Plecotus macrobullaris
Oreillard montagnard, Oreillard alpin
.jpg.webp)
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Classe | Mammalia |
| Ordre | Chiroptera |
| Sous-ordre | Yangochiroptera |
| Famille | Vespertilionidae |
| Sous-famille | Vespertilioninae |
| Genre | Plecotus |
- Plecotus auritus macrobullaris Kuzjakin, 1965
- Plecotus alpinus Keifer & Veith, 2002
- Plecotus microdontus Spitzenberger, 2002
L'Oreillard montagnard (Plecotus macrobullaris), ou Oreillard alpin, est une espèce de chauves-souris de la famille des Vespertilionidae peuplant les massifs montagneux du sud-ouest et du centre du Paléarctique, des Pyrénées au Moyen-Orient. Il mesure autour de cinq centimètres du bout du museau à la base de la queue, pour un poids de 6 à 10 g et une envergure de 24 à 30 cm. Comme tous les membres du genre Plecotus, il possède d'immenses oreilles, mesurant les trois-quarts de la longueur de son corps. Il ressemble fortement à l'Oreillard roux (P. auritus), l'Oreillard gris (P. austriacus) et l'Oreillard des Balkans (P. kolombatovici), qui partagent en partie son aire de répartition, mais se distingue notamment de ces espèces par un petit triangle charnu sur la lèvre inférieure.
L'Oreillard montagnard se rencontre depuis le niveau de la mer jusqu'à 2 800 mètres d'altitude, mais s'il peut fréquenter les côtes et les plaines, il reste une espèce essentiellement montagnarde affectionnant les zones escarpées. Son régime alimentaire est presque exclusivement constitué de papillons de nuit, qu'il capture en milieux ouverts. Les colonies de mise-bas sont situées dans les combles ou les clochers d'églises, dans d'autres bâtiments, dans des crevasses rocheuses ou dans des éboulis. Elles comptent de quelques femelles à quelques dizaines de femelles, qui mettent-bas en juin-juillet et qui restent avec leur jeune jusqu'en septembre.
Sa morphologie intermédiaire entre celle des oreillards roux et gris a fait que cette espèce distincte est demeurée non reconnue comme telle jusqu'au début du XXIe siècle — avec l'essor des outils génétiques —, bien qu'un nom scientifique, publié en 1965, concernant les populations du Caucase russe ait été exhumé pour le désigner. Deux lignées mitochondriales majeures existent, avec une zone de contact dans les Alpes dinariques, et sont parfois traitées comme des sous-espèces distinctes, mais leur statut reste à être confirmé. Bien que la taille des populations et leurs tendances démographiques soient inconnues, l'Union internationale pour la conservation de la nature considère l'espèce comme de « préoccupation mineure ».
Description

Morphologie externe
L'Oreillard montagnard est un oreillard (genre Plecotus) de taille moyenne à grande, pesant entre 6 et 10 grammes, et la tête et le corps mesurant 46-55 mm de long. L'avant-bras mesure de 37,3 à 46 mm, pour une envergure comprise entre 24 et 30 centimètres. Comme tous les membres du genre Plecotus, l'Oreillard montagnard a de très grandes oreilles, qui mesurent de 34 à 38 mm, avec un tragus mesurant 5 à 6 mm de long[1]. Le pied mesure 6,6 à 9,0 mm, le pouce 5,5 à 7,9 mm, et sa griffe 0,9 à 3,6 mm. Comme chez la plupart des chauves-souris, les femelles sont en moyenne plus grandes que les mâles[2].
Les oreilles sont brun clair, le tragus aussi, avec la base de couleur chair. Les yeux sont assez grands et noirs, la face est brun foncé[2]. Le pelage est dense et long, à dominante gris clair sur le dos, avec des poils bicolores, sombres à leur base et plus clairs à leur pointe[1] - [2]. Le pelage ventral est parfois blanc pur[1], mais souvent blanchâtre avec des traces jaunâtres sur les côtés du cou et les épaules. Le patagium est brun clair, glabre[2].
Émissions sonores
Comme les autres oreillards, l'Oreillard montagnard émet des fréquences modulées, abruptes et marquées de plusieurs harmoniques. Les fréquences terminales sont de 10-25 kHz et la largeur de bande est de 17-35 kHz[3]. La première harmonique commence autour de 46 kHz et se termine vers 23 kHz avec un pic de fréquence autour de 35 kHz, et la seconde harmonique commence autour des 67 kHz et ne chevauche pas la première[4] - [5]. Ces cris durent de 1 à 7 ms[3], pouvant être aussi courts que 0,8 ms dans les environnements encombrés, ou durer jusqu'à 7,3 ms dans les environnements bien dégagés. Plus le signal est long, plus les fréquences utilisées sont basses[5]. Les caractéristiques de ces émissions recouvrent celles d'autres espèces d'oreillards coexistant avec l'Oreillard montagnard[3], et leur structure est particulièrement proche de celle de l'Oreillard gris (P austriacus), bien que le montagnard soit plus étroitement apparenté avec l'Oreillard roux (P. auritus)[5].
Caractères crâniens

La formule dentaire de l'Oreillard montagnard est la suivante (incisives, canines, prémolaires, molaires) : 2.1.2.33.1.3.3 × 2 = 36. Son crâne est semblable à celui des autres oreillards, et les espèces du genre sont difficiles à distinguer sans l'utilisation de multiples critères (voir le paragraphe Espèces similaires). La plus grande longueur du crâne est de 15,9–18,3 mm (en moyenne 17,0 mm), la longueur condylocanine (mesurée depuis la partie la plus postérieure des condyles occipitaux jusqu'au bord antérieur de la canine) de 14,1–16,1 mm (en moyenne 15,1 mm), la longueur condylobasale de 14,7–16,9 mm (en moyenne 15,8 mm), la hauteur de la boîte crânienne de 7,1–8,1 mm (en moyenne 7,6 mm), la largeur de la boîte crânienne de 7,7–9,3 mm (en moyenne 8,5 mm), la largeur interorbitale de 3,1–3,9 mm (en moyenne 3,5 mm), le diamètre de la bulle tympanique de 4,2–4,9 mm (en moyenne 4,6 mm), la largeur de la partie antérieure du palais de 3,4–4,2 mm (en moyenne 3,8 mm), la largeur du maxillaire de 5,9–6,7 mm (en moyenne 6,3 mm), la longueur de la rangée dentaire supérieure mesurée depuis la première incisive jusqu'à la troisième molaire de 6,3–7,2 mm (en moyenne 6,7 mm) et la longueur de la rangée dentaire supérieure mesurée depuis la canine jusqu'à la troisième molaire de 5,3–6,3 mm (en moyenne 5,6 mm)[6] - [7] - [8] - [2].
Espèces similaires

Dans de nombreuses zones d'Europe, l'Oreillard montagnard coexiste avec l'Oreillard roux (P. auritus), l'Oreillard gris (P. austriacus) et l'Oreillard des Balkans (P. kolombatovici), desquels il est difficile à différencier. En moyenne, l'Oreillard montagnard est légèrement plus grand que le roux, et son masque facial est moins marqué que chez le gris[9]. La coloration du tragus (couleur chair sur le tiers basal et pigmenté de gris en son extrémité), le poil très blanc sur le ventre et à pointes foncées sur le dos sont donnés comme critères complémentaires[9], mais une très forte variation dans la coloration du pelage existe et peut varier du gris au brun pour les trois espèces d'Europe de l'Ouest[10]. La longueur du pouce a également été donnée comme diagnostique pour séparer l'Oreillard montagnard — dont le pouce mesure plus de 6,2 mm — du gris ou de celui des Balkans — dont le pouce mesure moins de 6,6 mm[11] - [12]. Le meilleur caractère distinctif pour identifier l'Oreillard montagnard des deux autres espèces reste un triangle charnu présent sur la lèvre inférieure[10], d'une forme distinctive en triangle aux bords concaves, se terminant en pointe vers le menton[13].
