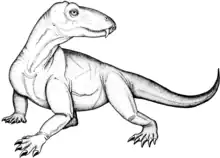
Pampaphoneus
Pampaphoneus biccai

Pampaphoneus est un genre éteint de reptiles carnivores de l'ordre des thérapsides et du sous-ordre des dinocéphales. Il appartient à la famille des Anteosauridae. Il vécut au Permien moyen (Wordien, entre 268 et 265 millions d’années environ), dans ce qui est aujourd’hui le Brésil. Le genre n’est représenté que par son espèce type, Pampaphoneus biccai, qui fut nommée en 2012 par Juan Carlos Cisneros (d) et des collègues à partir d’un crâne presque complet avec la mâchoire inférieure encore articulée, découvert sur les terres de la Ferme Boqueirão, près de la ville de São Gabriel, dans l’État du Rio Grande do Sul. Un second spécimen provenant de la même localité a été signalé en 2019 et en 2020 mais n’a pas encore été décrit. Il est composé d’un crâne associé à des restes postcrâniens. Pampaphoneus est le premier genre de dinocéphale décrit en Amérique du Sud. Le groupe n’y était connu auparavant que par quelques dents isolées et un fragment de mâchoire signalés en 2000 dans la même région du Brésil. L’analyse phylogénétique conduite par Cisneros et des collègues révèle que Pampaphoneus est étroitement apparenté à des antéosaures de Russie européenne ce qui indique une relation faunistique plus étroite entre l’Amérique du Sud et l’Europe de l'Est qu’on ne le pensait auparavant, favorisant ainsi une reconstruction continentale de type B de la Pangée.
Étymologie
Le nom générique Pampaphoneus est composé de « pampa », le nom de la grande plaine du sud du continent sud-américain, et du mot du grec ancien « phoneus », « tueur » pour donner « tueur de la pampa ». Le nom spécifique biccai rend hommage à José Bicca, propriétaire du terrain où le fossile fût trouvé[1].
Description
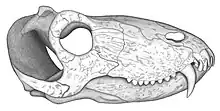
L’holotype (UFRGS PV386P) de Pampaphoneus est un crâne mesurant environ 32 centimètres de longueur. Comme chez tous les antéosaures, la marge ventrale du prémaxillaire est inclinée vers le haut et la barre postorbitaire est fortement courbée antéroventralement de telle sorte que la fosse temporale s’étend vers la région sous orbitaire. Le toit crânien est légèrement pachyostosé. Les fosses temporales mordent largement sur le toit crânien où la zone d’insertion de la musculature adductrice de la mandibule s’étend jusqu’à la bosse pinéale. Les frontaux contribuent à la zone d’insertion de cette musculature et participent également au bord antérieur de la bosse pinéale. Ces caractéristiques permettent d’attribuer Pampaphoneus au clade des Syodontinae. Au sein de ces derniers, c’est avec l’espèce Russe Syodon biarmicum que Pampaphoneus partage le plus de ressemblances. Comme ce dernier, il possède des canines très recourbées en forme de crochets et des postcanines très basses pourvues de serrassions. Les canines de l’holotype mesurent 7 cm de longueur. Pampaphoneus diffère toutefois de Syodon par sa plus grande taille, son museau plus robuste, ses postorbitaires épaissi formant une bosse orbitaire, et par une crête bien développée s’étendant de la bosse pinéale à la marge de l’orbite. Il diffère aussi des autres antéosaures par son squamosal pourvu d’un processus jugal dépassant le bord le plus antérieur de la fosse temporale[1]. Une particularité de l’holotype que l’on pensait être un caractère propre au genre est la présence de seulement quatre dents par prémaxillaire (la quatrième incisive, de petite taille, étant toutefois masquée latéralement par le maxillaire) et huit postcanines[1]. Cependant, un second crâne (encore non décrit) a été signalé montrant quelques différences avec l’holotype comme la présence de cinq dents prémaxillaires et seulement sept postcanines. Les autres éléments crâniens étant très similaires, ces différences sont interprétées comme relevant de la variation intraspécifique[2] - [3]. Bien qu’étant un Syodontinae, Pampaphoneus partage aussi plusieurs caractères avec les Anteosaurinae. Il possède ainsi une crête médiane bien développée sur le toit crânien, tandis qu’un épaississement prononcé de chaque postorbitaire forme une bosse similaire à celles d’un individu subadulte de Titanophoneus potens de Russie. A l’arrière de la mandibule, l’os angulaire porte une bosse comme chez Anteosaurus et les deux espèces de Titanophoneus. Cette bosse de l’angulaire est toutefois beaucoup moins développée chez Pampaphoneus[1].
