Stenonychosaurus
Stenonychosaurus inequalis
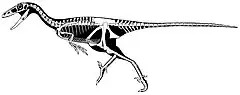
ex- « Troodon inequalis ».
Stenonychosaurus (qui signifie « lézard à griffes étroites ») est un genre éteint de dinosaures troodontidés ayant vécu au Campanien (Crétacé supérieur), soit il y a environ entre 83,6 à 72,1 millions d'années, et découvert dans la formation de Dinosaur Park en Alberta, au Canada, ainsi que (possiblement) dans la formation de Two Medicine à cheval sur le Montana et l'Alberta.
L'espèce type et seule espèce du genre, Stenonychosaurus inequalis, a été nommée par Charles Mortram Sternberg en 1932, basée sur un pied, des fragments de main et des vertèbres caudales. S. inequalis a été réaffectée en 1987 par Phil Currie au genre Troodon. En 2017, D. C. Evans et ses collègues[1], puis A. J. Van der Reest et Philip John Currie[2] l'assignent à un nouveau genre, Stenonychosaurus, distinct de Troodon, désormais considéré comme un genre possiblement douteux[1] - [2] - [3].
Description

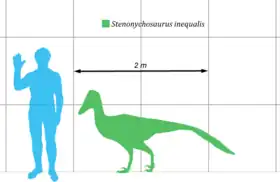




Stenonychosaurus était un petit dinosaure, mesurant jusqu'à 0,90 mètre de hauteur, 2,40 mètres de longueur[4] et pesait 35 à 50 kg[5] - [6]. Les plus grands spécimens sont de taille comparable à celles de Deinonychus et de Unenlagia[7] - [8]. Ses membres sont très longs et élancés suggérant que cet animal était capable de courir rapidement. Il possédait de grandes griffes rétractiles en forme de faucille sur le deuxième orteil, qui étaient redressées pendant la course.
Ses yeux étaient très grands (suggérant peut-être un style de vie partiellement nocturne) et légèrement tournés vers l'avant, ce qui donnait à Stenonychosaurus une certaine perception de la profondeur[9].
Cerveau et oreille interne
Parmi les dinosaures, Stenonychosaurus possédait l'un des plus gros cerveaux par rapport à sa masse corporelle (un rapport comparable à celui des oiseaux modernes)[10]. Le rapport cerveau / volume cérébral varie entre 31,5% et 63%, allant de celui des reptiles non aviens à celui des oiseaux[11] - [12]. De plus, il avait des crêtes osseuses soutenant les membranes tympaniques qui étaient ossifiées au moins dans leurs régions supérieure et inférieure. Les autres crêtes étaient soit cartilagineuses, soit trop délicates pour avoir été préservées. Ses pattes métotiques sont élargies comme chez Dromaeosaurus et certains oiseaux primitifs comme Archaeopteryx et Hesperornis[11] - [12].
Spécimens et caractéristiques distinctives
L'holotype de Stenonychosaurus inequalis, référencé CMN 8539, est un squelette partiel constitué de six vertèbres caudales, d'os de la main, de l'extrémité distale du tibia et de l'astragale gauches et d'un pied gauche complet. Quelques spécimens crâniens ont également été référés au genre : UALVP 52611 (un toit de crâne presque complet) et TMP 1986.036.0457 (une boîte crânienne partielle).
De nombreux spécimens de troodontidés de Dinosaur Park autrefois référés au genre Troodon ont été en 2017 référés à Stenonychosaurus et Latenivenatrix[1] - [2]. Les fossiles maintenant attribués à Stenonychosaurus comprennent plusieurs frontaux (UALVP 5282, TMP 1986.078.0040, TMP 1988.050.0088, TMP 1991.036.0690), un dentaire partiel (TMP 1982.019.0151) et la partie distale d’un métatarsien gauche III (TMP). 1998.068.0090).
Ce genre se distingue des autres troodontidés (en particulier de son proche parent Latenivenatrix) par les caractéristiques suivantes :
- ses métatarses III possèdent une surface antérieure convexe ;
- un frontal en forme de « L » avec un contact naso-frontal plat et ridé de manière antéropostérieure.
Histoire et découverte
L'historique des découvertes et de la taxonomie des troodontidés est longue et tourmentée. La définition du genre Troodon en se basant sur un seul fragment d'une petite dent est à l'origine de toutes ces difficultés. Beaucoup de dents isolées puis de parties de squelettes ont longtemps été systématiquement attribués au genre Troodon jusqu'en 1932, où Charles Mortram Sternberg attribue des os sont au genre Stenonychosaurus, sous le nom binominal de Stenonychosaurus inequalis.