.jpg.webp)
Chez les mâles, la forme du pénis peut aussi servir de caractère distinctif pour distinguer les oreillards européens. Chez l'Oreillard montagnard, le pénis a des bords parallèles et il se termine en pointe ; chez l'Oreillard gris et l'Oreillard des Balkans, les bords s'élargissent et l'extrémité du pénis est arrondie ; enfin, chez l'Oreillard roux, le pénis est globalement conique, les bords se rapprochant régulièrement depuis la base jusqu'à la pointe[14]. Pour les individus mâles morts, la taille et la forme du baculum (l'os pénien) est également diagnostique d'une espèce à l'autre : l'Oreillard roux a un baculum nettement plus grand et allongé que ceux du montagnard et du gris, et la base du baculum est plus massive chez l'Oreillard montagnard que chez les deux autres espèces[15].
Pour les spécimens adultes, différents caractères crâniens peuvent permettre l'identification des oreillards européens. La longueur de la rangée dentaire et de la bulle tympanique sont parfois données comme diagnostiques[16] - [17], mais un grand recouvrement existe entre l'Oreillard roux (crâne aux petites dimensions), l'Oreillard montagnard (intermédiaire) et l'Oreillard gris (grandes dimensions)[13]. D'après une étude réalisée en Croatie, le crâne de l'Oreillard des Balkans est encore plus difficile à différencier de celui de l'Oreillard montagnard, mais la largeur inter-orbitale figurerait parmi les meilleurs caractères séparant ces deux espèces, mesurant en moyenne 3,30 mm chez l'Oreillard des Balkans et 3,57 mm chez le montagnard[18]. Sur les crânes intacts, une combinaison de 13 caractères crâniens permet de distinguer les quatre oreillards d'Europe continentale (le montagnard, le roux, le gris et celui des Balkans)[18].
Toutes ces espèces d'oreillards se distinguent sans ambiguïté sur la base de marqueurs mitochondriaux, que ce soit le cytochrome b (cyt-b) et la région de contrôle du mitogénome (CR)[19] - [20], la première sous-unité de la cytochrome c oxydase (COI)[21] et l'ARN ribosomique 16S[13]. Pour permettre une identification moléculaire par simple PCR sans besoin de séquençage, plusieurs amorces ont été mises au point pour amplifier des fragments du 16S de tailles différentes et diagnostiques de P. macrobullaris, P. austriacus et deux lignées de P. auritus[13].
Écologie et comportement
Alimentation

En Europe de l'Ouest, le régime alimentaire de l'Oreillard montagnard est essentiellement constitué de papillons de nuit (entre 87 et 98 %), notamment de la famille des Noctuidae. Il compte aussi des diptères et coléoptères[23] - [24] - [22]. En Turquie, la présence de restes d'hémiptères et d'orthoptères dans les crottes a été rapportée[25]. En plus des lépidoptères, diptères et coléoptères, les études par métabarcoding de crottes ont livré en petites proportions des araignées, hémiptères, hyménoptères et névroptères, ainsi que quelques blattes, trichoptères, dermaptères, mécoptères, orthoptères, psoques et raphidioptères. La détection génétique de mouches tachinaires (diptères de la famille des Tachinidae) provient potentiellement de la consommation par les chauves-souris de chenilles de noctuelles infectées par les larves de ces insectes parasitoïdes[22]. De nombreux papillons de nuit disposant d'organes tympaniques, et détectant donc les cris d'écholocalisation de la plupart des chauves-souris, ont été retrouvés dans le régime alimentaire de l'Oreillard montagnard[26]. En effet, les oreillards sont des chauves-souris dites « chuchoteuses » ou « murmurantes », connues pour émettre en chasse des cris de très faible intensité leur permettant de consommer les papillons tympanés qui les éviteraient sinon[27] - [28]. À l'inverse, ils utilisent leur vision et l'écoute passive de leurs proies pour les détecter et les capturer[29] - [30]. Par rapport aux autres oreillards partageant son aire de distribution, l'Oreillard montagnard chasse dans des milieux ouverts ou montagneux (prairies, pelouses, pentes, éboulis et habitats rocheux). Parmi les proies de son régime alimentaire reflétant bien ses préférences écologiques distinctes, on peut notamment citer les noctuelles Euxoa aquilina et Bryophila domestica, ou les géomètres Gnophos furvata, Hemistola chrysoprasaria, Horisme radicaria, Nychiodes obscuraria et Yezognophos dilucidaria[22].
Déplacements
Comme les autres oreillards, l'Oreillard montagnard se déplace globalement peu pour s'alimenter, les terrains de chasse se trouvant généralement à moins de 4 kilomètres du gîte[23] - [31] - [26]. Cependant, des déplacements de 7 km dans une nuit de chasse dans les Pyrénées[32], ou de 9 km en Corse ont été rapportés[33]. Les terrains de chasse couvrent plusieurs centaines d'hectares, jusqu'à plus de 4 000 ha pour certains individus[34]. En Arménie, un déplacement de 79 km initialement attribué à l'Oreillard gris avant la découverte de l'Oreillard montagnard pourrait bien concerner cette dernière espèce[34].
Reproduction

Bien que son proche cousin l'Oreillard roux (P. auritus) soit l'espèce la plus commune sur les sites de regroupements automnaux (aussi connus sous le nom d'essaimage ou de « swarming ») où ont probablement lieu le choix des partenaires et l'accouplement, ce comportement n'est pas connu chez l'Oreillard montagnard[35]. L'essentiel des colonies de mise-bas connues sont situées dans des bâtiments, dans les charpentes couvertes de pierre, de zinc, d'ardoises ou de bois[36]. Les suivis télémétriques d'individus dans les Pyrénées ont cependant montré que l'espèce pouvait également établir ses sites de parturition en milieu rocheux, dans les crevasses comme dans les éboulis, en haute-montagne[32]. En milieu bâti, les Oreillards montagnards sont très fidèles à leur site de mise-bas, qu'ils réinvestissent d'une année sur l'autre. La fidélité au gîte est un peu plus faible concernant les colonies établies dans des crevasses, et encore plus faible dans les éboulis[32]. Le sexe et le statut reproducteur affecte aussi la fidélité des individus à leurs gîtes, les femelles allaitantes ne changeant quasiment pas de gîte, les femelles nullipares étant un peu plus mobiles et les mâles changeant de gîte quasiment tous les jours[32]. Les colonies de mise-bas de l'Oreillard montagnard comptent généralement 5 à 25 femelles[10], mais des colonies comptant une centaine d'individus, jeunes compris, existent notamment en France[37] - [38]. Ces colonies comptent des femelles nullipares, et la présence occasionnelle de mâles a également été signalée[32]. La mise-bas a lieu entre juin et juillet et les jeunes restent au gîte avec les adultes jusqu'en septembre[37].
