Phlorotanin
Les phlorotanins ou phlorotannins (du grec phloios, phlor-, « cortex », et tannin)[2] sont un type de tannin que l'on trouve dans les algues brunes et les diatomées. Ces composés sont des oligomères de phloroglucinol[3].

Les phlorotanins sont une composante de la paroi cellulaire des algues brunes, mais il apparaît qu'ils ont plusieurs autres rôles secondaires. Ils ont des effets anti-oxydants, antibactériens, radioprotectifs et pourraient notamment jouer un rôle dans la défense contre les herbivores.
Plus rare chez les diatomées, elle est notamment présente chez Haslea ostrearia, navicule dont la production d'un pigment bleu (tanin appelé marennine) colore les branchies d’huîtres en affinage[4].
Biosynthèse et localisation des phlorotanins
La biosynthèse des phlorotanins demeure obscure, mais il apparaît qu'ils soient formés à partir du phloroglucinol par synthèse de polycétides (acetate-malonate pathway)[5]. Il a été mis en évidence, qu’une polycétide synthase de type III (PKSIII), prédite dans le génome de l’algue brune Ectocarpus siliculosus, permet la formation de phloroglucinol à partir de malonyl-CoA. C’est une étape majeure dans la formation des phlorotanins. Cette fonction enzymatique a été montré après expression hétérologue dans E.coli du gène EsiPKS et caractérisation du produit après ajout du substrat. Il a également été démontré que in vivo l’expression de EsiPKS est corrélée avec la quantité de phloroglucinol chez une souche Ectocarpus d’eau douce lors de son acclimatation à la salinité. De plus, il semble que ce gène ait été acquis d’une actino-bacterie par transfert horizontal[6]. Les mécanismes menant à la condensation des monomères de phloroglucinol en différents phlorotanins restent à élucider.
On les trouve à l'intérieur des cellules des algues brunes dans des vésicules nommées physodes[3], où sont séquestrés les phlorotanins solubles et polaires, et dans la paroi cellulaire sous forme insoluble[7] - [8]. Leur concentration serait plus importante dans les cellules végétatives du crampon de fixation et le plus souvent de la couche externe du cortex du thalle[9], du moins chez certaines espèces de la famille des Laminariaceae[10]. Il apparaît que la concentration en phlorotanins varie de façon considérable entre les espèces de phaeophycées de même qu'au sein d'une même espèce, selon le milieu, la synthèse de ces composés étant stimulée par divers facteurs environnementaux[11]. Les phlorotanins sont également exsudés par les algues brunes[12] - [13].
Il a été proposé que les phlorotanins sont d'abord séquestrés à l'intérieur des physodes avant d'être complexés à l'acide alginique de la paroi cellulaire par une peroxydase[14]. À ce jour, peu est su de la synthèse des phlorotanins[5]. La formation des physodes, qui contiennent des composés phénoliques chez plusieurs espèces de végétaux, a été l'objet de nombreuses recherches au cours des dernières années. On pensait que ces organites étaient synthétisés dans les chloroplastes ou dans leurs membranes, mais des études récentes suggèrent que leur formation est reliée au réticulum endoplasmique et à l'appareil de Golgi[15].
Structure
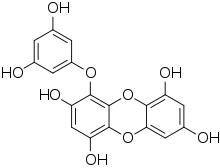
Les phlorotanins sont classés en fonction de l'arrangement de leur monomères de phloroglucinol. Plus de 150 composés sont connus, dont la masse moléculaire varie de 126 Da à650 kDa[3] - [16]. La plupart font entre 10 et 100 kDa[17].
On les classe en six groupes principaux : les fucols, les phloréthols, les fucophlorétols, les fuhalols et les eckols, que l'on trouve seulement chez les Ariaceae[5] - [18].
Le fucodiphlorethol G, chez Ecklonia cava[19], l'eckol chez les espèces d'Ecklonia[20] et le phlorofucofuroeckol chez Eisenia arborea[21] sont des exemples de phlorotannins.
Rôles des phlorotanins
La raison d'être des phlorotanins fait toujours l'objet de recherches (2012). Ils apparaît qu'ils sont à la fois impliqués dans les métabolismes primaire et secondaire[14], tant à l'échelle cellulaire qu'organismique[22].
Rôle structurel
Le rôle structurel des phlorotanins dans la paroi cellulaire est une fonction primaire de ces composés phénoliques[14] - [5]. Toutefois, comme des recherches ont démontré que les phlorotanins sont plus abondants dans le cytoplasme que dans la paroi cellulaire, ce rôle primaire pourrait toutefois ne pas être le principal[23].
