Protéine de transfert de lipides
Les protéines de transfert de lipides (ou PTL ou LTP pour les anglophones) sont un groupe de protéines produites par les animaux, les végétaux et certains microorganismes pathogènes (ex : élicitines sécrétées par Phytophthora et Pythium)[1]).
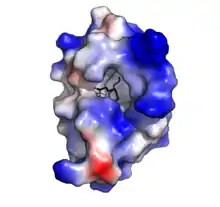
Comme l'indique leur nom, les protéines de transfert lipidique « embarquent » et « véhiculent » des phospholipides et d'autres groupes d'acides gras, en les transférant d'une membrane à une autre au sein de la cellule vers un point où la molécule transportée peut éventuellement alors traverser la membrane cellulaire[2].
Ces protéines PTL jouent un rôle majeur et beaucoup plus large chez les végétaux dits supérieurs (Tracheophyta) que chez les animaux.
Dans les cellules des végétaux, ce sont des protéines hautement conservées, d'environ 7 à 9 kDa, présentes dans de nombreux tissus [3], y compris dans le grain de pollen, sous sa cuticule protectrice[4] - [5]. Chez les plantes on subdivise les PTL en deux grandes sous-familles structurellement liées, selon leurs masses moléculaires LTP1 (9 kDa) et LTP2 (7 kDa)[6]. Différentes PTL se lient à un large éventail de ligands, dont des acides gras à longue de chaîne (C 10 à C 18), des dérivés acylés du coenzyme A (CoA), des phospholipides et galactolipides, la prostaglandine B 2, des stérols, des molécules de solvants organiques, et même divers polluants ou médicaments [5] PTL2.
Les PTL dans l'Histoire des sciences
La description des premières protéines de transfert de lipides (LTP) a été faite en 1975 par l'équipe de Jean-Claude Kader[7] qui montrera in vitro en 1996 que des phospholipides sont ainsi transportés d'une membrane donneuse à une membrane acceptrice[8].
Classification des PTL
Chez les végétaux ces petites protéines sont habituellement classées en 2 groupes :
- Le groupe 1 (ou PTL 1 ou LPT 1) : C'est celui des PTL de poids moléculaires de 10kDa environ ; Elles sont basiques, car leur point isoélectrique se situe entre 9 et 10 (Les acides aminés chargés, très abondants, sont très souvent conservés au sein des LTP 1 (Aspartate, Arginine et Lysine) notent Castro et al. en 2003[9].
Elles sont faites de 92 acides aminés environ (de 90 à 95). Elles sont caractérisées par la position, très conservée, de 8 résidus cystéine, qui forment 4 ponts disulfure, ce qui entraîne une très bonne stabilisation de la structure tertiaire[8].
Leur structure secondaire se compose de 4 hélices a mais aussi d'une longue région C-terminale (queue) apparemment peu ou pas structurée [10] - [11]. Leur structure tridimensionnelle a été étudiées par diffraction aux rayons-X et par Résonance Magnétique Nucléaire (RMN), montrant un repliement globulaire, stabilisé par les ponts-disulfures[10] - [11]. C'est dans ce repliement que les lipides sont transportés. Des LTP 1 ont notamment été trouvées et étudiées sur les parois cellulaires de différentes plantes dont la plante-modèle Arabidopsis thaliana[12], ou le ricin (Ricinus communis[13]) ;
- Le groupe 2 (ou PTL 2 ou LPT 2) : il rassemble des PTL plus courtes (70 acides aminés (7kDa) environ) mais dotées comme dans le groupe 1 d'un pI élevé, d'une activité de transfert de lipides, et de 4 ponts disulfure conservés)[9] - [14] - [15]. Elles ont le même profil de structure secondaire que les LTP 1, mais s'en différencient par le fait qu'elles ne forment que 3 hélices a et qu'elles présentent une région à 2 hélices plus courtes. La région C-terminale ne présente comme chez les PTL 1 aucune structuration particulière[14] - [16].
Description des PTL
Elle diffère pour chacun des deux groupes :
- Les PTL 1 disposent dans leur région centrale d'une cavité hydrophobe flexible formant un minuscule tunnel à 2 entrées (une petite et une large). Il est hydrophobe de ce tunnel parce que constitué de plusieurs résidus à chaînes latérales hydrophobes (notamment alanine, isoleucine, leucine, proline et valine)[10] - [11]. Deux acides aminés électropositifs (arginine et lysine) sont situés au voisinage de la plus grande entrée, interagissant avec les parties hydrophiles des lipides. La partie hydrophile des acides gras reste exposée au solvant et leur partie hydrophobe est aspirée dans le tunnel.