Hibernation
Peu de données sont connues sur l'hibernation de l'Oreillard montagnard. Seules deux mentions, d'individus hibernant dans des grottes à plus de 2 000 m d'altitude dans les Pyrénées[39] et dans les Alpes slovènes[40], ont rétrospectivement été attribuées à cette espèce après sa découverte[41].
Interaction avec les autres chiroptères
L'Oreillard montagnard forme des colonies monospécifiques[41], mais en Europe centrale l'espèce a été notée pouvant partager un même bâtiment avec l'Oreillard roux (P. auritus)[10] ou l'Oreillard des Balkans (P. kolombatovici)[12]. En Iran, l'espèce peut également partager une même grotte avec le Rhinolophe de Blasius (Rhinolophus blasii), le Minioptère pâle (Miniopterus pallidus), le Petit Murin (Myotis blythii) ou le Murin de Schaub (M. schaubi)[42].

Certains auteurs ont suggéré que l'Oreillard montagnard et l'Oreillard gris (P. austriacus) répondent au principe d'exclusion compétitive (ou principe de Gause), c'est-à-dire qu'ils partagereaint des écologies similaires empêchant leur coexistence durable dans le temps[43] - [26]. Plusieurs arguments sont avancés en ce sens. Ces deux espèces ont des cris d'écholocalisation et un comportement de vol très proches[4], chassent en milieux ouverts le même type de proies, des papillons de nuit. À l'échelle régionale, ces deux oreillards présentent parfois des répartitions parapatriques (c'est-à-dire adjacentes et sans superposition), comme en Suisse où l'Oreillard montagnard se trouve essentiellement dans les Alpes au sud du pays, et l'Oreillard gris est cantonné au nord du pays[43] - [13], ou en Croatie où le montagnard occupe les zones karstiques et le gris les plaines[12]. Dans les Pyrénées, une ségrégation altitudinale est observée, l'Oreillard montagnard se trouvant à haute altitude et le gris dans les zones plus basses[44]. Si cette hypothèse peut localement sembler valable, elle est cependant peu cohérente avec d'autres éléments. La distribution de l'Oreillard montagnard est fortement liée aux massifs montagneux sur l'ensemble de son aire de répartition, et notamment au Proche-Orient où l'Oreillard gris n'est pourtant pas présent. Pourtant, selon le principe de Gause on devrait s'attendre à ce que l'Oreillard montagnard occupe une niche bien plus large dans ces zones en l'absence de la compétition interspécifique avec l'Oreillard gris[45]. En outre, de nombreuses zones de sympatrie des deux espèces existent, comme dans les Pyrénées, dans les pré-alpes françaises[46], sur le canton de Genève[47] - [13] et dans les Alpes dinariques[12], mettant également à mal le principe de Gause[45]. L'étude du régime alimentaire par métabarcoding des déjections a en outre montré que les ressemblances superficielles du régime alimentaire des oreillards s'estompaient sitôt que les proies étaient identifiées plus finement. Oreillard montagnard et oreillard gris tendent à moins fréquenter les forêts que l'Oreillard roux (P. auritus), mais l'Oreillard montagnard se distingue des deux autres par la présence de proies de milieux très ouverts, trouvées en altitude, dans les prairies et pelouses, même quand les gîtes hébergeant les colonies sont situés en plaine[22].
Prédateurs et parasites
Des restes d'oreillards retrouvés dans des pelotes de réjection d'Effraie des clochers (Tyto alba), en Crète[48], et initialement attribués à l'Oreillard gris concernent plus vraisemblablement l'Oreillard montagnard[49].
Comme toutes les espèces de chauves-souris, l'Oreillard montagnard peut être l'hôte de différents groupes de parasites, mais peu de données ont été publiées à ce sujet depuis sa description. L'acarien Spinturnix plecotina — de la famille des Spinturnicidae et qui parasite d'autres vespertilionidés et notamment des oreillards — a été identifié chez P. macrobullaris en Albanie et en Turquie[50] - [51], ainsi que la protonymphe d'un acarien du genre Ornithonyssus, de la famille des Macronyssidae[52]. La tique Dermacentor marginatus a été signalée chez un Oreillard montagnard capturé en Turquie, mais correspond probablement à un cas accidentel[53].
Répartition et habitat
Distribution géographique
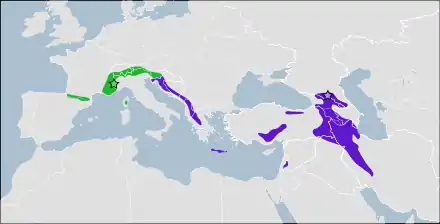
- Plecotus macrobullaris alpinus
- Plecotus macrobullaris macrobullaris
L'Oreillard montagnard est la seule espèce de chauves-souris du Paléarctique dont la répartition est restreinte aux massifs montagneux[55]. Il vit dans les chaînes de montagnes dans le sud-ouest et le centre du Paléarctique, ainsi que dans leurs piémonts et leurs proches environs[44].
À l'ouest, sa répartition commence dans les Pyrénées, entre l'Espagne et la France[56] - [44], puis couvre l'ensemble des Alpes : dans le sud-est de la France[57], tout le nord de l'Italie[58] - [59], au Liechtenstein et en Suisse[60] - [43] - [13], en Autriche[20] et en Slovénie[55]. L'Oreillard montagnard est également présent en Corse, bien que sa répartition exacte soit méconnue du fait des difficultés à le différencier de l'Oreillard gris qui se trouve également sur l'île[33]. Dans la péninsule des Balkans, la répartition de l'espèce est limitée aux Alpes dinariques, au Pinde et au mont Olympe[55] - [61], et concerne les pays suivants : Croatie[12], Bosnie-Herzégovine, Serbie[62], Monténégro[63], Albanie[64], Macédoine[65] et Grèce[44]. Dans ce dernier pays, l'Oreillard montagnard est également présent sur l'île de Crète, où il a été capturé dans la plupart des zones montagneuses[49]. Au Proche-Orient, l'Oreillard montagnard peuple les monts Kaçkar du nord de l'Anatolie, et les monts Taurus au sud de cette région. Plus à l'ouest, il vit dans le Grand Caucase, en Géorgie et en Russie[66] - [44], et dans le Petit Caucase, en Arménie et en Azerbaïdjan[66]. Plus au sud on le trouve dans l'Elbourz et les monts Zagros, en Iran[42]. Enfin, une population est connue de l'Anti-Liban, en Syrie[67].
Distribution altitudinale
L'Oreillard montagnard est une espèce essentiellement montagnarde, mais sa distribution altitudinale peut varier fortement d'une région à une autre, et globalement l'espèce est distribuée depuis le niveau de la mer jusqu'à près de 2 800 mètres d'altitude[44]. Dans les Pyrénées et les monts Zagros, l'essentiel des signalements sont situés à plus de 1 500 m d'altitude[55]. Dans les régions montagneuses très escarpées, l'Oreillard montagnard vit aussi bien à très haute altitude que près des côtes ou en plaine. En Corse par exemple, l'espèce semble distribuée sur tout le gradient altitudinal, de 0 à plus de 2 000 m[33] - [35]. Il en va de même dans l'arc Alpin, ou des nurseries ont été trouvées à plus de 2 000 m d'altitude[23] mais où des colonies sont connues en basses altitudes comme dans les calanques marseillaises[47], dans les plaines slovènes entre 300 et 600 m[68], dans le bassin genevois à 430 m d'altitude[47], ou en Croatie où l'essentiel des colonies sont situées à moins de 800 m d'altitude[69].