Paléoécologie
Les fossiles de Pampaphoneus proviennent du Membre de Morro Pelado de la Formation de Rio do Rasto affleurant sur les terres de la ferme Boqueirão dans la municipalité de São Gabriel (district de Catuçaba, Etat du Rio Grande do Sul). Les fossiles y sont conservés dans un grès fin rosâtre et sont recouverts d’une croûte sombre d’oxyde de fer[4]. Les sédiments du Membre de Morro Pelado se sont déposés dans des milieux fluvio-lacustres et deltaïques suggérant des conditions alluviales avec des plaines d'inondations coalescentes. Des faciès éoliens (dunes fossiles) présents dans la partie supérieure du Membre de Morro Pelado attestent d’une aridification progressive de l’environnement. Paléogéographiquement, le sud-est du Brésil était situé aux latitudes moyennes comprises entre le 35e et le 45e parallèle sud où régnait probablement un climat de type tempéré chaud avec une saison sèche prolongée[5]. La flore comportait des forêts à Glossopteris communis qui occupaient les plaines inondables et les digues de chenaux tandis que les biotopes plus humides étaient recouverts de tapis denses de sphénopsides comprenant des équisetales (Schizoneura et Phyllotheca) et des sphénophylles (Sphenophyllum paranaense)[5]. Outre Pampaphoneus, le site de la ferme Boqueirão contient également le temnospondyle Konzhukovia sangabrielensis[6] et le petit dicynodonte Rastodon[4]. D’autres sites du Membre de Morro Pelado ont également livré des restes de tétrapodes. Toujours dans le Rio Grande do Sul, plusieurs gisements des environs d’Aceguá dans le district de Minuano ont donné les fossiles les plus complets du paréiasaure Provelosaurus[7]. Le site de la ferme Fagundes (São Gabriel, district de Catuçaba) a fourni un crâne fragmentaire de Provelosaurus[8], des dents de dinocéphales Anteosauridae, Titanosuchidae et Tapinocephalidae indéterminés[9], et un amphibien indéterminé[10]. Dans le district de Tiarajú (également près de São Gabriel) fut trouvé l’anomodonte basal Tiarajudens[11] - [12]. De la région de Serra do Cadeado (près d’Ortigueira, État du Paraná), proviennent l’amphibien Australerpeton[10], une espèce indéterminée du dicynodonte Endothiodon[13] (possiblement E. bathystoma[14]), ainsi qu’un dinocéphale tapinocéphalidé indéterminé montrant des similitudes avec les genres Moschops et Moschognathus[10]. Il existe toutefois des incertitudes sur la contemporanéité de tous ces sites. Les fossiles de vertébrés de la Formation de Rio do Rasto se trouvent dans des affleurements dispersés, isolés et discontinus, à cause d’un couvert végétal dense, ce qui rend difficile l’établissement de corrélations locales. De plus, la localisation précise de plusieurs découvertes anciennes est incertaine et plusieurs taxa pourraient provenir de niveaux stratigraphiques différents au sein de la Formation de Rio do Rasto. C’est le cas des fossiles du Paraná, où les restes de tapinocephalidé et du dicynodonte Endothiodon pourraient provenir de deux niveaux distincts[10]
Âge de la Formation de Rio do Rasto
L’estimation de l’âge de la formation de Rio do Rasto a longtemps reposée uniquement sur la base de corrélations biostratigraphiques. Depuis 2018, des datations radiométriques ont permis de préciser l’âge d’une partie de la formation[15] - [16].