- Chez les PTL 2, la cavité hydrophobe n'a pas la forme d'un tunnel, mais d'une cavité de section globalement triangulaire, dont les parois sont des acides aminés hydrophobe : alanine, cystéine, isoleucine, leucine, phénylalanine et valine[16]. Par modélisation moléculaire on voit que cette cavité pourrait être plus flexible que celle en tunnel des LTP 1, ce que confirme l'association de stérols avec des LTP 2 plutôt qu'avec des LTP 1[17] - [16].
En N-terminal, un peptide signal est présent dans les 2 familles. Dans le groupe 1, sa longueur varie de 21 à 27 acides aminés P 1 [18] - [19], contre 27 à 35 acides aminés dans le groupe 2 [20] - [21]. Quand le peptide signal est excisé, la LTP mature est exportée vers l'apoplaste.
Dans les deux groupes, les protéines PTL présente une certaines élasticité permettant d'adapter le volume de la cavité à différentes tailles de phospholipides (il n'y a pas de spécificité dans les interactions) ce pourquoi les LTP sont aussi dénommées Protéines de Transfert de Lipides non spécifiques (nsLTP)[22] - [16].
Diversité des PTL
Chez les végétaux, elle varie selon les espèces ou essences, mais elle est toujours élevée (ex : 12 PTL connues dans le riz Oryza sativa[10], 11 trouvées dans le haricot mungo Vigna radiata var. radiate[23], 11 LTP 1 et 13 LTP 2 dans le mil Sorghum vulgare[24] et 11 LTP 1 dans le colza Brassica napus[10], et chacune de ces protéines peut transporter plusieurs types de molécules lipidiques.
Activité biologique chez les végétaux
Chez le végétaux, les protéines de transfert de lipides forment l'une des classes très importantes de défense PRPS, dont beaucoup ont des activités antimicrobiennes et enzymatiques, ou sont des inhibiteurs d'enzymes. Différentes protéines de cette classe peuvent présenter les activités suivantes[5] :
- antibactérien ;
- antifongique ;
- antiviral ;
- antiprolifératif.
Fonctions des PTL
Elles sont probablement encore incomplètement connues. On sait avec certitude depuis les années 1990 que chez les plantes une même protéine PTL peut avoir des fonctions différentes (car dans la cellule, plusieurs gènes codent des LTP[19] - [24] - [18]). Étudier ce qui déclenche l'expression de chacun de ces gènes, seuls ou combinés entre eux ou à d'autres gènes pourra éclairer les rôles précis de ces protéines mais cela demandera un important travail[25]
Le premier rôle à avoir été mis en évidence chez les végétaux est le transport des lipides de la membrane plasmique et des membranes des organelles, lipides qui ont un rôle dans de nombreux processus (catabolisme, transport de molécules, et métabolisme cellulaire). Ainsi le glyoxysome est un organite intracellulaire dont la membrane et faite de 3 types de lipides (phosphatidylcholine, phosphatidylglycérol et de phosphatidyléthanolamine) qu'il ne peut lui-même fabriquer, faute de la machine enzymatique permettant la synthèse de ces lipides. Ces trois lipides sont donc importés à partir du réticulum endoplasmique où ils ont été synthétisés. De même les membranes des chloroplastes et des mitochondries sont faites de lipides importés[26] ; ceci implique un système de transport intracellulaire spécifique, dédié aux lipides , en effet la plupart des lipides ne peuvent pas spontanément quitter une cellule ou y entrer en traversant sa membrane car leur hydrophobicité les rend peu solubles dans l'eau.
Les PTL permettent ou facilitent le mouvement des lipides entre cellules au travers des membranes en se liant et en les solubilisant. Chaque LTP est généralement spécifique d'un type de substrat, mais peut interagir avec une large variété de lipides [27], ce qui leur confère des fonctions élargies :
Chez les plantes supérieurs les PTL peuvent ainsi être impliqués dans :
- le métabolisme, transport et catabolisme des lipides ;
- la biosynthèse de la cutine ;
- la biosynthèse de cire protectrice, en surface de la cuticule foliaire ;
- la croissance mitochondriale ;
- les réactions de défense des plantes face à de nombreux agents pathogènes[1] ;
- l'adaptation aux changements environnementaux [28] - [1] ;
- la fertilisation des plantes à fleurs (on les retrouve dans le pistil et le grain de pollen, sous la cuticule protectrice) ;
- l'adaptation des plantes à certains facteurs de stress (stress induits par leurs pathogènes [1]) ;
- l'activation et la régulation de cascades de signalisation ;
- l'apoptose (suicide cellulaire);
- certaines symbioses ;
- la maturation des fruits[5]...