Habitat
.jpg.webp)
Une étude de modélisation de la niche écologique de l'Oreillard montagnard a montré que sa répartition semble avant tout contrainte par des facteurs topographiques plutôt que climatiques : l'espèce n'est pas principalement cantonnée aux zones montagneuses pour leur climat froid, mais pour leur topographie particulière. Ainsi, comparée à celles de tous les mammifères et oiseaux européens, la répartition géographique de l'Oreillard montagnard se rapproche beaucoup de celle d'oiseaux peuplant des zones très escarpées, comme la Niverolle alpine (Montifringilla nivalis), le Chocard à bec jaune (Pyrrhocorax graculus), le Tichodrome échelette (Tichodroma muraria), et l'Accenteur alpin (Prunella collaris). La cinquième espèce à la distribution la plus similaire est un mammifère, le Campagnol des neiges (Chionomys nivalis)[45]. L'importance relativement faible du climat se retrouve d'ailleurs dans la diversité de milieux peuplés par l'Oreillard montagnard, des montagnes extrêmement froides et pluvieuses des Alpes, aux massifs très secs et chauds du Proche-Orient[26]. Ces terrains escarpés procurent possiblement à cet oreillard les zones rocheuses et ouvertes qui sont propices à sa technique de chasse et à l'établissement de ses colonies de mise-bas[45]. L'Oreillard montagnard est probablement à l'origine une espèce liée aux milieux karstiques, offrant de nombreuses crevasses, éboulis et grottes. Dans le sud-est de son aire de répartition, l'espèce est communément trouvée dans des grottes[49], et elle est également trouvée dans les environnements calcaires dans les Pyrénées[44] et les Balkans[12].
Dans les Pyrénées, le suivi télémétrique d'individus capturés en haute altitude a montré que l'Oreillard montagnard utilisait essentiellement des crevasses rocheuses comme gîte, et des éboulis pour établir ses colonies de mise-bas[32]. À l'inverse, dans les Alpes les sites de mise-bas connus de l'espèce se trouvent systématiquement dans les bâtiments, et l'espèce est l'une des plus communes dans les combles et clochers d'églises[43]. Cette différence est peut-être expliquée par le climat globalement plus froid dans les Alpes (de 2 °C) qui pousse les chauves-souris à privilégier la relative chaleur des bâtiments, ou par un biais de prospection favorisant la découverte des colonies dans le milieu bâti[26]. Les cavités arboricoles ne semblent pas utilisées pour établir les colonies de mise-bas, même quand elles sont disponibles[33], mais la mention d'un individu mâle dans une telle cavité est connue[26]. Pour les colonies situées dans les bâtiments, l'environnement immédiat tend à être plutôt forestier dans les Alpes[43], et elles sont entourées par ou proches de forêts décidues dans les Pyrénées[32], mais les colonies établies dans les zones rocheuses ont tendance à être situées près de prairies ou de forêts ouvertes[32] - [26].
De nombreux indices directs et indirects laissent penser que l'Oreillard montagnard chasse dans les milieux ouverts, au moins dans les Pyrénées[26] : il a été capturé chassant à proximité du sol dans les pelouses d'altitude, bien au-dessus de la limite des arbres[44], de nombreuses colonies sont situées loin de tout environnement forestier[32] et la caractérisation précise de son régime alimentaire a livré des espèces de papillons de nuit typiques des milieux ouverts[24]. En Italie, des suivis télémétriques ont également montré que l'Oreillard montagnard évitait les zones boisées[31]. À l'inverse, en Croatie et dans les Alpes suisses, l'Oreillard montagnard a été décrit comme chassant en forêt décidue[69] ou privilégiant le couvert végétal structuré verticalement[43].
Systématique
Taxinomie
Le zoologiste russe Alexandre Kouziakine (dont le nom est transcrit en « Kuzjakin » pour la citation d'auteur zoologique) décrit Plecotus auritus macrobullaris en 1965[70], qu'il considère donc être une sous-espèce de l'Oreillard roux (P. auritus), d'après un matériel type provenant du Caucase, près de Vladikavkaz (Ossétie du Nord, Russie). La dénomination subspécifique, « macrobullaris », signifie « à grosses bulles », en référence à la taille des bulles tympaniques plus grandes que chez la sous-espèce nominale de l'Oreillard roux (P. auritus auritus).
À cette époque, dans le reste de l'Europe seul l'Oreillard roux typique (P. a. auritus) et l'Oreillard gris (P. austriacus) sont reconnus, et ce depuis 1960, année où l'on se met à distinguer P. austriacus de P. auritus. Cette situation prévaut jusqu'au début du XXIe siècle, quand deux lignées génétiques très divergentes sont identifiées chez l'Oreillard roux en Europe de l'Ouest. L'un des clades est attribué au « vrai » P. auritus et l'autre est assigné, mais à tort, à l'Oreillard des Balkans (P. kolombatovici), un taxon décrit de Croatie en 1980 comme sous-espèce de l'Oreillard gris[71] - [7]. Dans la foulée, deux équipes de chiroptérologues allemands décrivent indépendamment cette seconde lignée comme nouvelle espèce, sous les noms de Plecotus alpinus Kiefer & Veith, 2002, à partir d'un spécimen de Ristolas (Hautes-Alpes, France)[6], et Plecotus microdontus Spitzenberger, 2002, à partir d'un spécimen de Lienz (Tyrol, Autriche)[11]. La description des deux noms paraît à quelques semaines d'intervalle, Plecotus alpinus ayant de peu la priorité[72].
En 2003, l'examen de matériel du Caucase et d'Asie mineure à l'aide d'analyses morphologiques et génétiques montrent que l'espèce nouvellement décrite des Alpes (sous les noms d'alpinus et de microdontus) est conspécifique avec macrobullaris décrit du Caucase. Ce dernier taxon est alors élevé au rang d'espèce, et Plecotus macrobullaris Kuzjakin, 1965 devient le nom valide de l'espèce selon le principe de priorité du code international de nomenclature zoologique[7]. Malgré les fortes ressemblances morphologiques et acoustiques entre les Oreillards montagnard, roux et gris, les études de génétique des populations conduites sur des zones de contact entre les espèces n'ont pas trouvé de traces d'hybridation récente ou ancienne parmi les trois espèces[35].
Sous-espèces
En raison de divergences génétiques marquées entre différentes populations de l'Oreillard montagnard, certains auteurs proposent de distinguer deux sous-espèces[8] - [73] :
- Plecotus macrobullaris alpinus Kiefer & Veith, 2002, dans les Pyrénées et les Alpes, ainsi qu'en Corse ;
- Plecotus macrobullaris macrobullaris Kuzjakin, 1965, dans les Alpes dinariques et les Balkans, en Crète et à l'est jusqu'au Caucase, monts Taurus et monts Zagros.