Rôle reproductif
Les phlorotanins présents dans le cytoplasme, de même que ceux exsudés, semblent jouer un rôle dans la reproduction des algues brunes en contribuant à la formation de la paroi cellulaire du zygote[8], et peut-être en évitant la fertilisation multiple en inhibant le mouvement des spermatozoïdes[5].
Rôles secondaires
Des études ont démontré que les phlorotanins semblent protéger les algues brunes de plusieurs manières. Voici quelques exemples.

Défense contre l'herbivorisme
Comme certaines études ont démontré que l'herbivorie peut induire la synthèse de phlorotanins. Il a été suggéré que ces derniers jouent un rôle défensif contre les herbivores, notamment en réduisant la digestion chez certains poissons par la formation d'un complexe phlorotannins-protéines[11]. Toutefois, les résultats des recherches menées en ce sens montrent que cette relation est ambiguë[24] - [25] - [26]. Le rôle dissuasif des phlorotanins quant à l'herbivorisme pourrait être hautement dépendant à la fois de l'espèce de l'algue et de l'herbivore[27].
Protection de la radiation UV et des métaux lourds
Les phlorotanins sont principalement localisés à la périphérie des cellules, en tant que composants de la paroi cellulaire. Aussi, ils absorbent la lumière à des longueurs d'onde entre 280 et 320 nm[28], correspondant aux UV-B, et montrent des pics d'absorption à 200 et 265 nm[3], ce qui correspond à des longueurs d'onde incluses dans les UV-C. Des études in situ ont démontré que la production de phlorotanins est positivement influencée par l'intensité du rayonnement solaire chez Ascophyllum nodosum et Fucus vesiculosus[29]. Pour ces raisons, il a été suggéré que les phlorotanins jouent un rôle photoprotecteur[30]. Des études menées avec les espèces Lessonia nigrescens[30] et Macrocystis integrifolia[31] montrent que les radiations UV-A et les UV-B peuvent induire la synthèse de phlorotanins solubles et que cette dernière est corrélée avec la réduction de l'inhibition de la photosynthèse, de même qu'avec la réduction des dommages à l'ADN, deux des principaux effets de la radiation ultraviolette sur les cellules végétales. De plus, le fait que ces composés phénoliques sont exsudés dans le milieu environnant permet de réduire l'exposition aux UV auxquels sont soumis les méiospores des algues, le phytoplancton et les autres habitants des forêts de kelp, où la biomasse algale est élevée et où l'eau est moins brassée[31].
Les phlorotanins pourraient aussi être impliqués dans la séquestration de métaux lourds tels les ions Sr2+, Mg2+, Ca2+, Be2+, Mn2+, Cd2+, Co2+, Zn2+, Ni2+, Pb2+ et Cu2+[32]. Toutefois, si les propriétés chélatrices de certains phlorotanins ont été démontrées in vitro, des études in situ suggèrent que ces caractéristiques sont propres à certaines espèces[33] - [34].
Effet antibiotique
Des études ont démontré que les phlorotanins peuvent agir en tant qu'antibiotique contre certaines espèces de dinoflagellés[35].
Propriétés thérapeutiques
Il a été démontré que les phlorotanins peuvent avoir des effets anti-diabétiques, anti-cancérigènes, anti-oxydants, antibactériens, radioprotectifs et anti-VIH[36] - [37].
Applications en cosmétologie
Effet antioxydant
Source[40].
Les phlorotannins comme le dieckol, eckol et eckstolonol de l'algue brune Ecklonia cava ou encore le diphlorethohydroxycarmalol de l'algue brune Ishige okamurae possèdent des propriétés antioxydantes. Ces composés sont utilisés pour la protection aux UV en diminuant les dommages à l'ADN par les UV-B. En effet, ces phloratannins contenus dans ces algues brunes permettent le contrôle de la fragmentation nucléaire dans les cellules de fibroblaste, composant essentiel à la souplesse et la cohérence du derme. D'autres mécanismes seraient aussi contrôlés par les phlorotannins d'algues brunes comme le blocage de l'apoptose ou encore le blocage des mauvais fonctionnements de la mitochondrie.
Augmentation de la croissance capillaire
Source[41].
Des métabolites secondaires comme le 7-phloeckol ou le dieckol provenant d'algues brunes interviennent dans l'augmentation de la croissance capillaire. Il a été démontré qu'il y aurait une augmentation de la prolifération des follicules pileux chez l'humain lorsque l'on traite ces derniers avec ces métabolites secondaires.
