Mouche tsé-tsé
Les mouches tsé-tsé (aussi orthographié tsétsé[1]) ou glossines sont un genre de mouches hématophages africaines qui peuvent être vectrices de trypanosomiases humaines (THA ou maladie du sommeil) ou animales (nagana). Ces maladies peuvent être mortelles.
Glossina

Sous-genres de rang inférieur
Histoire
Le mot « tsé-tsé » vient de la langue tswana, une des langues bantoues parlée dans plusieurs pays d'Afrique australe, et signifie « mouche qui tue le bétail[2] ».
Les premières espèces décrites (Glossina longipalpis et G. palpalis) l'ont été en 1830, mais leur rôle dans la transmission des trypanosomes africains n'est établi qu'à partir de 1895[3].
L'entomologiste Ernest Edward Austen (1867-1936) fait partie des grands spécialistes de la mouche tsé-tsé de son époque.
En 2004 est créé l'International Glossina Genome Initiative (IGGI) destiné à la mobilisation des ressources pour l'étude du génome des glossines en Afrique subsaharienne[4].
Morphologie
Adulte

Une mouche tsé-tsé ou glossine est un insecte diurne, mouche piqueuse de 6 à 16 mm selon les espèces (généralement un peu plus grande que la mouche domestique), allongée et robuste, de couleur gris sombre à brun clair, sans aucun reflet métallique[5] - [3].
Les mâles et les femelles ont la même apparence, les femelles étant toujours un peu plus grandes que les mâles[5] - [3].
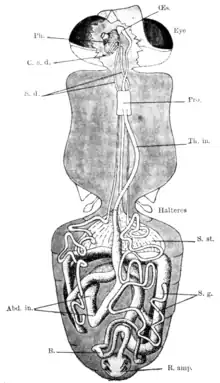
La tête porte deux yeux composés et trois ocelles, et deux petites antennes formées de trois articles dont le dernier plus allongé porte dorsalement des soies sensorielles (antenne dite aristée)[5].
Les pièces buccales forment une trompe (ou proboscis) constituée de trois pièces impaires : le labre, replié en gouttière, qui délimite le canal alimentaire, l'hypopharynx traversé sur sa longueur par un canal salivaire et le labium. Il existe aussi une pièce paire : les maxilles qui sont atrophiées mais garnies de palpes maxillaires aussi longs que la trompe qu'ils protègent dorsalement. Au repos, ces pièces longues et fines, renflées à leur base, restent horizontales et dirigées vers l'avant ; elles s'abaissent verticalement lors de la piqûre[5]. Lors du repas de sang, le labre et l'hypopharynx sont les seules pièces vulnérantes qui pénètrent le vertébré[3].

Le thorax porte une paire d'ailes et trois paires de pattes. Il s'agit d'ailes à nervures constantes et caractéristiques, notamment formant une cellule centrale en forme de hache ou couperet de boucher[5]. On reconnait immédiatement une glossine au repos à ses ailes croisées sur le dos comme les lames d'une paire de ciseaux[3].
À la base des ailes, à la limite de l'abdomen, se trouvent deux balanciers ou haltères. Ce sont des organes sensoriels qui permettent l'équilibre en vol.
Les six pattes sont formées chacune de cinq segments, le dernier est le tarse, constitué lui-même de cinq articles dont la couleur permet de différencier les sous-genres[5].
L'abdomen se compose de huit segments, dont sept sont visibles dorsalement. Le huitième porte les organes génitaux externes ou genitalia, saillantes et complexes chez les mâles, plus simples chez les femelles[5].
Larve
La femelle pond, sur un sol meuble, une larve au stade 3 (voir section reproduction), de la forme d'un asticot d'un cm de long environ, segmenté en 13 anneaux, de couleur jaune pâle et qui est souvent plus lourd que la mouche tsé-tsé adulte[6]. Cette larve porte à son extrémité deux protubérances noires représentant des lobes stigmatiques (rôle respiratoire), dits aussi lobes polypneustiques, qui sont caractéristiques du genre[5] - [7].
Cette larve est mobile mais ne se nourrit pas, elle s'enfonce dans le sol de quelques cm pour se transformer en une quinzaine de minutes en nymphe qui, comme celle des autres diptères, est appelée pupe, et qui change de couleur en quelques heures[7] - [8].
Pupe

Une pupe est semblable à la larve, mais de dimension inférieure (5 à 8 mm), noirâtre ou marron foncée, les deux lobes respiratoires restant visibles. Cette pupe est immobile, elle réalise ses métamorphoses dans une enveloppe protectrice étanche (durcissement du tégument de la larve) en forme de tonnelet appelé puparium[5].
Au bout de 4 à 6 semaines (durée variable selon l'espèce, le sexe et la température), l'insecte parfait ou imago sort de son puparium, émerge du sol, et y reste moins d'une heure avant de s'envoler[7] - [8]. En conséquence pour la lutte antivectorielle, on ne peut lutter directement que contre les adultes[7].
Biologie
Par rapport aux autres Muscoidea, les glossines se distinguent par leurs pièces buccales adaptées à la piqûre et leur mode de reproduction particulier, de type vivipare.