Un squelette plus complet de Stenonychosaurus a été décrit par Dale Russell en 1969 à partir de la même formation géologique, qui a finalement constitué le fondement scientifique d'une célèbre sculpture grandeur nature de Stenonychosaurus, accompagnée de son descendant fictif et humanoïde, le « dinosauroïde ».
Phil Currie a relancé le débat en 1987, en concluant que les différences supposées dans la structure de la dent et de la mâchoire parmi les troodontidés et les saurornithoididés étaient liées à l'âge des spécimens âge et la position de la dent dans la mâchoire, plutôt que sur une différence d'espèces[13]. P. Currie a ainsi assigné tous les fossiles de troodontidés d'Amérique du Nord à l'espèce Troodon formosus, ce qui a été largement adoptée par les paléontologues, et tous les spécimens autrefois appelés Stenonychosaurus ont été renommés Troodon dans la littérature scientifique du début du XXIe siècle.
En 2017, Evans et ses collègues ont souligné de la nature non diagnostique de l'holotype de Troodon formosus et ont suggéré que les squelettes découverts dans la formation de Dinosaur Park soient attribués au genre Stenonychosaurus[1]. Plus tard au cours de la même année, Aaron J. van der Reest et P. Currie en arrivent à la même conclusion, mais séparent une grande partie du matériel attribué à Stenonychosaurus pour l'assigner à un nouveau genre : Latenivenatrix[2].
Cependant, ce consensus n'est pas général, car en 2018, Martin Varricchio et ses collègues rejettent ces propositions, en citant le fait que le nom de genre Stenonychosaurus n'avait pas été utilisé depuis trente ans, depuis que Currie et ses collègues l'avaient mis en synonymie avec Troodon. Ils indiquent que, pour eux, « Troodon formosus reste le nom correct de ce taxon »[3].
Le dinosauroïde
En 1982, le paléontologue Dale Russell émit l'hypothèse (une uchronie) que si la grande extinction de la fin du Crétacé n'était pas intervenue il y a environ 66 Ma (millions d'années), Troodon aurait pu évoluer en êtres intelligents avec un corps de type humain, des mains à trois doigts, et un crâne très volumineux par augmentation de son coefficient d'encéphalisation ; un être imaginaire surnommé dinosauroïde. Il a commandé à l'artiste taxidermiste Ron Seguin la réalisation d'une sculpture de ce « dinosauroïde », réalisée en 1982[14], selon ses directives, et qui est exposée entre autres au musée canadien de la nature à Ottawa[15].
Dès les années 1980, cette hypothèse a été fortement critiquée. En effet, la plupart des paléontologues (comme par exemple les paléontologues américains Gregory S. Paul et Thomas R. Holtz, Jr.) estiment que la créature imaginée est bien trop anthropomorphique. En 2006, Darren Naish a fait valoir qu'un troodontidé à cerveau volumineux et très intelligent conserverait un plan corporel de théropodes classiques, avec une posture horizontale et une longue queue, et manipulerait des objets plutôt avec le museau et les pieds à la manière d'un oiseau, plutôt qu'avec des « mains » de type humain[16].
Cependant, la quasi-totalité des restes fossiles attribués à Troodon l'ayant rendu célèbre pour son intelligence imaginée, sont aujourd'hui attribués à Stenonychosaurus inequalis.
Paléobiologie
Stenonychosaurus montre des caractéristiques généralement attribuées aux prédateurs. Il possède en effet une longue griffe en faucille sur chaque pied et une bonne vision binoculaire[17].
Ses mâchoires présentent cependant une symphyse large et en forme de « U » similaire à celle d'un iguane actuel, une espèce de lézards adaptée à un régime herbivore. Une étude comparative de l'appareil d'alimentation suggère par ailleurs que Stenonychosaurus aurait pu être un omnivore[17].
Les dents de Stenonychosaurus sont différentes de celles de la plupart des autres théropodes. Elles portent de larges dentelures appelées denticules dont la pointe est dirigée vers le sommet de chaque dent. Ces dents présentent des facettes d'usure sur leurs côtés. Thomas R. Holtz (1998) a finalement souligné que les caractéristiques mises en avant pour identifier Stenonychosaurus comme un prédateur : ses mains agrippantes, son cerveau de grande taille et sa vision stéréoscopique, sont aussi des caractéristiques communes aux primates herbivores / omnivores et aux ratons laveurs omnivores[17].