Effet blanchissant de la peau
Source[42].
L'alcool 4-hydroxyphenethyl de l'algue brune Hizikia fusiformis via son activité inhibitrice de la tyrosinase intervient au niveau de la biosynthèse de la mélanogenèse pour en inhiber la synthèse de mélanine, pigment de la peau lui donnant sa couleur plus ou moins colorée et protégeant la peau des effets néfastes des UV. La concentration inhibitrice médiane (IC50) de ce phlorotannin est plus faible que les composés actuels utilisés en tant qu'agent blanchissant de la peau en Asie tels que l'acide kojique ou encore l'arbutine et semblerait donc être plus efficace. Cette inhibition de la synthèse de mélanine par ces phlorotannins passerait par la voie de signalisation cellulaire ERK.
Effet anti-rides
Source[43].
Les métalloprotéinases matricielles (MMPs) sont des constituants de la matrice extra cellulaire (MEC) avec une activité protéolytique permettant le remodelage et la dégradation du derme. Il a été montré que certains phlorotannins comme l'eckol et le dieckol de l'algue brune Ecklonia stolonifera et le 6,6'-bieckol et le dioxinodehydroeckol de l'algue brune Ecklonia cava inhibent l'activité protéolytique de ces MMPs à l'origine de la dégradation du collagène et donc de l'apparition des rides. Cette inhibition de la dégradation du collagène par ces phlorotannins passerait par la voie de signalisation cellulaire NF-kB.
Références
- Marc-André Selosse (ill. Arnaud Rafaelian), Les Goûts et les couleurs du monde: Une histoire naturelle des tannins, de l'écologie à la santé, Actes Sud, , p. 87
- (en) Alexander Senning, The Etymology of Chemical Names, Walter de Gruyter GmbH & Co KG, , p. 324
- M.A. Ragan et K.W. Glombitza. 1986. Phlorotannins, brown algal polyphenols. Progress in Phycological Research, 4, 129-241.
- (en) Jean-Bernard Pouvreau et al, « Preliminary characterisation of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen », Journal of Applied Phycology, vol. 18, no 6, , p. 757–767.
- Riitta Koivikko, 2008, Brown algal phlorotannins: Improving and applying chemical methods, Ph. D. Thesis, University of Turku, Turku, Finland.
- (en) Laurence Meslet-Cladière, Ludovic Delage, Cédric J.-J. Leroux et Sophie Goulitquer, « Structure/Function Analysis of a Type III Polyketide Synthase in the Brown Alga Ectocarpus siliculosus Reveals a Biochemical Pathway in Phlorotannin Monomer Biosynthesis », The Plant Cell, vol. 25, no 8, , p. 3089–3103 (ISSN 1040-4651 et 1532-298X, PMID 23983220, DOI 10.1105/tpc.113.111336, lire en ligne, consulté le )
- Schoenwaelder, M. E. A. 2002. The occurrence and cellular significance of physodes in brown algae. Phycologia, 41, 125–139.
- Schoenwaelder, M. E. A. et Clayton, M. N. 1998. Secretion of phenolic substances into the zygote wall and cell plate in embryos of Hormosira and Acrocarpis (Fucales, Phaeophyceae). Journal of Phycology, 34, 969–980.
- Selon le principe de l'allocation des ressources, l'investissement énergétique dans la synthèse de phlorotannins se fait au détriment de la croissance. Ce principe explique que les algues économisent leurs ressources en les synthétisant au niveau des crampons de fixation ou le plus souvent dans leurs tissus de surface, que leur production reste limitée quand la pression d'herbivorie reste faible ou au printemps lorsque la croissance consomme toutes ces ressources.
- T. Shibata, S. Kawaguchi, Y. Hama, M. Inagaki, K. Yamaguchi et T. Nakamura. 2004. Local and chemical distribution of phlorotannins in brown algae. Journal of Applied Phycology, 16, 291-296.
- V. Jormalainen, T. Honkanen, R. Koivikko et J. Eränen. 2003. Induction of phlorotannin production in a brown alga: defense or resource dynamics?. Oikos, 103, 640–650.
- M. A. Ragan et A. Jensen. 1978. Quantitative studies on brown algal phenols. II. Seasonal variation in polyphenol content of Ascophyllum nodosum (L.) Le Jol. and Fucus vesiculosus (L.). Journal of Experimental Marine Biology and Ecology, 34 (3), 245-258. DOI 10.1016/S0022-0981(78)80006-9
- J.S. Jennings et P.D. Steinberg. 1994. In situ exudation of phlorotannins by the sublittoral kelp Ecklonia radiata. Mar. Biol., 121, 349–354.