La zone de contact entre ces deux groupes se situe dans le nord des Alpes dinariques où les deux lignées coexistent[74] ; par exemple, un spécimen de la lignée orientale a été identifié dans le Frioul[20]. Les deux lignées présentent une dissimilarité d'environ 3,4 % de leur mitogénome, correspondant à un temps de divergence de plus d'un million d'années, et à une différence comparable à celle séparant le Zèbre de Grévy (Equus grevyi) du Zèbre de Burchell (Equus quagga), ou plus importante que celle séparant le Putois (Mustela putorius) du Vison de Sibérie (Mustela sibirica)[74]. Ces différences n'ont cependant été caractérisées qu'à partir de l'ADN mitochondrial et mériteraient d'être confirmées par l'étude de marqueurs nucléaires[75]. En outre, les analyses crâniométriques effectuées à ce jour ne retrouvent pas la structure est-ouest mise en évidence par la génétique, mais rapprochent d'une part les populations des Alpes (lignée mitochondriale de l'ouest) et celles du Caucase (lignée de l'est), et d'autre part celles des Balkans, de Turquie et de Syrie[20] - [75]. Des analyses de génétique des populations conduites en 2018 ont montré une légère différentiation nucléaire entre les populations d'Oreillard montagnard de Corse et celles des Alpes iséroises et suisses, mais l'étude en question n'incluait pas d'individus de la lignée mitochondriale de l'est[35].
Phylogénie
| Phylogénie partielle du genre Plecotus selon Spitzenberger et al. (2006)[20] et Kruskop et al. (2021)[76] : | |
|
Dans l'arrangement du genre Plecotus proposé par Spitzenberger et al. (2006), l'Oreillard montagnard appartient au groupe d'espèces de Plecotus auritus. Au sein de ce groupe, l'Oreillard du Turkménistan (P. turkmenicus) semble avoir divergé le premier de toutes les autres espèces. Ces dernières se répartissent en deux clades bien soutenus. Un premier, asiatique, regroupe l'Oreillard du Japon (P. sacrimontis), l'Oreillard de Kozlov (P. kozlovi), l'Oreillard d'Ognev (P. ognevi) et l'Oreillard de Strelkov (P. strelkovi). Le second comprend l'Oreillard roux (P. auritus), l'Oreillard sarde (P. sardus), l'Oreillard montagnard (P. macrobullaris) et l'Oreillard de l'Himalaya (P. wardi). Dans la plupart des analyses, l'Oreillard montagnard est d'abord rapproché de l'Oreillard sarde, l'Oreillard roux formant leur lignée sœur et l'Oreillard de l'Himalaya étant frère des trois autres, mais les nœuds sont statistiquement peu soutenus et les relations de parenté exactes entre les espèces doivent encore être précisées[20].
Menaces et conservation
L'Union internationale pour la conservation de la nature considère l'Oreillard montagnard comme de « préoccupation mineure »[54], mais du fait de sa découverte récente le statut de l'espèce est toujours incertain[77]. Dans la Directive « Habitats, Faune, Flore », l'espèce est classée en annexe IV, avec un danger actuellement difficile à estimer, mais elle est vraisemblablement menacée par la destruction de ses gîtes lors de la restauration de bâtiments[78]. L'espèce semble abondante dans plusieurs milieux montagneux, des Pyrénées aux Alpes dinariques en passant par les Alpes[77], mais les effectifs exacts de même que leurs tendances démographiques sont toujours inconnus[79]. Les analyses de marqueurs nucléaires ayant mis en évidence des différences marquées entre les populations de Corse et celle des Alpes occidentales, celles-ci forment des « unités évolutives significatives » distinctes et méritent possiblement des mesures de conservation propres[35]. Comme l'essentiel des colonies connues dans les Alpes se situent en milieu bâti[43], et notamment dans les greniers et clochers d'églises qui offrent des abris chauds dans un climat très froid, la conservation et la gestion de ces sites de mise-bas pourraient être localement essentielles pour la préservation de l'espèce[77]. La protection de l'Oreillard montagnard passe également par celle de ses terrains de chasse comprenant des habitats diversifiés (prairies, friches, haies vives, lisières…) qui lui fournissent les lépidoptères constituant l'essentiel de son alimentation[47].
Annexes
Bibliographie
- (fr) Laurent Arthur et Michèle Lemaire, Les Chauves-souris de France, Belgique, Luxembourg et Suisse, Mèze et Paris, Biotope et Muséum national d'histoire naturelle, , 2e éd., 544 p. (ISBN 978-2-36662-139-6), « L'Oreillard montagnard. Plecotus macrobullaris Kuzjakin, 1965 », p. 439-442
- (fr) Christian Dietz et Andreas Kiefer, Chauves-souris d'Europe : Connaître, identifier, protéger, Paris, Delachaux et Niestlé, , 399 p. (ISBN 978-2-603-02158-3), « Oreillard montagnard. Plecotus macrobullaris (Kuzjakin, 1965) », p. 360-363
- (en) Antton Alberdi et Ostaizka Aizpurua, « Plecotus macrobullaris (Chiroptera: Vespertilionidae) », Mammalian Species, ASM et OUP, vol. 50, no 958, , p. 26–33 (ISSN 0076-3519 et 1545-1410, OCLC 46381503, DOI 10.1093/MSPECIES/SEY003)
Références taxinomiques
- (en) Référence Mammal Species of the World (3e éd., 2005) : Plecotus austriacus macrobullaris Kuzyakin, 1965
- (en) Référence BioLib : Plecotus macrobullaris Kuzjakin, 1965 (consulté le )
- (en) Référence Catalogue of Life : Plecotus macrobullaris Kuzyakin, 1965 (consulté le )
- (fr+en) Référence ITIS : Plecotus macrobullaris Kuzyakin, 1965 (consulté le )
Liens externes
- (en) Référence UICN : espèce Plecotus macrobullaris Kuzjakin, 1965 (consulté le )
- (en) Référence Fauna Europaea : Plecotus macrobullaris Kuzjakin, 1965 (consulté le )
- (fr) Référence INPN : Plecotus macrobullaris Kuzjakin, 1965 (TAXREF) (consulté le )
- (en) Référence NCBI : Plecotus macrobullaris (taxons inclus) (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023
- (en) Plecotus macrobullaris Kuzjakin, 1965, sur le portail GBIF
Notes et références
- Arthur & Lemaire (2015), p. 439-440, « Caractères distinctifs »
- Alberdi & Aizpurua (2018), p. 28, « General characters »
- Arthur & Lemaire (2015), p. 440, « Émissions sonores »