Alimentation
Les deux sexes sont hématophages obligatoires. En condition naturelle, le premier repas de sang est pris un à deux jours après l'émergence au stade adulte, puis tous les 4 jours en moyenne pour les mâles. En sus, les femelles doivent se nourrir trois fois en cours de gestation. Le rythme des repas varie selon l'activité de l'individu en fonction des conditions climatiques et des hôtes disponibles[7].
.TIF.jpg.webp)
La piqûre a lieu de jour, et dans des zones particulières fréquentées par les hôtes (bords de routes, pistes des troupeaux, points d'eau, gués...)[8]. Dans des conditions très favorables, l'intervalle entre les repas peut augmenter jusqu'à une dizaine de jours. En situation défavorable (stress nutritionnel), les femelles donnent des pupes de plus petite taille, avec baisse immunitaire, en étant plus à risque de s'infecter par trypanosomes[7].
La piqûre de glossine est plus ou moins douloureuse avec un petit saignement, et la mouche piqueuse peut être tuée par un mouvement défensif de l'hôte. Les hôtes préférés sont les animaux à sang chaud, en particulier les ongulés sauvages et domestiques où, pour plus de sûreté, la tsé-tsé a tendance à piquer sur l'abdomen ou la partie inférieure des membres antérieurs. Certaines espèces de glossines sont capables de s'alimenter aussi sur des reptiles qui ne se défendent guère contre les piqûres[7].
Le métabolisme des glossines ne se baserait pas principalement sur la consommation de glucides, mais sur celle de l'alanine, de la proline, des diglycérides et triglycérides, ce qui serait confirmé par l'analyse de leur génome[7].
Reproduction
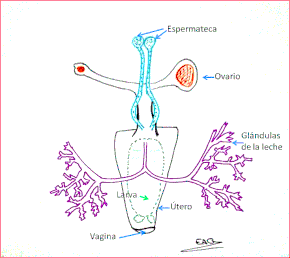
L'accouplement a lieu quelques heures, au maximum quelques jours, après émergence des adultes. Cet accouplement est long (deux à trois heures) au cours duquel le mâle dépose un spermatophore à la base de l'utérus. Bien que plusieurs accouplements soient possibles pour la femelle, on considère que seul le premier accouplement est fécondant, la femelle conservant le premier sperme dans deux spermathèques[7] - [8].
La mouche tsé-tsé femelle produit un œuf à la fois qu'elle conserve dans son utérus où il éclot et se développe en larve[6].
Le cycle ovarien de la glossine est alternatif entre ovaire droit et ovaire gauche : l'ovaire qui a donné un ovocyte se met au repos, tandis que l'autre développe un nouvel ovocyte. Une nouvelle ovulation est alors déclenchée après parturition ou avortement du germe précédent[9].
Les glossines se reproduisent par viviparité dite adénotrophique : après fécondation, l'œuf donne une larve L1 au bout de 3 à 4 jours. Cette larve continue à se développer jusqu'au stade L3 dans l'utérus de la femelle pleine. Ce temps de gestation dure deux semaines environ. Cette larve intra-utérine respire par ses deux stigmates[8], et elle est alimentée par des sécrétions de glandes lactifères annexées à l'utérus. Il s'agit de sécrétions de protéines et de lipides, proche du « lait » des marsupiaux[3].

Une femelle ne pond pas des œufs mais « accouche » d'un seul asticot (larve au stade L3), phénomène appelé larviposition. Cette larviposition dure quelques minutes, par dépôt au sol. Les sites de larviposition doivent combiner l'ombre et l'humidité, de préférence sur un sol meuble. Ces sites peuvent varier selon l'habitat (savane, forêt, rivière) des espèces de glossines[7] - [8].
Par exemple, pour les espèces de rivières, ils varient selon la saison, les crues et décrues : en saison sèche, la larviposition se fait sur les berges des cours d'eau, et à plus longue distance à la saison des pluies. Pour les espèces de savane et de forêt, elle peut se faire dans des terriers d'animaux ou à la base de certaines plantes[7].
Contrairement à d'autres insectes qui pondent des centaines d'œufs, la progéniture d'une tsé-tsé est en faible nombre (de 6 à 10 le plus souvent au cours de sa vie, l'intervalle entre deux larvipositions d'un seul asticot étant de 9-10 jours environ). Ce petit nombre est compensé par[3] - [9] :
- de meilleures chances de survie au cours du développement (puparium souterrain) ;
- durée de vie adulte relativement longue (2 à 4 mois, jusqu'à 5 en conditions optimales) avec des capacités à survivre en faible densité ;
- le fait que les femelles âgées restent fertiles sans baisse reproductive.
Activité
Les glossines sont peu mobiles, du fait d'un métabolisme énergétique peu efficace, basé non pas sur les glucides mais sur une combinaison proline-alanine (« mauvais carburant ») et qui nécessite des moments de récupération fréquents pour la tsé-tsé[10].