- T. M. Arnold et N. M. Targett. 2003. To grow and defend: lack of tradeoffs for brown algal phlorotannins. Oikos, 100 (2), 406-408.
- M.E.A. Schoenwaelder et M.N. Clayton. 2000. Physode formation in embryos of Phyllospora comosa and Hormosira banksii (Phaeophyceae). Phycologia, 39, 1-9.
- M.E. Hay et W. Fenical. 1988. Marine plant–herbivore interactions: the ecology of chemical defense. Annu. Rev. Ecol. Syst., 19, 111–45.
- A.A. Boettcher et N.M. Targett. 1993. Role of polyphenolic molecular-size in reduction of assimilation efficiency in Xiphister mucosus. Ecology, 74, 891-903.
- N.M. Targett et T. M. Arnold. 1998. Predicting the effects of brown algal phlorotannins on marine herivores in tropical and temperate oceans. Journal of Phycology, 34, 195-205.
- Young Min Ham, Jong Seok Baik, Jin Won Hyun et Nam Ho Lee, Bull. 2007. Isolation of a new phlorotannin, fucodiphlorethol G, from a brown alga Ecklonia cava « Copie archivée » (version du 25 avril 2012 sur Internet Archive). Korean Chem. Soc., 28 (9), 1595.
- Changjong Moon, Sung-Ho Kim, Jong-Choon Kim, Jin Won Hyun, Nam Ho Lee, Jae Woo Park et Taekyun Shin. 2008. Protective effect of phlorotannin components phloroglucinol and eckol on radiation-induced intestinal injury in mice. Phytotherapy Research, 22 (2), 238–242.
- Y. Sugiura, K. Matsuda, Y. Yamada, M. Nishikawa, K. Shioya, H. Katsuzaki, K. Imai et H. Amanoo. 2006. Isolation of a new anti-allergic phlorotannin, phlorofucofureckol-B from an edible brown alga, Eisenia arborea. Biosci. Biotechnol. Biochem., 70, 60417-1-5.
- Monica E. A. Schoenwaelder. 2002. The occurrence and cellular significance of physodes in brown algae. Phycologia 41(2): 125-139.
- R. Koivikko, J. Loponen, T. Honkanen et V. Jormalainen. 2005. Contents of soluble, cell-wall-bound and exuded phlorotannins in the brown alga Fucus vesiculosus, with implications on their ecological functions. Journal of Chemical Ecology 31(1):195-212. DOI 10.1007/s10886-005-0984-2
- A. Hemmi, T. Honkanen et V. Jormalainen. 2004. Inducible resistance to herbivory in Fucus vesiculosus–duration, spreading and variation with nutrient availability. Marine Ecology Progress Series 273:109–120. DOI 10.3354/meps273109
- S. Rohde, M. Molis et M. Wahl. 2004. Regulation of anti-herbivore defence by Fucus vesiculosus in response to various cues. Journal of Ecology 92:1011-1018.DOI 10.1111/j.0022-0477.2004.00936.x
- F. Haavisto, T. Välikangas et V. Jormalainen. 2010. Induced resistance in a brown alga: phlorotannins, genotypic variation and fitness costs for the crustacean herbivore. Oecologia 162:685-695. DOI 10.1007/s00442-009-1494-7
- C.D. Amsler, V.A. Fairhead. 2006. Defensive and sensory chemical ecology of brown algae. Adv. Bot. Res. 43:1–91.
- H. Pavia, G. Cervin, A. Lindgren et Per Aberg. 1997. Effects of UV-B radiation and simulated herbivory on phlorotannins in the brown alga Ascophyllum nodosum. Marine Ecology Progress Series 157:139-146.
- Henrik Pavia et Gunilla B. Toth. 2000. Influence of light and nitrogen on the phlorotannin content of the brown seaweeds Ascophyllum nodosum and Fucus vesiculosus, Hydrobiologia 440: 299-305
- Ivan Gómez et Pirjo Huovinen. 2010. Induction of phlorotannins during exposure mitigates inhibition of photosynthesis and DNA damage in the kelp Lessonia nigrescens. Photochemistry and Photobiology 86:1056-1063.DOI 10.1111/j.1751-1097.2010.00786.x
- A. K. Swanson et L. D. Druehl. 2002002. Induction, exudation and the protective role of kelp phlorotannins. Aquatic botany 73:241-253.