- (en) Susanne Dietrich, Diana P. Szameitat, Andreas Kiefer, Hans-Ulrich Schnitzler et Annette Denzinger, « Echolocation signals of the plecotine bat, Plecotus macrobullaris Kuzyakin, 1965 », Acta Chiropterologica, Musée et institut de zoologie de l'Académie polonaise des sciences (d), vol. 8, no 2, , p. 465-475 (ISSN 1508-1109 et 1733-5329, DOI 10.3161/1733-5329(2006)8[465:ESOTPB]2.0.CO;2)
- Alberdi & Aizpurua (2018), p. 29, « Form and Function »
- (en) Andreas Kiefer et Michael Veith, « A new species of long-eared bat from Europe (Chiroptera: Vespertilionidae) », Myotis: international journal of bat research, Musée Alexander Koenig, vol. 39, , p. 5-16 (ISSN 0580-3896, lire en ligne)
- (en) Friederike Spitzenberger, Petr P. Strelkov et Elisabeth Haring, « Morphology and mitochondrial DNA sequences show that Plecotus alpinus Kiefer & Veith, 2002 and Plecotus microdontus Spitzenberger, 2002 are synonyms of Plecotus macrobullaris Kuzjakin, 1965 », Natura Croatica, Musée d'histoire naturelle de Croatie (d), vol. 12, no 2, , p. 39-53 (ISSN 1330-0520 et 1848-7386, OCLC 28837675, lire en ligne)
- (en) Petr Benda, Andreas Kiefer, Vladimír Hanák et Michael Veith, « Systematic status of African populations of long-eared bats, genus Plecotus (Mammalia: Chiroptera) », Folia Zoologica, Institut de Biologie des vertébrés de l'Académie tchèque des sciences (d), vol. 53, , p. 1-47 (ISSN 0139-7893 et 1210-9681)
- Arthur & Lemaire (2015), p. 440, « Confusions possibles et variations géographiques »
- (en) Sohrab Ashrafi, Fabio Bontadina, Andreas Kiefer, Igor Pavlinić et Raphaël Arlettaz, « Multiple morphological characters needed for field identification of cryptic long‐eared bat species around the Swiss Alps », Journal of Zoology, Wiley-Blackwell, vol. 281, no 4, , p. 241-248 (ISSN 1469-7998, 0022-5460 et 0952-8369, OCLC 15264754, DOI 10.1111/J.1469-7998.2010.00697.X)
- (en) Friederike Spitzenberger, Elisabeth Haring et Nikola Tvrtković, « Plecotus microdontus (Mammalia, Vespertilionidae), a new bat species from Austria », Natura Croatica, Musée d'histoire naturelle de Croatie (d), vol. 11, , p. 1-18 (ISSN 1330-0520 et 1848-7386, OCLC 28837675, lire en ligne)
- (en) Nikola Tvrtković, Igor Pavlinić et Elisabeth Haring, « Four species of long-eared bats (Plecotus, Geoffroy, 1818; Mammalia, Vespertilionidae) in Croatia: field identification and distribution », Folia Zoologica, Institut de Biologie des vertébrés de l'Académie tchèque des sciences (d), vol. 54, nos 1-2, , p. 75-88 (ISSN 0139-7893 et 1210-9681)
- (en) Tommy Andriollo et Manuel Ruedi, « Novel molecular tools to identify Plecotus bats in sympatry and a review of their distribution in Switzerland », Revue suisse de Zoologie, MHNG, vol. 125, no 1, , p. 61-72 (ISSN 0035-418X, DOI 10.5281/ZENODO.1196013)
- (de) Christian Dietz, Otto von Helversen et Dietmar Nill, Handbuch der Fledermäuse Europas und Nordwestafrikas : Biologie, Kennzeichen, Gefährdung, Stuttgart, Kosmos, , 399 p. (ISBN 3-440-09693-9)
- (en) Anna Nele Herdina, Barbara Herzig-Straschil, Helge Hilgers, Brian Metscher et Hanns Plenk, « Histomorphology of the penis bone (baculum) in the gray long-eared bat Plecotus austriacus (Chiroptera, Vespertilionidae). », Anatomical Record, Hoboken, Wiley, vol. 293, no 7, , p. 1248-1258 (ISSN 1932-8486 et 1932-8494, OCLC 70853202, PMID 20583269, DOI 10.1002/AR.21148)
- (en) Petr Benda et Teodora Ivanova, « Long-eared bats, genus Plecotus (Mammalia: Chiroptera) in Bulgaria: a revision of systematic and distributional status », Časopis Národního Muzea. Rada Přírodovědna, vol. 172, , p. 157-172 (ISSN 1214-0635)
- (fr) Michel Blant, Benoît Magnin et Manuel Ruedi, « Clé des crânes (Chiroptères) », dans Paul Marchesi, Michel Blant & Simon Capt, Mammifères de Suisse - Clés de détermination, vol. 21, Neuchâtel, Fauna Helvetica, CSCF & SSBF, , p. 132-145
- (en) Igor Pavlinić et Maja Đaković, « Identification of four Plecotus species (Chiroptera, Vespertilionidae) in Croatia based on cranial characters », Mammalia, De Gruyter, vol. 80, no 4, , p. 385-394 (ISSN 0025-1461 et 1864-1547, DOI 10.1515/MAMMALIA-2014-0031)
- (en) Javier Juste, Carlos Ibáñez, Joaquin Muñoz, Domingo Trujillo, Petr Benda, Ahmet Karataş et Manuel Ruedi, « Mitochondrial phylogeography of the long-eared bats (Plecotus) in the Mediterranean Palaearctic and Atlantic Islands », Molecular Phylogenetics and Evolution, Academic Press et Elsevier, vol. 31, no 3, , p. 1114-1126 (ISSN 1055-7903 et 1095-9513, PMID 15120404, DOI 10.1016/J.YMPEV.2003.10.005)
- (en) Friederike Spitzenberger, Petr P. Strelkov, Hans Winkler et Elisabeth Haring, « A preliminary revision of the genus Plecotus (Chiroptera, Vespertilionidae) based on genetic and morphological results », Zoologica Scripta, Wiley-Blackwell, vol. 35, no 3, , p. 187-230 (ISSN 0300-3256 et 1463-6409, DOI 10.1111/J.1463-6409.2006.00224.X)
- Alberdi & Aizpurua (2018), p. 27, « Diagnosis »
- (en) Tommy Andriollo, Johan R. Michaux et Manuel Ruedi, « Food for everyone: Differential feeding habits of cryptic bat species inferred from DNA metabarcoding », Molecular Ecology, Wiley-Blackwell, vol. 30, no 18, , p. 4584-4600 (ISSN 0962-1083 et 1365-294X, OCLC 39265322, DOI 10.1111/MEC.16073)
- (en) Sohrab Ashrafi, Andres Beck, Marianne Rutishauser, Raphaël Arlettaz et Fabio Bontadina, « Trophic niche partitioning of cryptic species of long-eared bats in Switzerland: implications for conservation », European Journal of Wildlife Research, Springer Science+Business Media, vol. 57, no 4, , p. 843-849 (ISSN 1612-4642, 1439-0574 et 0044-2887, DOI 10.1007/S10344-011-0496-Z)
- (en) Antton Alberdi, Inazio Garin, Ostaizka Aizpurua et Joxerra Aihartza, « The foraging ecology of the mountain long-eared bat Plecotus macrobullaris revealed with DNA mini-barcodes », PLOS One, PLoS, vol. 7, no 4, , e35692 (ISSN 1932-6203, OCLC 228234657, PMID 22545129, PMCID 3335802, DOI 10.1371/JOURNAL.PONE.0035692)
- (en) John O. Whitaker Jr. et Ahmet Karataş, « Food and feeding habits of some bats from Turkey », Acta Chiropterologica, Musée et institut de zoologie de l'Académie polonaise des sciences (d), vol. 11, no 2, , p. 393-403 (ISSN 1508-1109 et 1733-5329, DOI 10.3161/150811009X485611)