Une femelle se déplace en moyenne 5 minutes par jour, et jusqu'à 50 minutes pour les mâles. Les vols sont rapides mais de courte durée et à hauteur très faible de 0,5 m, ce qui a fait parler de « sauts ». Ces déplacements ont pour but la recherche d'un hôte (repas de sang) ou d'un site de repos pour les deux sexes, de femelles pour les mâles, et de sites de larviposition pour les femelles gravides[10].
Des femelles âgées sont capables de se déplacer ainsi sur plus de 10 km. Un déplacement passif est possible (mouches transportées par bovins transhumants ou moyen humain de transport)[10].
Les capacités visuelles et olfactives des glossines ont été très étudiées (mise au point de systèmes de piégeage). Les glossines du groupe Fusca et Palpalis sont attirées par l'odeur humaine, laquelle repousse au contraire les glossines du groupe Morsitans[10].
Génome
Le séquençage du génome de la mouche tsé-tsé est disponible depuis 2014[3], il s'agissait de Glossina morsitans[4]. Depuis plusieurs autres génomes de glossines représentant l'ensemble des sous-genres sont connus et font l'objet d'étude comparative[11].
Une glossine possède trois paires de chromosomes. Son génome est d'une taille totale de 366 mégabases, soit le double de Drosophila melanogaster mais avec un nombre plus faible de gènes (environ 12 000). Plusieurs caractéristiques de la tsé-tsé se retrouvent dans leurs gènes[3] :
- Hématophagie : 10 gènes codent des aquaporines qui facilitent le passage de l'eau à travers les membranes biologiques, c'est le nombre le plus élevé trouvé chez un insecte, en rapport avec son régime sanguin exclusif. D'autre part, les glandes salivaires sécrètent plus de 250 protéines différentes liées à cette hématophagie ;
- les gènes de l'olfaction sont en nombre réduit par rapport aux autres insectes ; les gènes visuels montrent une conservation de la détection du mouvement et une vision dans le champ des longueurs d'onde du bleu[11] ;
- viviparité : des gènes codent des protéines spécifiques pour la production de « lait », dont l'un des principaux a été appelé ladybird late[9] ;
- faible nombre de gènes du métabolisme glucidique, une réduction marquée par rapport à celle des moustiques ou des phlébotomes par exemple ;
- dans l'ADN de glossine on trouve des transferts d'ADN de bactéries symbiotiques, en particulier 1 mégabase d'ADN de Wolbachia.
Ces études permettent d'appréhender l'évolution moléculaire des glossines et d'orienter les recherches vers de nouvelles stratégies de contrôle de leurs populations[11].
Bactériome et symbiotes
Le régime hématophage exclusif des glossines nécessite une supplémentation nutritionnelle, notamment en vitamines. Cette supplémentation est apportée par des microorganismes associés, tels que des bactéries intracellulaires (bactériome) qui peuvent être aussi en endosymbiose[10] - [9].
Trois symbiotes sont transmis de la mère à la larve intra-utérine, soit verticalement (par les ovaires) soit par l'intermédiaire des sécrétions lactées[12] :
- Wigglesworthia glossinidia est situé dans les cellules épithéliales de l'intestin, et extracellulaire dans les glandes lactifères. Il synthétise des vitamines B nécessaires à fertilité et à l'immunité. Wigglesworthia doit être transmis par le lait à la larve intrautérine, faute de quoi l'adulte issu de cette larve présentera des déficits immunitaires, et si c'est une femelle des avortements[10] - [9] ;
- Wolbachia, notamment W. pipientis, est présent dans les tissus des gonades. Il intervient dans la reproduction, l'immunité et la longévité[10] - [12] ;
- Sodalis glossinidius (en) est présent de manière intra- et extracellulaire, il affaiblit la résistance immunitaire de la tsé-tsé, en facilitant les infections à trypanosomes[10] - [13].
D'autres bactéries et virus, symbiotiques ou non symbiotiques, ont été décrits chez la glossine, comme la bactérie Serratia glossinae dont on ne connait pas exactement le rôle, et le virus de l'hyperplasie des glandes salivaires (appartenant à la famille des Hystrosaviridae (en)) qui peut faire s'effondrer les populations de glossines en insectarium[10] - [12].
L'identification, et la connaissance du rôle exact, de ces symbiotes pourrait en faire des cibles potentielles pour le contrôle des populations de tsé-tsé[13].
Écologie et répartition géographique
Conditions climatiques
Les glossines ont besoin d'une température et d'une humidité tropicales. Leur climat habituel est celui de zone boisée avec plus de 600 mm de pluviométrie annuelle, un optimum thermique de 25 °C pour toutes les espèces, et une humidité de 50-60 % (glossines de savane) à 65-85 % (glossines de forêt et de rivière)[3].
Les glossines souffrent lors de température supérieure à 36 °C et meurent à plus de 38 °C. À température inférieure de 16 °C, les adultes sont paralysés par le froid et les pupes ne se développent pas. Elles sont donc absentes des zones sahéliennes et désertiques, comme des régions montagneuses et fraîches[3].
Répartition
Les glossines sont uniquement présentes en Afrique subsaharienne et dans quelques îles proches (celle de Zanzibar a été éradiquée dans les années 2000) ; quelques spécimens ont été trouvés dans le sud-ouest de l'Arabie saoudite. Depuis le milieu du XXe siècle, leur aire géographique est fortement réduite par anthropisation et épisodes de sècheresse, surtout en zones de savane où les mouches tsé-tsé ne se trouvent plus que le long de certaines rivières ou dans des zones naturelles classées (parcs et forêts)[3].