Enfin en 2001, un squelette partiel de Stenonychosaurus a été découvert avec des marques de morsures, une observation extrêmement rare sur des os de théropodes carnivores[18].
Reproduction
Des nids et des œufs de dinosaures ont été découverts par John R. Horner en 1983 dans la formation de Two Medicine au Montana[19]. David J. Varriccho et ses collègues ont décrit huit de ces œufs en 2002, qui sont conservés au Museum of the Rockies à Bozeman dans le Montana[20].
Horner a découvert, dans le même niveau stratigraphique et à proximité de ces nids, des os isolés ou des éléments du squelette de dinosaures d'un hypsilophodontidé, Orodromeus[19]. Par contre, en 1996, une étude des embryons fossilisés dans le œufs, conduit Horner et Weishampel à considérer qu'ils appartiennent à « Troodon » et non à Orodromeus[21]. En 1997, Varricchio et ses collègues confirme cette détermination après l'étude d'un squelette partiel de « Troodon » (MOR 748) en contact avec une couvée d'au moins cinq œufs (MOR 750), probablement dans une position de couvaison[22]. En 2017, Van der Reest et Currie retiennent la possibilité que le troodontidé de Two Medicine soit un Stenonychosaurus[2].
Nids
Varricchio et al., dans leur article de 1997, décrivent la structure exacte des nids. Ils sont construits avec des sédiments, en forme de grand plat circulaire d'un diamètre intérieur d'environ 1 mètre, avec un rebord en relief prononcé entourant les œufs. Les nids les mieux conservés contiennent de 16 (MOR 246) à 24 (MOR 963) œufs. Les œufs sont de forme allongée, avec leur extrémité plus conique pointée vers le bas et enfoncée à mi-hauteur dans le sédiment. Les œufs sont inclinés de manière que la moitié supérieure soit en moyenne plus proche du centre du nid. Il n'y a aucune preuve de la présence de matière végétale dans le nid[22]. L'étude de ces nids permet de mieux définir le comportement reproductif des troodontidés. Ils en concluent que la biologie de la reproduction de Stenonychosaurus et des troodontidés est intermédiaire entre celle des crocodiles et celle des oiseaux, un résultat conforme aux résultats de la phylogénie.
Ponte
Les œufs sont statistiquement groupés par paires, ce qui suggère que la femelle avait deux oviductes fonctionnels, comme les crocodiles, et non un seul comme les oiseaux. Les crocodiles pondent de grande quantité d’œufs de petite taille par rapport à celle de l’animal, tandis que les oiseaux en pondent peu et de plus grande taille. Le troodontidé de Two Medicine était dans une situation intermédiaire, pondant des œufs de 500 grammes pour une masse de l'adulte estimée à 50 kg. Ses œufs sont environ 10 fois plus gros que ceux des reptiles de même masse, mais environ deux fois plus petits que pour un oiseau de masse identique[22].
Varricchio et ses collègues ont également mis en évidence que la ponte était itérative. Ainsi la femelle adulte pond une paire d’œufs tous les un ou deux jours, puis assure l'éclosion simultanée en retardant la couvaison jusqu'à ce que tous les œufs soient pondus. Le groupe de fossiles référencé MOR 363 montre 22 œufs vides (éclos) et les embryons trouvés dans les œufs de MOR 246 sont dans des états de développement très similaires, ce qui implique que tous les jeunes ont éclos à peu près simultanément. Les embryons présentaient un développement squelettique avancé et les œufs vides étaient relativement non écrasés, ce qui impliquait que les nouveau-nés avaient un rythme de croissance rapide. Les auteurs ont estimé la durée d'occupation du nid par les adultes, pour la ponte, la couvée et l'éclosion, entre 45 et 65 jours[20].
Couvaison par les mâles ?