Structure des PTL
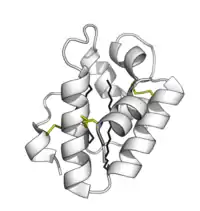
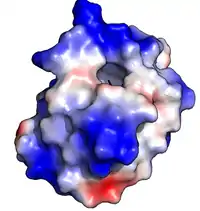
Les protéines de transfert des lipides sont chez les végétaux de petite taille (10 kDa) et composées d’environ 90 acides aminés, formant 4 hélices alpha (α) insérées dans une super hélice droitière à topologie de feuille pliée qui a grossièrement la forme compacte repliée d'un petit saxophone. « Quatre ponts disulfure conservés qui relient les différentes régions de la chaîne polypeptidique assurant à ces protéines une résistance remarquable à la dénaturation thermique (cuisson) et à l’attaque des protéases digestives »[3]. Chacune de ces protéines forme une structure présentant une cavité hydrophobe interne où 1 à 2 lipides peuvent être liés et transportés. La surface externe de la protéine étant hydrophile, le complexe "protéine + lipide" devient soluble.Des interactions hydrophobes, et très peu d'interactions chargées, permettent à la protéine d'accepter une large gamme de lipides[27].
Résistance des PTL à la chaleur, au froid, à la digestion
Cette résistance a été testée à la toute fin du XXe siècle et dans les années suivantes : on savait déjà que ces protéines résistent à la congélation et assez bien à la cuisson (un chauffage à plus de 100 °C, ne détruit pas assez les LPTL de pêche et de maïs pour empêcher leur interaction avec les IgE de patients allergiques[29] - [30] - [31].
Introduites dans un environnement gastrique simulé (mise en contact avec la pepsine et d'autres enzymes digestives dans un environnement aussi acide que celui d'un estomac), elles sont encore intacts après 2 heures et seul 40% de l'allergène Pru p 3 avait été dégradé en 24 h[32] - [33] - [34] - [35].
Cette résistance explique que des jus de fruits (même pasteurisés par un choc thermique), des bières, du vin ou des confitures restent allergènes pour certaines personnes, bien que contenant des restes de fruits ou graines/malt fortement dénaturés[29] - [32] - [35] - [36] ; ils peuvent encore chez des personnes sensibilisées, causer des allergies aux PTL, qui vont d'une simple simple rhinite allergique à des cas plus sévères par exemple observées dans les pays du pourtour méditerranéen (urticaire généralisée, asthme, œdème, choc anaphylactique).
Niveaux d'expression des PTL
Ces niveaux varient selon l'espèce et selon le tissu et selon le stade de développement, mais aussi selon les conditions physiologiques et environnementales [18] - [37] - [38].
Par exemple le froid ou le stress salin modifient les profils d'expression de diverses LTP chez la plante ; de même quand elle est infectée par des bactéries ou champignons phytopathogènes[39] - [40].
Les molécules régulatrices pourraient notamment être l'acide abscissique, l'acide salicylique, l'éthylène (gaz) ou le jasmonate qui semblent impliqués dans les voies de signalisation activant l'expression des gènes de LTP [39] - [40] - [41] - [42]. Ce rôle a été confirmé par le génie génétique : par exemple l'introduction de LTP d'orge dans des sujets transgéniques d'arabette et de tabac (Nicotiana tabacum) a accru la tolérance à l'infection par un pathogène. Ce tabac transgénique retardait notamment la croissance de la bactérie Pseudomonas syringae pv. tabaci, tout en réduisant fortement le nombre de nécroses quantifiées chez les plantes infectées. De même A. thaliana résistait mieux à la bactérie P. syringae pv. tomato et aux attaques de champignon Botrytis cinerea[43]). Ces expériences intéressent beaucoup les partisans du génie génétique dans l'agroalimentaire, mais posent la question des risques de transferts d'allergénicité lors de transferts de gènes de résistances aux maladies impliquant des PTL.
Les PTL ont été classés au sein de la grande famille des protéines PR-14, en raison de leur activité antimicrobienne, leurs similarités de séquence et leur induction à la suite d'une infection par un phyto-pathogène. Certaines PTL ont une activité néanmoins très spécifique (ainsi la LTP purifiée du riz, exprimée chez E. coli est active contre la champignon Pyricularia oryzae mais pas du tout contre Xanthomonas oryzae[44].
En 1996, Kader postule que les LTP interagissent négativement avec le pathogène en perméabilisant sa membrane, ce qui a ensuite été confirmé par des expériences en laboratoire in vitro sur des membranes-modèles ou sur des levures vivantes[45] - [46] mais les mécanisme précis d'action anti-microbienne des LTP sont encore à éclaircir.