Différentes espèces de glossines peuvent être sympatriques dans le même habitat. Toutes les espèces sont capables de transmettre des trypanosomes, mais avec une compétence vectorielle différente, ce qui implique la nécessité d'identifier les espèces ayant un rôle-clé dans cette transmission[14].
La répartition géographique des glossines est fragmentée en de nombreux habitats avec différentes tendances spatio-temporelles. Leur densité apparente générale peut être diminuée, mais avec des foyers locaux beaucoup plus denses. Par exemple, des espèces vectrices dangereuses, comme G. palpalis et G. fuscipes, sont capables de s'adapter à l'environnement humain. Les zones péri-urbaines ainsi touchées sont celles de villes comme Kinshasa, Libreville, Bonon et Bangui[14].
En Afrique Australe, où se trouvent des régions montagneuses, le réchauffement climatique pourrait entrainer l'occupation de nouveaux territoires pour la tsé-tsé[3].
Espèces de glossines
Systématique
Les glossines sont des diptères (ordre) brachycères (sous-ordre) cyclorrhaphes (infra-ordre). Elles constituent une seule famille, celle des Glossinidae, proche des Muscidae. La famille de Glossinidae ne comprend qu'un seul genre, le genre Glossina[3].
En 2019, le genre Glossina comporte vingt-deux espèces ou trente-trois en ajoutant les sous-espèces[11]. Il se répartit en trois sous-genres (renommés en « groupe » par les anglophones) basés sur la combinaison de la distribution géographique, de l'habitat et des caractères morphologiques (forme des pièces génitales externes, coloration des pattes, présence de soies…)[15] - [16].
Plusieurs sous-espèces seraient des espèces différentes, mais il s'agit d'un problème réservé à un petit nombre de spécialistes (par exemple approches informatiques sur l'évolution des génomes ou sur la géométrie des nervures des ailes) . La classification reste inchangée dans ses grandes lignes[16], malgré des discussions sur certaines espèces comme G. austeni[11].
Mouches de savane
Elles constituent le sous-genre Glossina ss (sensu stricto) ou groupe Morsitans. Elles sont de taille moyenne (8 à 10 mm) avec un abdomen à taches sombres sur fond clair[17]. Elles se trouvent surtout dans les savanes boisées, les fourrés denses et les forêts claires.
._Coloured_drawing_by_A.J.E_Wellcome_V0022550.jpg.webp)
Elles se nourrissent principalement sur la faune sauvage et le bétail domestique (bovidés et suidés). Elles tendent à disparaitre avec l'anthropisation (extension des terres cultivées). Ce sont les meilleurs vecteurs de la trypanosomose animale, en particulier ceux indiqués par un astérisque ci-dessous[16]:
- Glossina austeni (Newstead, 1912) ;
- Glossina longipalpis (Wiedemann, 1830) ;
- Glossina morsitans centralis (Machado, 1970)* ;
- Glossina morsitans morsitans (Wiedemann, 1850)* ;
- Glossina morsitans submorsitans (Newstead, 1911) ;
- Glossina pallidipes (Austen, 1903)* ;
- Glossina swynnertoni (Austen, 1923)*.
G. austeni est aussi un vecteur important du point de vue économique et qui, selon les auteurs, devrait être déplacé dans le groupe Palpalis ou dans un quatrième sous-genre Machadomia[11].
Mouches de forêt
Sous-genre Austenina ou groupe Fusca. Elles sont de grande taille (11 à 14 mm) avec un abdomen brun plus ou moins clair[17]. Elles occupent le plus souvent les forêts denses et humides, à l'exception de celles de l'est africain comme G. brevipalpis (fourrés d'Afrique orientale et australe) et G. longipennis (savanes arides du Kenya)[16].
Selon les espèces, elles se nourrissent sur des suidés ou de grands bovidés, mais aussi dans l'est africain, sur les éléphants, rhinocéros et hippopotames[8]. Elles résistent mal à l'anthropisation, et leur rôle vecteur parait peu important à l'exception de G. brevipalpis[16].
- Glossina brevipalpis (Newstead, 1911)* ;
- Glossina fusca congolensis (Newstead and Evans, 1921) ;
- Glossina fusca fusca (Walker, 1849) ;
- Glossina fuscipleuris (Austen, 1911) ;
- Glossina frezili Gouteux, 1988[18] ;
- Glossina haningtoni (Newstead and Evans, 1922) ;
- Glossina longipennis (Corti, 1895) ;
- Glossina medicorum (Austen, 1911) ;
- Glossina nashi (Potts, 1955) ;
- Glossina nigrofusca hopkinsi (Van Emden, 1944) ;
- Glossina nigrofusca nigrofusca (Newstead, 1911) ;
- Glossina severini (Newstead, 1913) ;
- Glossina schwetzi (Newstead and Evans, 1921) ;
- Glossina tabaniformis (Westwood, 1850) ;
- Glossina vanhoofi (Henrard, 1952).