À partir des faunes de conchostracés et de bivalves, Holz et des collègues ont suggéré que la succession sédimentaire de cette formation s’étendait du Wordien (Guadalupien moyen) au Wuchiapingien (base du Lopingien)[17]. Parmi les tétrapodes, la présence de dinocéphales sur les terres des fermes de Fagundes et de Boqueirão (État du Rio Grande do Sul) ainsi que dans la région de Serra do Cadeado (État du Paraná) indique un âge Guadalupien pour ces localités, étant donné que ce groupe de synapside est limité à cet âge dans le reste du monde. Dans le site de la ferme de Fagundes, les dinocéphales coexistent avec le paréiasaure Provelosaurus. Ce dernier est présent dans la région d’Aceguà environ 2 m au-dessus d’une couche de bentonite de 30 cm d'épaisseur daté radiométriquement à 266 ± 5,4 Ma[16] - [7]. La grande marge d’erreur de cette datation radiométrique situe les sites à dinocéphales du Membre de Morro Pelado dans l’intervalle de temps Roadien inférieur – Capitanien moyen, ce qui confirme l’âge Guadalupien de cette partie du Membre de Morro Pelado[7]. Les autres tétrapodes du Membre de Morro Pelado présents dans le Rio Grande do Sul suggèrent un âge ne dépassant pas le Wordien. Les tapinocéphalidés ne sont connus ailleurs que dans les roches du Wordien et du Capitanien, l’anomodonte basal Tiarajudens est étroitement apparenté au genre sud-africain Anomocephalus de la Zone d’association à Tapinocephalus dont l’âge s’étend du Wordien terminal au Capitanien supérieur, et le temnospondyle Konzhukovia, présent sur le site de la ferme Boqueirão à São Gabriel, est également connu dans l’est de la Russie européenne dans des strates limitées à l’intervalle Wordien - Capitanien[7]. Le probable âge Wordien du Membre de Morro Pelado est également renforcé par un âge radiométrique du Membre de Serrinha (partie inférieure de la formation de Rio do Rasto) obtenu à partir d’une couche de cendres volcaniques altérées en tonsteins qui a donnée un âge de 270.61 + 1.76/− 3.27 Ma correspondant au Roadien (Guadalupien basal)[15].
Classification
En décrivant Pampaphoneus, Cisneros et al. ont présenté plusieurs cladogrammes confirmant la reconnaissance des clades Anteosaurinae et Syodontinae érigés par le paléontologue Américain Christian F. Kammerer un an plus tôt. Dans toutes leurs analyses, Pampaphoneus est identifié comme le Syodontinae le plus basal[1] :
Paléobiogéographie
Durant le Permien, la plupart des terres émergées étaient réunies en un seul supercontinent, la Pangée, qui avait grossièrement la forme d’un C. Ses parties nord (la Laurasia) et sud (le Gondwana) étaient reliés à l’ouest mais séparés à l’est par une très vaste baie océanique - la mer de Téthys[18]. Un long chapelet de microcontinents, regroupés sous le nom de Cimmérie, divisait la Téthys en deux : la Paleo-Tethys au nord, et la Neo-Tethys au sud[19]. Dans l’hémisphère nord, les antéosaures sont connus dans l’est de la Russie d’Europe, au Kazakhstan, et en Chine, des territoires correspondant à des paléolatitudes moyennes comprises entre le 30e et le 40e parallèle nord[20]. Dans l’hémisphère sud, les fossiles de ces animaux se rencontre en Afrique du Sud, en Zambie et au Zimbabwe, des régions qui étaient toutes situées à des paléolatitudes élevées localisées au niveau du 60e parallèle sud[20] - [21]. Les gisements Brésiliens se trouvaient eux un peu plus au nord entre le 35e et le 45e parallèle sud[5]. La parenté étroite du Pampaphoneus brésilien avec des antéosaures sud-africains et russes suggère une dispersion à travers la Pangée occidentale plutôt que par la Pangée orientale via le pont cathaysien. Ce dernier incluait une partie du nord de la Chine, la Corée, le sud de la Chine et l’Indochine et a joué un rôle important dans la dispersion des thérapsides au Permien supérieur et au Trias. Cependant ce pont n’était probablement pas encore constitué au Permien moyen et des barrières marines morcelaient les différents blocs Cathaysiens rendant difficiles la dispersion d’animaux depuis la Cathaysie vers l’Afrique australe via l’Europe de l’Est. Cisneros et des collègues suggèrent une voie de migration à priori plus facile via l’ouest de la Pangée en privilégiant une reconstruction continentale de type Pangée B où l’Amérique du Sud était juxtaposée aux Appalaches et était ainsi plus proche de la Russie que sur le modèle de la Pangée A. Dans ce dernier modèle, les montagnes Appalachiennes et Mauritanides aurait été des barrières naturelles difficiles à franchir. En revanche, dans une configuration Pangée B, le Brésil était non seulement plus proche de l’Europe de l'Est, mais la seule barrière montagneuse le long du chemin était les Hercynides européennes d’altitudes modérées. Les antéosaures auraient migrés de la Russie (d’où proviennent les plus anciens spécimens) jusqu’en Afrique australe en passant par l’Europe orientale et l’Afrique occidentale (en contournant la chaîne des Mauritanides) puis, par l’actuel Brésil[1]. La découverte de probables empreintes de pas d’antéosaures dans le sud de la France[N 1], qui était alors situé à une paléolatitude basse au niveau du 10e parallèle nord, soutient également cette hypothèse, le sud-ouest de l’Europe se trouvant dans ce couloir migratoire[22] - [23] - [24] - [25].