Localisation dans l'organisme
Chez la plante, les PTL semblaient toujours extracellulaires, mais il existe des contre-exemples, par exemple
- chez le ricin, où une isoforme de LTP a été retrouvée dans le glyoxysome, ce qui lui permettrait d'activer l'acétyl-CoA oxydase in vitro, suggérant une possible implication des LTP dans la b - oxydation et la régulation du transport et du stockage des lipides[13] ;
- dans le grain de blé où les vacuoles de stockage des graines de Niébé (Vigna unguiculata) en contiennent, de même que les grains d'aleurone contenus dans les grains de blé (alors que les LTP, en 1998 n'avaient toujours pas été retrouvées dans les parois cellulaires qui ont pourtant aussi besoin d'être défendues)[47] mais le rôle physiologique exact n'était en 2007 pas encore été expliqué[25].
l'expression du gène de la LTP 1 varie dans le temps et des LTP jouent parfois un rôle en surface de la feuille. Sur les jeunes feuilles de chou rouge (Brassica oleracea), les LTP constituent près de 50% des protéines présentes sur la cuticule cireuse des jeunes feuilles alors qu'elles ne sont que plus que 4% dans la feuille mature ; on suppose qu'elles transportent en surface le monomère de la cutine, lors de la croissance foliaire[48]. A. thaliana en produit également plus dans les tissus jeunes, sans doute aussi pour répondre au besoin de cutine nécessaire à la croissance foliaire et d'autres organes de la plante[49]. On a montré en 1991 que la PTL transporte bien la cutine dans ce cas pour former la membrane lipophilique qui protège les cellules embryonnaires[50] - [51]
Rôle dans les voies de signalisation des plantes ?
C'est une hypothèse défendue par plusieurs auteurs [52] - [53], qui reste à expliciter, mais qui a été suscitée par plusieurs observations :
- Chez A. thaliana, un gène (dir1-1, très proche de celui des PTL) joue un rôle dans la transmission du signal intracellulaire ; Ce gène code une protéine de 102 acides aminés, laquelle interagi avec des lipides pour produire un complexe impliqué dans la résistance systémique des plantes face à leur spathogènes[54] ;
- Chez le Tabac, on observe (in vitro) aussi une interaction de la LTP de blé avec des récepteurs des élicitines[55] qui sont de petites protéines (+/-10kDa) produites par des Oomycètes du genre Phytophthora ou Pythium ; introduites chez le tabac, elles y induisent une réaction d'hypersensibilité[56]. Or, à la manière des PTL, les élicitines sont de petites protéines basiques possédant des ponts disulfure et une structure secondaire constituée surtout d'hélices a, qui forment également une poche hydrophobe permettant de capter des molécules hydrophobes (stérols notamment) ;
- une LTP recombinante de tabac fixant l'acide jasmonique forme un complexe reconnu par le récepteur des élicitines[57]. Une fois infiltré dans des plants de tabacs, ce complexe y crée une protection systémique (c'est-à-dire de toutes les parties de la plante) contre plusieurs agents pathogènes (dont P. parasitica) ; en 2004, on ignore cependant si ce complexe est le signal d'activation, ou s'il ne fait qu'indirectement activer une voie de signalisation encore à découvrir.
Autres protéines apparentées
Les protéines de transfert des lipides des plantes partagent le même domaine structural [58] que les protéines de stockage des graines [59] et les inhibiteurs de la trypsine alpha-amylase[60] - [61] Ces protéines partagent le même faisceau de quatre hélices stabilisé au disulfure superhélique contenant une cavité interne.
Il n'y a pas de similarité de séquence entre les PTL d'origine animale et végétale. Chez les animaux, la protéine de transfert du cholestérol (LTC), également appelée protéine de transfert des lipides plasmatiques, est une protéine plasmatique facilitant le transport des esters et des triglycérides du cholestérol entre les lipoprotéines.
Interactions avec la santé humaine
De nombreuses protéines de défense biosynthétisées par les plantes sont sensibilisantes et/ou allergisantes : c'est le cas par exemple des chitinases, des protéines Bet v 1-like mais aussi des protéines de transfert lipidique PTL. Leur impact allergique et l'effet de certains procédés agricoles ou technoalimentaires sur l'allergénicité des aliments végétaux font l'objet d'études.
Allergénicité
De nombreux allergènes de type PTL peuvent, peu à peu, après ingestion, provoquer des manifestations de sensibilisation puis de réactions allergiques, ce qui les a fait classer parmi les allergènes alimentaires de classe [6].
Leur allergénicité est induite par « trois épitopes liant les IgE exposés en surface. L’épitope 2 est un épitope consensus conservé dans de nombreuses LTP de fruits (ex raisin), du latex (Hev b 9) ou de graines (Cor a 8 de la noisette) »[3].
Chez des personnes allergiques (ou en cours de sensibilisation allergique), ces protéines (extrêmement répandues chez le monde des végétaux, dans les racines, graines, fruits, pollens, feuilles) peuvent devenir de puissants panallergènes alimentaires[62]. Le cas le plus connu est la PTL de la pêche (Pru p 3). Cette « communauté épitopique » explique les fréquentes réactivités croisées entre des PTL d’origine très différente. Elle explique aussi pourquoi des personnes allergiques à la pêche ont un profil si diffèrent selon qu'ils vivent dans le Nord ou le Sud d'un continent tel que l’Europe : un espagnol risque de se sensibiliser à la pêche en ingérant directement la PTL (Pru p 3) de ce fruit avec des réactions allergiques généralisées, alors que plus au nord un néerlandais ou un autrichien se sensibiliseront à la pêche via des contacts répétés et allergisant avec le Bet v 1, en inhalant par exemple des pollens dégradés ou pollués de bouleau et en développant une allergie à la pêche par réaction croisée entre Bet v 1 et son homologue Mal d 1 synthétisé par le pommier et retrouvé dans la pomme[3].