Mouches de rivière
Sous-genre Nemorhina ou groupe Palpalis. Elles sont de petite taille (6 à 10 mm) avec un abdomen sombre ou tacheté de noir, des pattes postérieures garnies de grosses soies noires ou brunes. Elles sont liées à la végétation (site de dépôt de larve) bordant les plans et cours d'eau (forêt galerie, berge des rivières et des lacs, mangrove, bolons, niayes…)[17].
._Coloured_drawing_by_A.J.E._Wellcome_V0022551.jpg.webp)
Elles s'adaptent à l'anthropisation, en occupant les zones péri- et suburbaines. Très opportunistes du point de vue alimentaire (homme, bétail, reptile en particulier le crocodile), elles transmettent les trypanosomes humains aussi bien qu'animaux[8] - [16].
- Glossina caliginea (Austen, 1911) ;
- Glossina fuscipes fuscipes (Newstead, 1911)* ;
- Glossina fuscipes martinii (Zumpt, 1935) ;
- Glossina fuscipes quanzensis (Pires, 1948)* ;
- Glossina pallicera pallicera (Bigot, 1891) ;
- Glossina pallicera newsteadi (Austen, 1929) ;
- Glossina palpalis palpalis (Robineau-Desvoidy, 1830)* ;
- Glossina palpalis gambiensis (Vanderplank, 1911)* ;
- Glossina tachinoides (Westwood, 1850)*.
L'astérisque indique les vecteurs principaux de trypanosomes[16].
Glossines et trypanosomes
Les glossines sont les insectes vecteurs exclusifs de trypanosomes, parasites sanguins à l'origine de trypanosomiases africaines, humaine (maladie du sommeil) dont la gravité est marquée par une méningoencéphalite, et animale (nagana) qui infecte le bétail, le rendant anémique, moins fertile et réduisant la production de viande et de lait.
Les glossines ne transmettent que des trypanosomes, à l'exclusion des autres agents pathogènes. Cependant, par leur nombre et la fréquence de leurs piqûres, elles peuvent aussi entrainer des anémies graves du bétail, et par les blessures infligées favoriser d'autre parasitoses comme les myiases[19].
Cycle interne chez la glossine
Les trypanosomes sont ingérés lors d'un repas sanguin sur un hôte infecté (mammifère). Ils évoluent dans le tube digestif de l'insecte selon un parcours complexe, plus ou moins long, pour se multiplier et se différencier.
Ils remontent ensuite à contre-courant pour se focaliser en dernier lieu dans les glandes salivaires. C'est là où s'effectue la dernière transformation du trypanosome, celle qui aboutit à la forme métacyclique, forme infestante qui est transmise par la salive à un nouvel hôte mammifère[20] - [21].
Trois types de développement sont décrits selon les espèces de trypanosomes :
- sous-genre Duttonella (principal représentant T. vivax), le cycle est court se limitant à la trompe (proboscis) et à la cavité buccale (cibarium) de la glossine ;
- sous-genre Nannomonas (comme T. congolense), le cycle va jusqu'à l'intestin antérieur ;
- sous-genre Trypanozoon (comme T. brucei gambiense et T. brucei rhodesiense), le cycle va jusqu'à l'intestin postérieur.
Ces trajets distincts seraient le résultat d'une coévolution entre glossines et trypanosomes, pour limiter les compétitions entre trypanosomes et favoriser leur transmission[22].
Outre leur rôle de maintien et de transmission des trypanosomes, les glossines sont un lieu d'échanges génétiques, sexués ou non, entre populations de trypanosomes. De plus, lors de la migration finale vers les glandes salivaires, il se produit un goulot d'étranglement évolutif où le nombre des individus s'effondre, ce qui favorise ensuite la multiplication d'éventuels variants rares. Dès lors, les trypanosomes seraient le plus souvent « génétiquement modifiés » lors de leur transmission par glossine[22].
Transmission
Une fois infestée, une glossine le reste toute sa vie, mais seule une petite minorité des mouches infestées (le plus souvent moins de 10 %, voire moins de 1 % pour T. brucei ) sont susceptibles de transmettre la maladie. Ceci à cause de la complexité du cycle interne et selon la durée, de quelques jours pour Trypanosoma vivax, à trois semaines pour T. brucei[22].
Sur l'ensemble des espèces et sous-espèces connues de glossines, seules une douzaine sont des vecteurs efficaces de transmission. Le taux d'infection et la compétence vectorielle des tsé-tsé varient selon la région, le climat, l'espèce de tsé-tsé et l'espèce de trypanosome[22].
L'aptitude d'une glossine à assurer un développement parasitaire complet dépend aussi de son statut individuel : âge, sexe, statut nutritionnel, flore endosymbiotique. Le système immunitaire de la glossine joue un rôle crucial. Par exemple, une glossine jeune, au système immunitaire immature, en état de stress nutritionnel, sera plus sensible à l'infection et plus susceptible de transmettre des parasites par sa salive[22].