Varricchio et ses collègues en 2008 étudient l'histologie des os du spécimen MOR 748 de Two Medicine. Ils constatent qu'il lui manque les figures de résorption osseuse classiques indiquant qu'il s'agissait d'une femelle pondeuse. Ils ont également mesuré le rapport entre le volume total des œufs dans les couvées et la masse corporelle de l'adulte. Ils ont tracé un graphique des corrélations entre ce rapport et le type de stratégies parentales utilisées par les oiseaux et les crocodiles actuels et ont constaté que ce rapport chez le troodontidé était cohérent avec celui des oiseaux où seul le mâle adulte couve les œufs. Ils en ont conclu que les femelles troodontidés ne couvaient probablement pas d’œufs, contrairement aux mâles, ce qui pourrait être une caractéristique partagée entre les dinosaures maniraptoriens et les oiseaux basaux[23]
Cependant ces conclusions ont été remises en cause en 2013 par Geoffrey F. Birchard Geoffrey et ses collègues. Après une étude sur la masse des pontes aviaires, ils concluent que le type de comportement (protection) parental ne peut pas être déterminé à l'aide de méthodes allométriques classiques telles que celle utilisées par Varricchio et ses collègues[24].
Paléoécologie
Stenonychosaurus inequalis est connu de la formation de Dinosaur Park du sud de l'Alberta, au Canada, qui était à l'époque une plaine d'inondation côtière recouverte de forêts tempérées à chaudes. Les superprédateurs de cet environnement étaient des tyrannosaures tels que Daspletosaurus et Gorgosaurus. Les herbivores comprenaient des hadrosauridés tels que Lambeosaurus, Corythosaurus et Prosaurolophus, des cératopsides tels que Styracosaurus, Centrosaurus et Chasmosaurus, des ankylosaures tels que Scolosaurus, Euoplocephalus et Edmontonia, et des pachycéphalosaures tels que Stegoceras et Foraminacephale.
Classification
Le cladogramme suivant résulte de l'analyse phylogénétique des Troodontidae conduite par Aaron van der Reest et Phil Currie en 2017[2] :
| ◄ Troodontidae |
| ||||||||||||||||||||||||||||||||||||
* probablement Stenonychosaurus[2].
Les auteurs ne nomment pas formellement le « troodontidé de la Two Medicine », même s'ils le considèrent comme appartenant probablement au genre Stenonychosaurus[2] :
Voir aussi
Notes et références
Références
- (en) D. C. Evans, T.M. Cullen, D.W. Larson et A. Rego, « A new species of troodontid theropod (Dinosauria: Maniraptora) from the Horseshoe Canyon Formation (Maastrichtian) of Alberta, Canada », Canadian Journal of Earth Sciences, vol. 54, no 8, , p. 813–826 (DOI 10.1139/cjes-2017-0034, Bibcode 2017CaJES..54..813E)
- (en) A. J. van der Reest et P. J. Currie, « Troodontids (Theropoda) from the Dinosaur Park Formation, Alberta, with a description of a unique new taxon: implications for deinonychosaur diversity in North America », Canadian Journal of Earth Sciences, vol. 54, no 9, , p. 919–935 (DOI 10.1139/cjes-2017-0031, Bibcode 2017CaJES..54..919V, hdl 1807/78296, lire en ligne).
- (en) D. J. Varricchio, M. Kundrát et J. Hogan, « An Intermediate Incubation Period and Primitive Brooding in a Theropod Dinosaur », Scientific Reports, no 1, (DOI 10.1038/s41598-018-30085-6, lire en ligne)
- (en) Nick Attfield, Dinosaur Jr.'s You’re Living All Over Me, Bloomsbury Publishing, , 140 p. (ISBN 978-1-4411-8778-9, DOI 10.5040/9781501396823, lire en ligne)
- (en) G. S. Paul, The Princeton Field Guide to Dinosaurs 2nd Edition, Princeton, Princeton University Press, , 160 p. (ISBN 978-0-691-16766-4)
- (en) G.S. Paul, Predatory Dinosaurs of the World, New York, Simon and Schuster, (ISBN 978-0-671-61946-6, lire en ligne), p. 396
- (en) Alan H. Turner, Mark A. Norell, Diego Pol, Julia A. Clarke et Gregory M. Erickson, « A Basal Dromaeosaurid, And Size Evolution, Preceding Avian Flight », Science Magazine, vol. 317, no 5843, , p. 1378–81 (PMID 17823350, DOI 10.1126/science.1144066, Bibcode 2007Sci...317.1378T)