Ces protéines sont souvent impliquées dans les cas de sensibilisation et d'allergies croisée à des éléments aussi différents qu'un pollen de bouleau, du raison, du latex ou des noisettes. Elles peuvent expliquer le développement d'allergies graves au pollen à la suite d'une exposition allergénisante à du latex ou à un aliment végétal, et inversement. Pour ces raisons, les manifestations cliniques peuvent concerner les voies respiratoire, digestive et/ou cutanées [63], l'allergie pouvant éventuellement être d'origine professionnelles (travail sous-serre, jardinier, travail dans l'industrie agroalimentaire..).
Parce que thermostables et résistantes à la protéolyse gastro-intestinale (c'est-à-dire très résistantes à la digestion) [64], chez les personnes sensibles ou sensibilisées, elles causent des réactions allergiques bénignes à graves. En zone méditerranéennes, ces protéines sont les phytoallergènes majeurs, en cause dans les allergies alimentaires aux rosacées et cause de rhume des foins en présence de pollen d'espèces de Parietaria (Pariétaire officinale, principalement). La voie qui initie l'allergie semble le plus souvent alimentaire.
La diversité des aliments impliqués, les risques d'allergies croisées, la fréquence des sensibilisations asymptomatiques et l’importance des cofacteurs sont des éléments à prendre en compte dans la démarche diagnostique[65].
Usage médicinal
Chez des patients qui n'y sont pas allergiques, ils peuvent être utilisés comme antioxydants, et préviennent certaines maladies[66].
Importance commerciale
La protéine de transfert lipidique 1 (de l’orge) est responsable, lorsqu’elle est dénaturée par le processus de brassage, de la majeure partie de la mousse qui se forme sur la bière[67].
Voir aussi
Articles connexes
- Mousse de bière
- Allergie
Bibliographie
- Borges J.P (2008) Caractérisation structurale et immunologique d'allergènes alimentaires: les protéines de transfert de lipides de fruits (Doctoral dissertation, Immunologie, Université de Toulouse, Université Toulouse III-Paul Sabatier). PDF, 208 p|URL =http://thesesups.ups-tlse.fr/209/1/Borges_Jean-Philippe.pdf
- Malandain H & Lavaud F (2004) Allergénicité des protéines de défense végétale. Revue française d'allergologie et d'immunologie clinique, 44(5), 469-475 (résumé).
Références
- Buhot, N. (2003). Rôle des élicitines et des protéines de transfert de lipides dans l'induction de la résistance des plantes à leurs agents pathogènes (Doctoral dissertation, Dijon).
- « Lipid-Transfer Protein in Plants », Annual Review of Plant Physiology and Plant Molecular Biology, vol. 47, , p. 627–654 (PMID 15012303, DOI 10.1146/annurev.arplant.47.1.627)
- Rougé, P., Borges, J. P., Culerrier, R., Brulé, C., Didier, A., & Barre, A. (2009) Les protéines de transfert des lipides : des allergènes importants des fruits. Revue Française d'Allergologie, 49(2), 58-61 |(résumé)
- « Lipid transfer protein: a pan-allergen in plant-derived foods that is highly resistant to pepsin digestion », International Archives of Allergy and Immunology, vol. 124, nos 1–3, , p. 67–9 (PMID 11306929, DOI 10.1159/000053671)
- « Lipid Transfer Proteins As Components of the Plant Innate Immune System: Structure, Functions, and Applications », Acta Naturae, vol. 8, no 2, , p. 47–61 (PMID 27437139, PMCID 4947988)
- « Plant Pathogenesis-Related Proteins PR-10 and PR-14 as Components of Innate Immunity System and Ubiquitous Allergens », Current Medicinal Chemistry, vol. 24, no 17, , p. 1772–1787 (PMID 27784212, DOI 10.2174/0929867323666161026154111)
- Kader JC (1975) Proteins and the intracellular exchange of lipids. I. Stimulation of phospholipid exchange between mitochondria and microsomal fractions by proteins isolated from potato tuber. Biochim Biophys Acta 380, 31-44.
- Kader J (1996) Lipid-transfer proteins in plants. Annu Rev Plant Physiol Plant Mol Biol 47, 627-654
- Castro MS, Gerhardt IR, Orrù S, Pucci P and Bloch CJ (2003). Purification and characterization of a small (7.3 kDa) putative lipid transfer protein from maize seeds. J Chromatogr B Analyt Technol Biomed Life Sci 794, 109-114.