La transmission à l'Homme se fait dans un même biotope par piqûres fréquentes, soit d'homme à homme (cas de Trypanosoma brucei gambiense), soit d'animal sauvage à l'homme (cas de Trypanosoma brucei rhodesiense). La transmission aux animaux domestiques (principalement bovins) se fait lors de transhumance, rassemblement lors des marchés, ou proximité d'animaux sauvages[23].
On ne sait pas pourquoi la trypanosomiase africaine animale (nagana) des animaux domestiques peut apparaitre sur l'ensemble des territoires occupés par les glossines, alors que l'humaine (maladie du sommeil) n'existe que par foyers bien délimités, en dehors desquels la maladie n'existe pas bien que tous les acteurs soient présents (mêmes biotopes, glossines et hôtes)[23].
Surveillance et contrôle
Historique
Le premier piège à glossines a été mis au point et utilisé dans les plantations de l'île de Sao Tomé-et-Principe en 1910, il s'agissait de blouses noires enduites de glu (Da Costa et al., 1916). Pendant la première moitié du XXe siècle, les méthodes utilisées par les colonisateurs consistèrent surtout à réaliser des opérations de débroussaillage et d'abattage des animaux sauvages. Néanmoins, la lutte contre la maladie du sommeil, notamment pendant la grande pandémie qui a ravagé l'Afrique centrale et de l'Ouest entre 1920 à 1950, est d'abord passée par le dépistage et le traitement aux moyens d'équipes mobiles. C'est à cette époque que les chercheurs s'intéresseront vraiment aux pièges à glossines (Harris, 1930 ; Chorley, 1933 ; Morris, 1949). C'est avec l'apparition du DDT dans les années 1940 que les insecticides vont être utilisés massivement jusque dans les années 1960, période des indépendances pour de nombreux États africains.
Les pulvérisations aériennes ont été largement utilisées en Afrique australe, notamment au Zimbabwe (Rhodésie à l'époque), mais les problèmes posés par le rapport coût / efficacité de la méthode ainsi que son impact sur la faune non ciblée ont amené des chercheurs à mettre au point de nouveaux procédés. De plus, le fait que les glossines enterrent leurs larves rendait l'éradication quasi impossible et nécessitait des pulvérisations répétées. C'est ainsi que les pièges à glossines vont faire un retour en force à la fin des années 1960 jusque vers les années 2000. Le piège de référence en matière de lutte contre les glossines est le piège biconique (Challier et Laveissière, 1973), conçu pour attirer les glossines des groupes palpalis et fusca. Plusieurs déclinaisons de ce piège seront mises au point dans les années 1980, améliorant l'efficacité et réduisant les coûts de fabrication : le piège pyramidal (Lancien, 1981 ; Gouteux et Lancien, 1986), le piège monoconique « vavoua » (Laveissière et Grébaut, 1990).

En 1978, la conférence internationale d'Alma-Alta reconnaît aux communautés le droit de s'impliquer dans leur propre système de santé. Plusieurs campagnes massives de lutte contre les tsé-tsé sont alors lancées, notamment en Afrique francophone. Ces campagnes démontrent l'efficacité de la méthode. Un tel succès est dû pour l'essentiel à l'implication d'équipes spécialisées, capables d'appliquer à la lettre les procédures de mise en place des dispositifs. Malheureusement, dans les pays qui s'essaient ensuite à des campagnes de grande envergure, le non-respect de ces procédures provoque des échecs qui relèguent au second plan la prophylaxie par piégeage du vecteur.
Concernant la protection du bétail, d'autres techniques que le piégeage sont préconisées, qui passent par l'utilisation d'insecticide directement sur l'animal (pour-on, imprégnation des membres inférieurs). En effet, si l'on fait abstraction de la pulvérisation aérienne, le défi majeur de la protection du bétail réside dans l'immensité des surfaces à protéger. La mise en œuvre d'une protection directement sur l'animal présente dès lors tout son intérêt.
La technique du lâcher d'insectes stériles ou SIT, mise au point dans les années 1960 et utilisée avec succès dans les grandes plantations industrielles du Sud des États-Unis et d'Amérique centrale, a été testée dans les années 1980 au Burkina Faso (Politzar et Cuisance, 1984). Cette technique consiste à produire une grande quantité de mâles et à les stériliser en les soumettant à des radiations. Ces insectes sont ensuite relâchés dans la nature et vont s'accoupler avec les femelles sauvages. Si le nombre de mâles stériles est au moins dix fois plus élevé que celui des mâles sauvages et si la population de glossines visée est géographiquement isolée, l'espèce est rapidement éradiquée.
Cette technique a été utilisée sur l'île de Zanzibar de 1994 à 1998 où, en quatre ans, plus aucune mouche tsé-tsé n'a été repérée[24]. Le programme d'éradication, mené en coopération entre la FAO et l'Agence internationale de l'énergie atomique (AIEA) a consisté dans un premier temps à réduire drastiquement la population de mouches tsé-tsé à l'aide d'insecticide[25], pour permettre à la technique de l'insecte stérile d'être efficace, ce n'est qu'après, que les mouches mâles stériles ont été relâchées. Un rapport de 50 pour 1 a été utilisé sur Zanzibar, de manière à s'assurer du succès de l'opération.