Notes et références
Notes
- Ces empreintes de pas, initialement nommées Planipes caudatus et P. brachydactylus, sont aujourd’hui regroupées sous le nom de Brontopus antecursor (cf. références Marchetti et al. 2019).
Références
- (en) J.C. Cisneros, F. Abdala, S. Atayman-Güven, B.S. Rubidge, A.M.C. Şengör et C.L. Schultz, « Carnivorous dinocephalian from the Middle Permian of Brazil and tetrapod dispersal in Pangaea », Proceedings of the National Academy of Sciences of the United States of America, vol. 109, no 5, , p. 1584-1588 (DOI 10.1073/pnas.1115975109, lire en ligne)
- (pt-BR) M. Anilson Costa Santos et al., « Um Esqueleto Parcial do Dinocefálio Anteossaurídea Pampaphoneus biccai (Permiano do Rio Grande do Sul) », Anais do XXVI Congresso Brasileiro de Paleontologia. Campinas : Galoá, vol. 2, , p. 270-271 (lire en ligne)
- (pt-BR) C. Perceval Paz, M. Anilson Costa Santos et F. Lima Pinheiro, « O Primeiro Registro de Pampaphoneus biccai (Anteosauridae) Com Ossos Pós-Cranianos », Revista Multidisciplinar De Educação E Meio Ambiente, vol. 1 (1), , p. 133 (lire en ligne)
- (en) A.D.S. Boos, C.F. Kammerer, C.L. Schultz, M.B. Soares et A.L.R. Ilha, « A New Dicynodont (Therapsida: Anomodontia) from the Permian of Southern Brazil and Its Implications for Bidentalian Origins », Plos ONE, vol. 11, no 5, , e0155000 (DOI 10.1371/journal.pone.0155000)
- (en) N.R. Cúneo, « Permian phytogeography in Gondwana », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 125, , p. 75-104 (DOI 10.1016/S0031-0182(96)00025-9)
- (en) C.P. Pacheco, E. Eltink, R.T. Müller et S.A. Dias-da-Silva, « A new Permian temnospondyl with Russian affinities from South America, the new family Konzhukoviidae, and the phylogenetic status of Archegosauroidea », Journal of Systematic Palaeontology, vol. 15, no 3, , p. 241-256 (DOI 10.1080/14772019.2016.1164763)
- (en) J.C. Cisneros, P. Dentzien-Dias et H. Francischini, « The Brazilian Pareiasaur revisited », Frontiers in Ecology and Evolution, vol. 9, , p. 758802 (DOI 10.3389/fevo.2021.758802)
- (en) J.C. Cisneros, F. Abdala et M.C. Malabarba, « Pareiasaurids from the Rio do Rasto Formation, southern Brazil: Biostratigraphic implications for Permian faunas of the Paraná Basin », Revista Brasileira de Paleontologia, vol. 8, no 1, , p. 13-24 (DOI 10.4072/rbp.2005.1.02)
- (en) M.C. Langer, « The first record of dinocephalians in South America: Late Permian (Rio do Rasto Formation) of the Paraná Basin, Brazil », Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, vol. 215, , p. 69-95 (DOI 10.1127/njgpa/215/2000/69)
- (en) A.D.S. Boos, C.F. Kammerer, C.L. Schultz et V.D. Paes Neto, « A tapinocephalid dinocephalian (Synapsida, Therapsida) from the Rio do Rasto Formation (Paraná Basin, Brazil): Taxonomic, ontogenetic and biostratigraphic considerations », Journal of American Earth Sciences, vol. 63, , p. 375-384 (DOI 10.1016/j.jsames.2015.09.003)
- (en) J.C. Cisneros, F. Abdala, B.S. Rubidge, P.C. Dentzien-Dias et A.O. Bueno, « Dental Occlusion in a 260-Million-Year-Old Therapsid with Saber Canines from the Permian of Brazil », Science, vol. 331, no 6024, , p. 1603-1605 (DOI 10.1126/science.1200305)
- (en) J.C. Cisneros, F. Abdala, T. Jashashvili, A.O. Bueno et P. Dentzien-Dias, « Tiarajudens eccentricus and Anomocephalus africanus, two bizarre anomodonts (Synapsida, Therapsida) with dental occlusion from the Permian of Gondwana », Royal Society Open Science, vol. 2, , p. 1-27 (DOI 10.1098/rsos.150090)