- Lee JY, Min K, Cha H, Shin DH, Hwang KY and Suh SW (1998). Rice non-specific lipid transfer protein: the 1.6 A crystal structure in the unliganded state reveals a small hydrophobic cavity. J Mol Biol 276, 437-448
- Shin DH, Lee JY, Hwang KY, Kim KK and Suh SW (1995). High-resolution crystal structure of the non-specific lipid-transfer protein from maize seedlings. Structure 3, 189-199
- Thoma S, Kaneko Y and Somerville C (1993). A non-specific lipid transfer protein from Arabidopsis is a cell wall protein. Plant J 3, 427-436
- Tsuboi S, Osafune T, Tsugeki R, Nishimura M and Yamada M (1992). Nonspecific lipid transfer protein in castor bean cotyledon cells: subcellular localization and a possible role in lipid metabolism. J Biochem (Tokyo) 111, 500-508
- Douliez JP, Pato C, Rabesona H, Mollé D and Marion D (2001). Disulfide bond assignment, lipid transfer activity and secondary structure of a 7-kDa plant lipid transfer protein, LTP2. Eur J Biochem 268, 1400-1403.
- Liu Y, Samuel D, Lin C and Lyu P (2002). Purification and characterization of a novel 7-kDa non-specific lipid transfer protein-2 from rice (Oryza sativa). Biochem Biophys Res Commun 294, 535-540.
- Samuel D, Liu Y, Cheng C and Lyu P (2002). Solution structure of plant nonspecific lipid transfer protein-2 from rice (Oryza sativa). J Biol Chem 277, 35267-35273
- Buhot N, Gomès E, Milat M, Ponchet M, Marion D, Lequeu J, Delrot S, Coutos-Thévenot P and Blein J (2004). Modulation of the biological activity of a tobacco LTP1 by lipid complexation. Mol Biol Cell 15, 5047-5052
- Arondel V, Tchang F, Baillet B, Vignols F, Grellet F, Delseny M, Kader JC and Puigdomènech P (1991). Multiple mRNA coding for phospholipid-transfer protein from Zea mays arise from alternative splicing. Gene 99, 133-136.
- Vignols F, Lund G, Pammi S, Trémousaygue D, Grellet F, Kader JC, Puigdomènech P and Delseny M (1994). Characterization of a rice gene coding for a lipid transfer protein. Gene 142, 265-270
- García-Garrido JM, Menossi M, Puigdoménech P, Martínez-Izquierdo JA and Delseny M (1998). Characterization of a gene encoding an abscisic acid-inducible type-2 lipid transfer protein from rice. FEBS Lett 428 , 193-199
- Kalla R, Shimamoto K, Potter R, Nielsen PS, Linnestad C and Olsen OA (1994). The promoter of the barley aleurone-specific gene encoding a putative 7 kDa lipid transfer protein confers aleurone cell-specific expression in transgenic rice. Plant J 6, 849-860.
- Han GW, Lee JY, Song HK, Chang C, Min K, Moon J, Shin DH, Kopka ML, Sawaya MR, Yuan HS, Kim TD, Choe J, Lim D, Moon HJ and Suh SW (2001). Structural basis of non-specific lipid binding in maize lipid-transfer protein complexes revealed by high-resolution X-ray crystallography. J Mol Biol 308, 263-278.
- Lin K, Liu Y, Hsu SD, Samuel D, Cheng C, Bonvin AMJJ and Lyu P (2005) Characterization and structural analyses of nonspecific lipid transfer protein 1 from mung bean. Biochemistry 44, 5703-5712
- Pelèse-Siebenbourg F, Caelles C, Kader JC, Delseny M and Puigdomènech P (1994). A pair of genes coding for lipid-transfer proteins in Sorghum vulgare. Gene 148, 305-308.
- Carvalho ADO and Gomes VM (2007). Role of plant lipid ransfer proteins in plant cell physiology-a concise review. Peptides 28, 1144-1153.
- Moreau P, Bessoule JJ, Mongrand S, Testet E, Vincent P and Cassagne C (1998). Lipid trafficking in plant cells. Prog Lipid Res 37, 371-391.