Depuis le succès de Zanzibar, l'Union africaine désire étendre l'utilisation de cette technique à d'autres pays africains.
Les opposants à ce projet soutiennent que la suppression totale de cette espèce risque de déséquilibrer l'écosystème, dans lequel la mouche tsé-tsé a sans doute sa place[26]. Un programme important d'éradication de la mouche tsé-tsé a déjà été appliqué dans les années 1970-1980 dans la région de Ngaoundéré. Une continuité de ce programme au début des années 1990 consistait à utiliser des pièges à insectes de façon à constituer un barrage naturel à l'expansion de l'insecte dans la zone considérée comme éradiquée. Cette initiative aura permis une amorce du développement de la production laitière entraînant avec elle un ralentissement des transhumances du bétail et un début de la culture fourragère sur le plateau de l'Adamaoua.
Un point faible du SIT est constitué par sa lourdeur et son coût. La production des mâles stériles sur un mode industriel est délicate, onéreuse et nécessite un équipement spécialisé pour la stérilisation (bombe au cobalt). De plus, la réussite de cette technique, qui se traduit par l'éradication d'une population de glossines, passe par l'identification de populations de tsé-tsé isolées en s'appuyant sur de longues et coûteuses études de génétique.
La région de Dafinso au Burkina Faso a fait l'objet d'une campagne d'éradication de ces vecteurs en 1993 et 1994, la maladie ayant affecté des ânes notamment en 1992[27].
PATTEC
À la conférence de l'Organisation de l'unité africaine (OUA) de 2000 à Lomé (Togo), les chefs d'État africains ont décidé de faire un effort significatif en matière de lutte contre les glossines et ont lancé la Pan-African Tsetse and Trypanosomosis Eradication Campaign (PATTEC). Cette campagne, initiée à l'échelle continentale, vise à soutenir les États désireux de se débarrasser du problème trypanosomien. La FAO, l'OMS et l'IAEA sont les principaux partenaires de ce projet et le SIT son fer de lance. Il s'agit pour les États concernés de définir et de mettre en œuvre une politique visant l'éradication des tsétsés, qui passe par la formation de personnels, la mise en œuvre de moyens spécifiques (fabrications de pièges, production de mâles stériles, inventaire des populations de glossines génétiquement isolées, etc.) et de coordinations au niveau des sous-régions. Malheureusement, dix ans après l'initiation de cette campagne, le bilan reste très mitigé même si 13 États africains ont mis en œuvre des programmes dans le cadre de la PATTEC.
Le piégeage

Le principe de fonctionnement des pièges à glossines est fondé sur son attractivité visuelle. Si le bleu est la couleur la plus attractive pour les glossines, le bleu retenu pour la réalisation des pièges n'est pas le plus attirant. En effet, une des contraintes étant constituée par l'utilisation du tissu comme support à l'insecticide, une des qualités recherchée est la rémanence maximale du produit sur le support. Le bleu « roi » ou bleu « électrique » est le bleu de référence pour les pièges à glossines. Les pyréthrénoïdes (Deltamethrine, alphacypermethrine) sont les insecticides les plus performants, à condition que les tissus utilisés pour les pièges soient en coton/polyester, polyester ou polyamide. Le coton seul favorise le lessivage rapide du produit et est donc déconseillé. Si les moyens le permettent, l'attractivité peut être renforcée par l'adjonction d'un attractif olfactif ; les plus performants étant le CO2, l'acétone et certains phénols.
Le camouflage
Dans les années 1970, des recherches ont pointé le fait que la mouche tsé-tsé est attirée par la vue de larges zones monochromes : les rayures du zèbre permettraient ainsi de se protéger du parasite auquel les équidés sont plus sensibles que d'autres animaux sauvages. Il est à ce sujet significatif de constater que les zones de répartition des zèbres et des glossines coïncident exactement et que les rayures s'estompent chez les populations moins exposées au parasitisme par les trypanosomes[28] - [29].
Impact sur les sociétés africaines
Dans les régions où la mouche tsé-tsé est très présente, l'élevage de gros herbivores, notamment les bovins et les chevaux, devient improductif, ou même impossible, en raison de la mortalité des bêtes par trypanosomiases. Selon certains auteurs, cette donnée a considérablement ralenti le développement de l'Afrique tropicale[30].
En effet, la rareté ou l'absence de ces animaux a affecté les sociétés des régions touchées de plusieurs façons :
- agriculture moins productive, en l'absence d'animaux de trait et d'engrais naturels ;
- impossibilité de développer des réseaux de poste aux chevaux ;
- armées moins mobiles en l'absence de cavalerie, compliquant la constitution de grands empires.
Au cours du XIXe siècle, la déforestation a conduit à une extension spectaculaire des régions touchées par la mouche tsé-tsé. Ainsi, les premiers colons installés dans le futur Liberia avaient amené des chevaux et en faisaient le même usage qu'aux États-Unis. L'arrivée de la mouche tsé-tsé a causé l'effondrement des élevages équins de la colonie, ravageant son économie[31].