- Mark A Ragan, Olav Smidsrød et Bjørn Larsen. 1979. Chelation of divalent metal ions by brown algal polyphenols. Marine Chemistry 7(3): 265–271. DOI 10.1016/0304-4203(79)90043-4
- P. Huovinen, P. Leal et I. Gómez. 2010. Interacting effects of copper, nitrogen and ultraviolet radiation on the physiology of three south Pacific kelps. Marine and Freshwater Research 61:330-341.
- Toth G. et Pavia H. 2000. Lack of phlorotannin induction in the brown seaweed Ascophyllum nodosum in response to increased copper concentrations. Marine Ecology Progress Series 192: 119-126. Modèle:INIST
- Koki Nagayama, Toshiyuki Shibata, Ken Fujimoto, Tuneo Honjo et Takashi Nakamura. 2003. Algicidal effect of phlorotannins from the brown alga Ecklonia kurome on dinoflagellates. Aquaculture 218(1-4): 601-611. DOI 10.1016/S0044-8486(02)00255-7
- S. Gupt et N. Abu-Ghannam. 2011. Bioactive potential and possible health effects of edible brown seaweed. Trends in Food Science & Technology, 22, 315-326, DOI 10.1016/j.tifs.2011.03.011
- Yong-Xin Li, Isuru Wijesekara, Yong Li et Se-Kwon Kim. 2011. Phlorotannins as bioactive agents from brown algae. Process Biochemistry, 46, 2219-2224, DOI 10.1016/j.procbio.2011.09.015
- Kalu Kapuge Asanka Sanjeewa, Eun-A Kim, Kwang-Tae Son et You-Jin Jeon, « Bioactive properties and potentials cosmeceutical applications of phlorotannins isolated from brown seaweeds: A review », Journal of Photochemistry and Photobiology B: Biology, vol. 162, , p. 100–105 (ISSN 1011-1344, DOI 10.1016/j.jphotobiol.2016.06.027, lire en ligne, consulté le )
- Hyoseung Shin, A-Ri Cho, Dong Young Kim et Semchin Munkhbayer, « Enhancement of Human Hair Growth Using Ecklonia cava Polyphenols », Annals of Dermatology, vol. 28, no 1, , p. 15 (ISSN 1013-9087 et 2005-3894, DOI 10.5021/ad.2016.28.1.15, lire en ligne, consulté le )
- Soo-Jin Heo, Seok-Chun Ko, Seon-Heui Cha et Do-Hyung Kang, « Effect of phlorotannins isolated from Ecklonia cava on melanogenesis and their protective effect against photo-oxidative stress induced by UV-B radiation », Toxicology in Vitro, vol. 23, no 6, , p. 1123–1130 (ISSN 0887-2333, DOI 10.1016/j.tiv.2009.05.013, lire en ligne, consulté le )
- Jung-Il Kang, Sang-Cheol Kim, Min-Kyoung Kim et Hye-Jin Boo, « Effect of Dieckol, a Component of Ecklonia cava, on the Promotion of Hair Growth », International Journal of Molecular Sciences, vol. 13, no 5, , p. 6407–6423 (ISSN 1422-0067, DOI 10.3390/ijms13056407, lire en ligne, consulté le )
- Na Young Yoon, Tae-Kil Eom, Moon-Moo Kim et Se-Kwon Kim, « Inhibitory Effect of Phlorotannins Isolated from Ecklonia cava on Mushroom Tyrosinase Activity and Melanin Formation in Mouse B16F10 Melanoma Cells », Journal of Agricultural and Food Chemistry, vol. 57, no 10, , p. 4124–4129 (ISSN 0021-8561 et 1520-5118, DOI 10.1021/jf900006f, lire en ligne, consulté le )
- Noel Thomas et Se-Kwon Kim, « Beneficial Effects of Marine Algal Compounds in Cosmeceuticals », Marine Drugs, vol. 11, no 12, , p. 146–164 (ISSN 1660-3397, DOI 10.3390/md11010146, lire en ligne, consulté le )
Liens externes
- (en) Riitta Koivikko (2008), Brown algal phlorotannins: Improving and applying chemical methods, Ph. D. Thesis, University of Turku, Turku, Finland.
- (en) S. Gupt et N. Abu-Ghannam (2011), Bioactive potential and possible health effects of edible brown seaweed, Trends in Food Science & Technology, 22, 315-326, DOI 10.1016/j.tifs.2011.03.011.