- (en) A.D.S. Boos, C.L. Schultz, C.S. Vega et J.J. Aumond, « On the presence of the Late Permian dicynodont Endothiodon in Brazil », Palaeontology, vol. 56, , p. 837-848 (DOI 10.1111/pala.12020)
- (en) C.F. Kammerer et M.A. Ordoñez, « Dicynodonts (Therapsida: Anomodontia) of South America », Journal of South American Earth Sciences, vol. 108, , p. 103171 (DOI 10.1016/j.jsames.2021.103171)
- (en) H. Francischini, P. Dentzien-Dias, M. Guerra-Sommer, R. Menegat, J.O.S. Santos, J. Manfroi et C.L. Schultz, « A Middle Permian (Roadian) lungfish aestivation burrow from the Rio do Rasto Formation (Paraná Basin, Brazil) and associated U-Pb dating », Palaios, vol. 33, , p. 69-84 (DOI 10.2110/palo.2017.050)
- (en) A.C. Rocha-Campos, M.A.S. Basei, A.P. Nutman, P.R. Santos, C.R. Passarelli, F.M. Canile, O.C.R. Rosa, M.T. Fernandes, H. Santa Ana et G. Veroslavsky, « U-Pb zircon dating of ash fall deposits from the Paleozoic Paraná Basin of Brazil and Uruguay: a reevaluation of the stratigraphic correlations », The Journal of Geology, vol. 127, , p. 167-182 (DOI 10.1086/701254)
- (en) M. Holz, A.B. França, P.A. Souza, R. Iannuzzi et R. Rohn, « A stratigraphic chart of the Late Carboniferous/Permian succession of the eastern border of the Paraná Basin, Brazil, South America », Journal of South American Earth Sciences, vol. 29, , p. 381-399 (DOI 10.1016/j.jsames.2009.04.004)
- (en) S. McLoughlin, « The breakup history of Gondwana and its impact on pre-Cenozoic floristic provincialism », Australian Journal of Botany, vol. 49, , p. 271-300 (DOI 10.1071/BT00023)
- (en) A.M.C. Şengör, « Tectonics of the Tethysides: orogenic collage development in a collisional setting », Annual Review of Earth and Planetary Sciences, vol. 15, , p. 214-244 (DOI 10.1146/annurev.ea.15.050187.001241)
- (en) T.S. Kemp, « The origin and early radiation of the therapsid mammal-like reptiles : a palaeobiological hypothesis », Journal of Evolutionary Biology, vol. 19, no 4, , p. 1231-1247 (DOI 10.1111/j.1420-9101.2005.01076.x)
- (en) M.K. Bamford, « Fossil woods from the Upper Carboniferous to Lower Jurassic Karoo Basin and their environmental interpretation », dans Linol, B. & de Wit, M.J., Origin and evolution of the Cape Mountains and Karoo Basin, Cham, Regional Geology Reviews, Springer, (ISBN 978-3-319-40858-3), p. 159–167
- G. Gand, G. Demathieu et F. Ballestra, « La palichnofaune de vertébrés tétrapodes du Permien supérieur de l’Estérel (Provence, France) », Palaeontographica A235, vol. 4-6, , p. 97-139 (DOI 10.1127/pala/235/1995/97)
- G. Gand, J. Garric, G. Demathieu et P. Ellenberger, « La palichnofaune de vertébrés tétrapodes du Permien supérieur du bassin de Lodève (Languedoc-France) », Palaeovertebrata, vol. 29, no 1, , p. 1-82 (lire en ligne)
- (en) L. Marchetti, H. Klein, M. Buchwitz, A. Ronchi, R.M.H. Smith, W.J. De Klerk, L. Sciscio et G.H. Groenewald, « Permian-Triassic vertebrate footprints from South Africa: Ichnotaxonomy, producers and biostratigraphy through two major faunal crises », Gondwana Research, vol. 72, , p. 139-168 (DOI 10.1016/j.gr.2019.03.009)
- (en) S.L. Olroyd et C.A. Sidor, « A review of the Guadalupian (middle Permian) global tetrapod fossil record », Earth-Science Reviews, vol. 171, , p. 583-597 (DOI 10.1016/j.earscirev.2017.07.001)
Références taxinomiques
(en) Référence Paleobiology Database : Pampaphoneus Cisneros et al., 2012