- (en) « Britain's Hoverflies: A Field Guide (2nd edition)2016 053 Stuart Ball and Roger Morris Britain's Hoverflies: A Field Guide (2nd edition) Princeton, NJ and Oxford Princeton University Press 2015 313 pp. (ISBN 9780691164410) (print) (ISBN 9781400866021) (e-book), WILD Guides », Reference Reviews, vol. 30, no 2, , p. 32–32 (ISSN 0950-4125, DOI 10.1108/rr-12-2015-0294, lire en ligne, consulté le )
- (en) Philip J. Currie, « Bird-like characteristics of the jaws and teeth of troodontid theropods (Dinosauria, Saurischia) », Journal of Vertebrate Paleontology, vol. 7, no 1, , p. 72–81 (ISSN 0272-4634 et 1937-2809, DOI 10.1080/02724634.1987.10011638, lire en ligne, consulté le )
- (en) « The Macmillan illustrated encyclopedia of dinosaurs and prehistoric animals: a visual who's who of prehistoric life », Choice Reviews Online, vol. 26, no 08, , p. 26–4483-26-4483 (ISSN 0009-4978 et 1523-8253, DOI 10.5860/choice.26-4483, lire en ligne, consulté le )
- (en) Larsson, H.C.E. 2001. Endocranial anatomy of Carcharodontosaurus saharicus (Theropoda: Allosauroidea) and its implications for theropod brain evolution. pp. 19–33. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press
- (en) Stuart S Sumida, « Mesozoic Vertebrate Life: New Research Inspired by the Paleontology of Philip J. Currie. Life of the Past. Edited by Darren H Tanke and , Kenneth Carpenter; Art Editor: , Michael W Skrepnick. Published by Indiana University Press, Bloomington (Indiana), in association with the National Research Council of Canada. xviii + 577 p + 22 pl; ill.; index. (ISBN 0-253-33907-3). 2001. », The Quarterly Review of Biology, vol. 77, no 2, , p. 194–194 (ISSN 0033-5770 et 1539-7718, DOI 10.1086/343935, lire en ligne, consulté le )
- (en) P. Currie, « Theropods of the Judith River Formation », Occasional Paper of the Tyrrell Museum of Palaeontology, vol. 3, , p. 52–60
- (en) D. A. Russell et R. Seguin, « Reconstruction of the small Cretaceous theropod Stenonychosaurus inequalis and a hypothetical dinosauroid », Syllogeus, vol. 37, , p. 1–43 (lire en ligne)
- (en) Jeff Hecht, « Smartasaurus – If they hadn't been wiped out in a global catastrophe 65 million years ago, could dinosaurs have evolved intelligence? », sur cosmosmagazine.com, (consulté le )
- (en) Naish, D. (2006). Dinosauroids Revisited Darren Naish: Tetrapod Zoology, April 23, 2011
- (en) Holtz, Thomas R., Brinkman, Daniel L., Chandler, Chistine L. (1998) Denticle Morphometrics and a Possibly Omnivorous Feeding Habit for the Theropod Dinosaur Troodon. Gaia number 15. December 1998. pp. 159–166
- (en) Jacobsen, A.R. 2001. Tooth-marked small theropod bone: An extremely rare trace. p. 58-63. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press
- (en) Horner, John R. (1984) "The nesting behavior of dinosaurs". "Scientific American", 250:130–137
- (en) David J. Varricchio, John J. Horner et Frankie D. Jackson, « Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus », Journal of Vertebrate Paleontology, vol. 22, no 3, , p. 564–576 (DOI 10.1671/0272-4634(2002)022[0564:EAEFTC]2.0.CO;2)
- (en) Horner, John R., Weishampel, David B. (1996) "A comparative embryological study of two ornithischian dinosaurs – a correction." "Nature" 383:256–257
- (en) D.J. Varricchio, F. Jackson, J.J. Borkowski et J.R. Horner, « Nest and egg clutches of the dinosaur Troodon formosus and the evolution of avian reproductive traits », Nature, vol. 385, no 6613, , p. 247–250 (DOI 10.1038/385247a0, Bibcode 1997Natur.385..247V)
- (en) D. J. Varricchio, J. R. Moore, G. M. Erickson, M. A. Norell, F. D. Jackson et J. J. Borkowski, « Avian Paternal Care Had Dinosaur Origin », Science, vol. 322, no 5909, , p. 1826–8 (PMID 19095938, DOI 10.1126/science.1163245, Bibcode 2008Sci...322.1826V)
- (en) Birchard Geoffrey F., Ruta Marcello et Deeming D. Charles, « Evolution of parental incubation behaviour in dinosaurs cannot be inferred from clutch mass in birds », Biology Letters, vol. 9, no 4, , p. 20130036 (PMID 23676654, PMCID 3730617, DOI 10.1098/rsbl.2013.0036, lire en ligne)