- Alberdi & Aizpurua (2018), p. 29-31, « Ecology »
- (en) Gareth Jones et Jens Rydell, « Attack and defence: interactions between echolocating bats and their insect prey », dans Thomas H. Kunz et M. Brock Fenton, Bat Ecology, Chicago, University of Chicago Press, , 779 p., p. 301-345
- (en) Jon Russ, British bat calls : A guide to species identification, Exeter, , 204 p. (ISBN 978-1-907807-25-1)
- (en) M. Elizabeth Anderson et Paul Adrian Racey, « Feeding behaviour of captive brown long-eared bats Plecotus auritus », Animal Behaviour, Elsevier, vol. 42, no 3, , p. 489-493 (ISSN 0003-3472 et 1095-8282, OCLC 04699737, DOI 10.1016/S0003-3472(05)80048-X)
- (en) Johan Eklöf et Gareth Jones, « Use of vision in prey detection by brown long-eared bats, Plecotus auritus », Animal Behaviour, Elsevier, vol. 66, no 5, , p. 949-953 (ISSN 0003-3472 et 1095-8282, OCLC 04699737, DOI 10.1006/ANBE.2003.2272)
- (en) Damiano G. Preatoni, Martina Spada, Lucas A. Wauters, Guido Tosi et Adriano Martinoli, « Habitat use in the female Alpine long-eared bat (Plecotus macrobullaris): does breeding make the difference? », Acta Chiropterologica, Musée et institut de zoologie de l'Académie polonaise des sciences (d), vol. 13, no 2, , p. 355-364 (ISSN 1508-1109 et 1733-5329, DOI 10.3161/150811011X624820)
- (en) Antton Alberdi, Joxerra Aihartza, Ostaizka Aizpurua, Egoitz Salsamendi, R. Mark Brigham et Inazio Garin, « Living above the treeline: roosting ecology of the alpine bat Plecotus macrobullaris », European Journal of Wildlife Research, Springer Science+Business Media, vol. 61, no 1, , p. 17-25 (ISSN 1612-4642, 1439-0574 et 0044-2887, DOI 10.1007/S10344-014-0862-8)
- (fr) Jean-Yves Courtois, Delphine Rist et Grégory Beuneux, Les chauves-souris de Corse, Albiana, , 166 p. (ISBN 978-2-84698-377-8), p. 141-144, « L'Oreillard gris (Plecotus austriacus), l'Oreillard montagnard (P. macrobullaris) »
- Dietz & Kiefer (2015), p. 363, « Utilisation de l'espace »
- (en) Tommy Andriollo, Sohrab Ashrafi, Raphaël Arlettaz et Manuel Ruedi, « Porous barriers? Assessment of gene flow within and among sympatric long‐eared bat species », Ecology and Evolution, Wiley-Blackwell et Wiley, vol. 8, no 24, , p. 12841-12854 (ISSN 2045-7758, OCLC 757379582, PMID 30619587, PMCID 6309003, DOI 10.1002/ECE3.4714)
- Arthur & Lemaire (2015), p. 442, « Estivage »
- Arthur & Lemaire (2015), p. 442, « Reproduction »
- Dietz & Kiefer (2015), p. 363, « Comportement et régime alimentaire »
- (eu) Joxerra Aihartza, « Ipar-belarrihandia, bitxia bezain ezezaguna », Elhuyar, vol. 60, , p. 25-27
- (en) Boris Kryštufek, « Distribution of bats in Slovenia (Yugoslavia) », dans Vladimír Hanák, Ivan Horáček et Jiří Gaisler, European bat research 1987, Prague, Charles University Press, , p. 399-401
- Alberdi & Aizpurua (2018), p. 31, « Behavior »
- (en) Petr Benda, Kaveh Faizolâhi, Michal Andreas, Ján Obuch, Antonín Reiter, Martin Ševčík, Marcel Uhrin, Peter Vallo et Sohrab Ashrafi, « Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 10. Bat fauna of Iran », Acta Societatis Zoologicae Bohemicae, Czech Zoological Society (d), vol. 76, , p. 163-582 (ISSN 1211-376X)
- (en) Marianne D. Rutishauser, Fabio Bontadina, Veronika Braunisch, Sohrab Ashrafi et Raphaël Arlettaz, « The challenge posed by newly discovered cryptic species: disentangling the environmental niches of long‐eared bats », Diversity and Distributions, Wiley-Blackwell et Wiley, vol. 18, no 11, , p. 1107-1119 (ISSN 1366-9516 et 1472-4642, OCLC 39848596, DOI 10.1111/J.1472-4642.2012.00904.X)
- (en) Antton Alberdi, Inazio Garin, Ostaizka Aizpurua et José Ramón Ahiartza Azurtza, « Review on the geographic and elevational distribution of the mountain long-eared bat Plecotus macrobullaris, completed by utilising a specific mist-netting technique », Acta Chiropterologica, Musée et institut de zoologie de l'Académie polonaise des sciences (d), vol. 15, no 2, , p. 451-461 (ISSN 1508-1109 et 1733-5329, DOI 10.3161/150811013X679071)
- (en) Antton Alberdi, Ostaizka Aizpurua, Joxerra Aihartza et Inazio Garin, « Unveiling the factors shaping the distribution of widely distributed alpine vertebrates, using multi-scale ecological niche modelling of the bat Plecotus macrobullaris », Frontiers in Zoology, BMC et Springer Science+Business Media, vol. 11, no 1, , p. 77 (ISSN 1742-9994, OCLC 56719374, PMID 25389444, PMCID 4226887, DOI 10.1186/S12983-014-0077-6)
- Arthur & Lemaire (2015), p. 441-442, « Terrains de chasse »
- (fr) Jacques Gilliéron, Cyril Schönbächler, Céline Rochet et Manuel Ruedi, Atlas des chauves-souris du bassin genevois, Genève, Faune Genève - Volume 1. CCO-Genève, , 262 p. (ISBN 978-2-8399-1682-0), p. 204-211
- (de) Harald Pieper, « Fledermäuse aus Schleiereulen-Gewöllen von der Insel Kreta », Zeitschrift für Säugetierkunde, Elsevier, vol. 42, , p. 7-12 (ISSN 0044-3468, lire en ligne)
- (en) Petr Benda, Panagiotis Georgiakakis, Christian Dietz, Vladimír Hanák, Kosmoula Galanaki, Vessa Markantonatou, Adéla Chudárková, Pavel Hulva et Ivan Horáček, « Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 7. The bat fauna of Crete, Greece », Acta Societatis Zoologicae Bohemicae, Czech Zoological Society (d), vol. 72, , p. 105-190 (ISSN 1211-376X)
- (en) Konrad Sachanowicz, Ján Krištofík et Mateusz Ciechanowski, « Spinturnicid mites of bats in Albania – host spectrum and morphometrics as a tool of species separation », Journal of Natural History, Taylor & Francis, vol. 48, nos 43-44, , p. 2661-2674 (ISSN 0022-2933 et 1464-5262, DOI 10.1080/00222933.2014.939729)
- (en) Ahmet Karataş et Ferhat Toprak, « The wing mites (Acari: Spinturnicidae) of the Turkish bats, including new records », Kuwait Journal of Science, vol. 49, no 2, (ISSN 2307-4108 et 2307-4116, DOI 10.48129/KJS.11569, lire en ligne)
- (en) Petr Benda, Martin Ševčík, Ferdinand Bego, Konrad Sachanowicz, Friederike Spitzenberger, Přemysl Tájek, Pavla Tájková et Marcel Uhrin, « Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 15. The fauna of bats and bat ectoparasites of Albania with a catalogue of bats from the western Balkans in the collection of the National Museum, Prague », Acta Societatis Zoologicae Bohemicae, Czech Zoological Society (d), vol. 83, , p. 1-238 (ISSN 1211-376X)