- « Lipid binding in rice nonspecific lipid transfer protein-1 complexes from Oryza sativa », Protein Science, vol. 13, no 9, , p. 2304–15 (PMID 15295114, PMCID 2280015, DOI 10.1110/ps.04799704)
- Kader, « Science Direct », Trends in Plant Science, vol. 2, no 2, , p. 66–70 (DOI 10.1016/S1360-1385(97)82565-4, lire en ligne)
- Brenna OV, Pastorello EA, Farioli L, Pravettoni V and Pompei C (2004) Presence of allergenic proteins in different peach (Prunus persica) cultivars and dependence of their content on fruit ripening. J Agric Food Chem 52, 7997-8000
- Lindorff-Larsen K and Winther JR (2001) Surprisingly high stability of barley lipid transfer protein, LTP1, towards denaturant, heat and proteases. FEBS Lett 488, 145-148
- Pastorello EA, Farioli L, Pravettoni V, Ortolani C, Fortunato D, Giuffrida MG, Perono Garoffo L, Calamari AM, Brenna O and Conti A (2003). Identification of grape and wine allergens as an endochitinase 4, a lipid-transfer protein, and a thaumatin. J Allergy Clin Immunol 111, 350-359
- Duffort OA, Polo F, Lombardero M, Díaz-Perales A, Sánchez-Monge R, García-Casado G, Salcedo G and Barber D (2002). Immunoassay to quantify the major peach allergen Pru p 3 in foodstuffs. Differential allergen release and stability under physiological conditions. J Agric Food Chem 50, 7738-7741.
- Vassilopoulou E, Rigby N, Moreno FJ, Zuidmeer L, Akkerdaas J, Tassios I, Papadopoulos NG, Saxoni-Papageorgiou P, van Ree R and Mills C (2006). Effect of in vitro gastric and duodenal digestion on the allergenicity of grape lipid transfer protein. J Allergy Clin Immunol 118, 473-480.
- Asero R, Mistrello G, Roncarolo D, de Vries SC, Gautier MF, Ciurana CL, Verbeek E, Mohammadi T, Knul-Brettlova V, Akkerdaas JH, Bulder I, Aalberse RC and van Ree R (2000). Lipid transfer protein: a pan-allergen in plant-derived foods that is highly resistant to pepsin digestion. Int Arch Allergy Immunol 122, 20-32
- Scheurer S, Lauer I, Foetisch K, San Miguel Moncin M, Retzek M, Hartz C, Enrique E, Lidholm J, Cistero-Bahima A and Vieths S (2004). Strong allergenicity of Pru av 3, the lipid transfer protein from cherry, is related to high stability against thermal processing and digestion. J Allergy Clin Immunol 114, 900-907.
- García-Casado G, Crespo JF, Rodríguez J and Salcedo G (2001). Isolation and characterization of barley lipid transfer protein and protein Z as beer allergens. J Allergy Clin Immunol 108, 647-649.
- Vignols F, Wigger M, García-Garrido JM, Grellet F, Kader JC and Delseny M (1997) Rice lipid transfer protein (LTP) genes belong to a complex multigene family and are differentially regulated. Gene 195, 177-186
- Soufleri IA, Vergnolle C, Miginiac E and Kader JC (1996). Germination-specific lipid transfer protein cDNAs in Brassica napus| L. Planta 199, 229-237
- Jang CS, Lee HJ, Chang SJ and Seo YW (2004). Expression and promoter analysis of the TaLTP1 gene induced by drought and salt stress in wheat (Triticum aestivum L.). Plant Science 167, 995-1001
- Jung HW, Kim W and Hwang BK (2003) Three pathogen-inducible genes encoding lipid transfer protein from pepper are differentially activated by pathogens, abiotic, and environmental stresses. Plant Cell Environ 26, 915-928.
- Jung HW, Kim KD and Hwang BK (2005). Identification of pathogen-responsive regions in the promoter of a pepper lipid transfer protein gene (CALTPI) and the enhanced resistance of the CALTPI transgenic Arabidopsis against pathogen and environmental stresses. Planta 221,361-373.
- Jung HW, Lim CW, Lee SC, Choi HW, Hwang CH and Hwang BK (2007). Distinct roles of the pepper hypersensitive induced reaction protein gene CaHIR1 in disease and osmotic stress, as determined by comparative transcriptome and proteome analyses. Planta
- Molina A and García-Olmedo F (1997). Enhanced tolerance to bacterial pathogens caused by the transgenic expression of barley lipid transfer protein LTP2. Plant J 12, 669-675.
- Ge X, Chen J, Li N, Lin Y, Sun C and Cao K (2003). Resistance function of rice lipid transfer protein LTP110. J Biochem Mol Biol 36, 603-607.
- Subirade M, Salesse C, Marion D and Pézolet M (1995). Interaction of a nonspecific wheat lipid transfer protein with phospholipid monolayers imaged by fluorescence microscopy and studied by infrared spectroscopy. Biophys J 69, 974-988.
- Diz MSS, Carvalho AO, Rodrigues R, Neves-Ferreira AGC, Da Cunha M, Alves EW, Okorokova-Façanha AL, Oliveira MA, Perales J, Machado OLT and Gomes VM (2006). Antimicrobial peptides from chili pepper seeds causes yeast plasma membrane permeabilization and inhibits the acidification of the medium by yeast cells. Biochim Biophys Acta 1760, 1323-1332
- Dubreil L, Gaborit T, Bouchet B, Gallant DJ, Broekaert WF, Quillien L and Marion D (1998). Spatial and temporal distribution of the major isoforms of puroindolines (puroindoline-a and puroindoline-b) and non specific lipid transfer protein (ns-LTP1e1) of Triticum aestivum seeds. Relationships with their in vitro antifungal properties. Plant Science 138(2), 121-135.
