, vol. 2, no 9, , p. 736-746 (PMID 12951580, DOI 10.1038/nrd1175, Bibcode 2003Natur.002..736B, lire en ligne)
- (en) Sreerupa Challa et Francis Ka-Ming Chan, « Going up in Flames: Necrotic Cell Injury and Inflammatory Diseases », Cell Mol Life Sci, vol. 67, , p. 3241–53 (PMCID PMC3051829, lire en ligne)
- Micheau O, Tschopp J, Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes, Cell, 2003;114:181–190
- K. Kobayashi, N. Takahashi, E. Jimi et N. Udagawa, « Tumor necrosis factor alpha stimulates osteoclast differentiation by a mechanism independent of the ODF/RANKL-RANK interaction », The Journal of Experimental Medicine, vol. 191, no 2, , p. 275–286 (ISSN 0022-1007, PMID 10637272, PMCID PMC2195746, lire en ligne)
- Immunobiologie, Éditions De Boeck, 2009
- (en) D. Aderka, J.M. Le et J. Vilcek, « IL-6 inhibits lipopolysaccharide-induced tumor necrosis factor production in cultured human monocytes, U937 cells, and in mice », The Journal of Immunology, vol. 143, no 11, , p. 3517-23 (résumé)
Voir aussi
Articles connexes
Liens externes
- (en) 191160
- Les anti-TNFα, un site d'information réalisé par le laboratoire Abbott France (qui commercialise des anti-TNF)
- UniProt : P01375