- (en) Ahmet Bursali, Adem Keskin, Eray Şimşek, Aysun Keskin et Saban Tekin, « A survey of ticks (Acari: Ixodida) infesting some wild animals from Sivas, Turkey », Experimental and Applied Acarology, Springer Science+Business Media, vol. 66, no 2, , p. 293-299 (ISSN 0168-8162 et 1572-9702, OCLC 37785917, PMID 25784071, DOI 10.1007/S10493-015-9898-Z)
- Union internationale pour la conservation de la nature
- Alberdi & Aizpurua (2018), p. 28-29, « Distribution »
- (en) Inazio Garin, Juan L. García-Mudarra, José R. Aihartza, Urtzi Goiti et Javier Juste, « Presence of Plecotus macrobullaris (Chiroptera: Vespertilionidae) in the Pyrenees », Acta Chiropterologica, Musée et institut de zoologie de l'Académie polonaise des sciences (d), vol. 5, no 2, , p. 243-250 (ISSN 1508-1109 et 1733-5329, DOI 10.3161/001.005.0207)
- Arthur & Lemaire (2015), p. 440-441, « Connaissance des populations »
- (en) Roberta Chirichella, Sabrina Mattiroli, Mosè Nodari, Damiano G. Preatoni, Lucas A. Wauters, Guido Tosi et Adriano Martinoli, « The Adamello-Brenta Natural Park bat community (Mammalia, Chiroptera): distribution and population status », Hystrix, Rome, Associazione Teriologica Italiana (d), vol. 14, nos 1-2, , p. 29-45 (ISSN 0394-1914 et 1825-5272, OCLC 756316901, DOI 10.4404/HYSTRIX-14.1-2-4314)
- (en) Ilaria Trizio, Damiano Preatoni, Roberta Chirichella, Sabrina Mattiroli, Mosè Nodari, Simone Crema, Guido Tosi et Adriano Martinoli, « First record of the alpine long-eared bat (Plecotus macrobullaris Kuzjakin, 1965) in Lombardy (Northern Italy) revealed by DNA analysis », Natura Bresciana, Musée civique des sciences naturelles (d), vol. 34, , p. 171-175 (ISSN 0391-156X, OCLC 2441299)
- (it) Marzia Mattei-Roesli, « Situazione del genere Plecotus (Chiroptera) nel Cantone Ticino (Svizzera) », Bollettino della Società ticinese di Scienze naturali, vol. 98, , p. 31-34
- Dietz & Kiefer (2015), p. 360, « Répartition »
- (en) Ivana Budinski, Branko Karapandža, Vukašin Josipović, Jelena Jovanović et Milan Paunović, « The first record of alpine long-eared bat Plecotus macrobullaris in Serbia », Turkish Journal of Zoology, vol. 40, , p. 984-988 (ISSN 1300-0179, 1303-6114 et 1011-0895, DOI 10.3906/ZOO-1505-20, lire en ligne)
- (en) Primož Presetnik, Milan Paunović, Branko Karapandža, Marina Đurović, Čeda Ivanović, Maša Ždralević, Petr Benda et Ivana Budinski, « Distribution of bats (Chiroptera) in Montenegro », Vespertilio, vol. 17, , p. 129-156 (ISSN 1213-6123, lire en ligne)
- (en) Konrad Sachanowicz et Mateusz Ciechanowski, « Plecotus macrobullaris – new bat species for Albanian fauna (Chiroptera: Vespertilionidae) », Lynx, Musée national de Prague, vol. 37, , p. 241-246 (ISSN 0024-7774 et 1804-6460)
- (en) Jan Piet Bekker et Jan Boshamer, « First records of Plecotus auritus (Linnaeus, 1758) and Plecotus macrobullaris Kuzjakin, 1965 in the Republic of Macedonia », Lutra, vol. 50, no 1, , p. 43-48 (ISSN 0024-7634, lire en ligne)
- (en) Andreas Kiefer, Phylogeny of Western Palaearctic long-eared bats (Mammalia, Chiroptera, Plecotus) – a molecular perspective, Mainz, Johannes Gutenberg-Universität, , 132 p. (lire en ligne)
- (en) Petr Benda, Michal Andreas, Dieter Kock, Radek K. Lučan, Pavel Munclinger, Petra Nová, Ján Obuch, Katarzyna Ochman, Antonín Reiter, Marcel Uhrin et Dita Weinfurtova, « Bats (Mammalia: Chiroptera) of the Eastern Mediterranean. Part 4. Bat fauna of Syria: distribution, systematics, ecology », Acta Societatis Zoologicae Bohemoslovenicae, vol. 70, , p. 1-329 (ISSN 0862-5247, lire en ligne)
- Arthur & Lemaire (2015), p. 441, « Milieux »
- (en) Igor Pavlinić et Nikola Tvrtković, « Altitudinal distribution of four Plecotus species (Mammalia, Vespertilionidae) occurring in Croatia », Natura Croatica, Musée d'histoire naturelle de Croatie (d), vol. 13, no 4, , p. 395-401 (ISSN 1330-0520 et 1848-7386, OCLC 28837675)
- (ru) Alexander Petrovitch Kuzyakin, « Otrjad Rukokrylyje. Ordo Chiroptera », dans Nikolai Alekseevich Bobrinski, Boris Aleksandrovich Kuznetsov & Alexander Petrovitch Kuzyakin, Opredelitel' mlekopitayushchikh SSSR, Moscou, Prosveshtshenije, , p. 79-116
- (en) Friederike Spitzenberger, Jaroslav Piálek et Elisabeth Haring, « Systematics of the genus Plecotus (Mammalia, Vespertilionidae) in Austria based on morphometric and molecular investigations », Folia Zoologica, Institut de Biologie des vertébrés de l'Académie tchèque des sciences (d), vol. 50, no 3, , p. 161-172 (ISSN 0139-7893 et 1210-9681)
- (en) Dieter Kock, « The publication dates of Plecotus alpinus Kiefer and Veith, 2002 and of Plecotus microdontus Spitzenberger, 2002 », Acta Chiropterologica, Musée et institut de zoologie de l'Académie polonaise des sciences (d), vol. 4, no 2, , p. 219-220 (ISSN 1508-1109 et 1733-5329, DOI 10.3161/001.004.0212)
- Alberdi & Aizpurua (2018), p. 26, « Context and Content »
- (en) Antton Alberdi, M. Thomas P. Gilbert, Orly Razgour, Ostaizka Aizpurua, Joxerra Aihartza et Inazio Garin, « Contrasting population-level responses to Pleistocene climatic oscillations in an alpine bat revealed by complete mitochondrial genomes and evolutionary history inference », Journal of Biogeography, Wiley-Blackwell, vol. 42, no 9, , p. 1689-1700 (ISSN 0305-0270 et 1365-2699, OCLC 45446950, DOI 10.1111/JBI.12535)
- Alberdi & Aizpurua (2018), p. 27, « Nomenclatural Notes »
- (en) Sergei V. Kruskop, Ilya V. Artyushin, Alexandr P. Yuzefovich, Undrakhbayar Enkhbat, Anna S. Speranskya, Alexandra A. Lisenkova, Anna Andreevna Bannikova et Vladimir S. Lebedev, « Genetic diversity of Mongolian long-eared bats (Plecotus; Vespertilionidae; Chiroptera) », Acta Chiropterologica, Musée et institut de zoologie de l'Académie polonaise des sciences (d), vol. 22, no 2, (ISSN 1508-1109 et 1733-5329, DOI 10.3161/15081109ACC2020.22.2.002)
- Alberdi & Aizpurua (2018), p. 31-32, « Conservation »
- Dietz & Kiefer (2015), p. 363, « Menaces »
- Alberdi & Aizpurua (2018), p. 26, « Abstract »