- Pyee J, Yu H and Kolattukudy PE (1994). Identification of a lipid transfer protein as the major protein in the surface wax of broccoli (Brassica oleracea) leaves. Arch Biochem Biophys 311, 460-468
- Thoma S, Hecht U, Kippers A, Botella J, De Vries S and Somerville C (1994). Tissue-specific expression of a gene encoding a cell wall-localized lipid transfer protein from Arabidopsis. Plant Physiol 105, 35-45
- Cheng C, Samuel D, Liu Y, Shyu J, Lai S, Lin K and Lyu P (2004). Binding mechanism of nonspecific lipid transfer proteins and their role in plant defense. Biochemistry 43, 13628-13636.
- Sterk P, Booij H, Schellekens GA, Van Kammen A and De Vries SC (1991). Cell-specific expression of the carrot EP2 lipid transfer protein gene. Plant Cell 3, 907-921.
- Blein J, Coutos-Thévenot P, Marion D and Ponchet M (2002). From elicitins to lipid-transfer proteins: a new insight in cell signalling involved in plant defence mechanisms. Trends Plant Sci 7, 293-296.
- Mikes V, Milat ML, Ponchet M, Panabières F, Ricci P and Blein JP (1998). Elicitins, proteinaceous elicitors of plant defense, are a new class of sterol carrier proteins. Biochem Biophys Res Commun 245, 133-139.
- Maldonado AM, Doerner P, Dixon RA, Lamb CJ and Cameron RK (2002). A putative lipid transfer protein involved in systemic resistance signalling in Arabidopsis. Nature 419, 399-403.
- Buhot N, Douliez JP, Jacquemard A, Marion D, Tran V, Maume BF, Milat ML, Ponchet M, Mikès V, Kader JC and Blein JP (2001). A lipid transfer protein binds to a receptor involved in the control of plant defence responses. FEBS Lett 509, 27-30
- Ponchet M, Panabières F, Milat M-L, Mikes V, Montillet JL, Suty L, Triantaphylides C, Tirilly Y and Blein JP (1999). Are elicitins cryptograms in plant-Oomycete communications?. Cell Mol Life Sci 56, 1020-1047.
- Buhot N, Gomès E, Milat M, Ponchet M, Marion D, Lequeu J, Delrot S, Coutos-Thévenot P and Blein J (2004). Modulation of the biological activity of a tobacco LTP1 by lipid complexation. Mol Biol Cell 15, 5047-5052.
- « Characterization and structural analyses of nonspecific lipid transfer protein 1 from mung bean », Biochemistry, vol. 44, no 15, , p. 5703–12 (PMID 15823028, DOI 10.1021/bi047608v)
- « Solution structure of RicC3, a 2S albumin storage protein from Ricinus communis », Biochemistry, vol. 42, no 47, , p. 13839–47 (PMID 14636051, DOI 10.1021/bi0352217)
- « Tertiary and quaternary structures of 0.19 alpha-amylase inhibitor from wheat kernel determined by X-ray analysis at 2.06 A resolution », Biochemistry, vol. 36, no 44, , p. 13503–11 (PMID 9354618, DOI 10.1021/bi971307m)
- « Structure of the bifunctional inhibitor of trypsin and alpha-amylase from ragi seeds at 2.2 A resolution », Acta Crystallographica D, vol. 56, no Pt 3, , p. 287–93 (PMID 10713515, DOI 10.1107/s0907444999016601)
- Morris, « Food Allergy in Detail », Surrey Allergy Clinic
- Lavaud F, Fontaine, J. F., Perotin, J. M., Angelier, A. S., Meirhaeghe, D., & Lebargy, F. (2009). Manifestations cliniques de l’allergie aux protéines de transfert lipidique. Revue Française d'Allergologie, 49(5), 427-432.
- « A novel lipid transfer protein from the pea Pisum sativum: isolation, recombinant expression, solution structure, antifungal activity, lipid binding, and allergenic properties », BMC Plant Biology, vol. 16, , p. 107 (PMID 27137920, PMCID 4852415, DOI 10.1186/s12870-016-0792-6)
- Fontaine J-F (2015) Les protéines de transfert lipidique : actualités et implications cliniquesLipid transfer proteins: New and clinical implication ; Revue Française d'Allergologie ; Volume 55, Issue 3, April 2015, Pages 176-177 | https://doi.org/10.1016/j.reval.2015.01.025
- « Antioxidants in human health and disease », Annual Review of Nutrition, vol. 16, , p. 33–50 (PMID 8839918, DOI 10.1146/annurev.nu.16.070196.000341)
- « Foam », Carlsberg Research Laboratory