Le royaume d'Oyo, pays précolonial dans l'actuel Nigeria, entretenait une force de cavalerie. Si cette arme lui conférait un atout considérable sur ses voisins, elle avait un coût très important : la mortalité des chevaux était trop élevée pour permettre au royaume d'être autosuffisant, des animaux étaient donc en permanence importés du Sahel, région qui n'est pas infectée[32].
Références
- le Rapport de 1990 sur les rectifications orthographiques recommande cette écriture (Liste C).
- (fr) « Programme Against African Trypanosomiasis (PAAT) - Transmission », Organisation des Nations unies pour l'alimentation et l'agriculture (consulté le ).
- Duvallet 2017, p. 367-369.
- « Genome Sequence of the Tsetse Fly (Glossina morsitans): Vector of African Trypanosomiasis », Science (New York, N.Y.), vol. 344, no 6182, , p. 380–386 (ISSN 0036-8075, PMID 24763584, PMCID 4077534, DOI 10.1126/science.1249656, lire en ligne, consulté le )
- Rodhain 1985, p. 224-229.
- Lee R. Haines, Glyn A. Vale, Antoine M. G. Barreaux et Norman C. Ellstrand, « Big Baby, Little Mother: Tsetse Flies Are Exceptions to the Juvenile Small Size Principle », BioEssays, vol. 42, no 11, , p. 2000049 (ISSN 0265-9247 et 1521-1878, DOI 10.1002/bies.202000049, lire en ligne, consulté le )
- Duvallet 2017, p. 371-373.
- Rodhain 1985, p. 231-235.
- Joshua B. Benoit, Geoffrey M. Attardo, Aaron A. Baumann et Veronika Michalkova, « Adenotrophic Viviparity in Tsetse Flies: Potential for Population Control and as an Insect Model for Lactation », Annual review of entomology, vol. 60, , p. 351–371 (ISSN 0066-4170, PMID 25341093, PMCID 4453834, DOI 10.1146/annurev-ento-010814-020834, lire en ligne, consulté le )
- Duvallet 2017, p. 373-374.
- Geoffrey M. Attardo, Adly M. M. Abd-Alla, Alvaro Acosta-Serrano et James E. Allen, « Comparative genomic analysis of six Glossina genomes, vectors of African trypanosomes », Genome Biology, vol. 20, , p. 187 (ISSN 1474-7596, PMID 31477173, PMCID 6721284, DOI 10.1186/s13059-019-1768-2, lire en ligne, consulté le )
- Henry M. Kariithi, Drion G. Boucias, Edwin K. Murungi et Irene K. Meki, « Coevolution of hytrosaviruses and host immune responses », BMC Microbiology, vol. 18, no Suppl 1, , p. 183 (ISSN 1471-2180, PMID 30470186, PMCID 6251100, DOI 10.1186/s12866-018-1296-3, lire en ligne, consulté le )
- Anne Geiger, Imna Malele, Adly M Abd-Alla et Flobert Njiokou, « Blood feeding tsetse flies as hosts and vectors of mammals-pre-adapted African Trypanosoma: current and expected research directions », BMC Microbiology, vol. 18, no Suppl 1, , p. 162 (ISSN 1471-2180, PMID 30470183, PMCID 6251083, DOI 10.1186/s12866-018-1281-x, lire en ligne, consulté le )
- Florence Njeri Wamwiri et Robert Emojong Changasi, « Tsetse Flies (Glossina) as Vectors of Human African Trypanosomiasis: A Review », BioMed Research International, vol. 2016, , p. 6201350 (ISSN 2314-6133, PMID 27034944, PMCID 4789378, DOI 10.1155/2016/6201350, lire en ligne, consulté le )
- (en) A. M. Jordan, R. P. Lane (éditeur) et R. W. Crosskey (éditeur), Medical Insects and Arachnids, Chapman and Hall, , « Tsetse-flies (Glossinidae) ».
- Duvallet 2017, p. 369-371.
- Rodhain 1985, p. 229-231.
- (fr) J. P. Gouteux, « Une nouvelle glossine du Congo: Glossina (Austenina) frezili sp. nov. (Diptera: Glossinidae) », Tropical Medicine and Parasitology, Deutsche Tropenmedizinische Gesellschaft, vol. 38, no 2, , p. 97–100 (PMID 3629143, S2CID 91006636)
- Rodhain 1985, p. 241.
- Rodhain 1985, p. 236-238.
- Duvallet 2017, p. 377.
- Duvallet 2017, p. 380-381.
- Duvallet 2017, p. 382.
- (fr) La mouche tsé-tsé éradiquée dans l'île de Zanzibar, sur le site de la FAO.
- ce programme de réduction de la population par insecticide a été mené par la FAO et le FAO et le Programme des Nations unies pour le développement (PNUD).
- (fr) Éliminer la mouche tsé-tsé en Afrique, du magazine Afrique Relance (actuellement Afrique Renouveau) de mai 2003.
- Burkhard Bauer, Idrissa Kabore et Sandrine Amsler-Delafosse, « Lutte contre les vecteurs de la trypanosomose animale africaine au Burkina Faso », Cahiers Agricultures, vol. 4, no 6, , p. 440–443 (1) (ISSN 1777-5949, lire en ligne, consulté le )
- Bernard Vray, Le parasitisme : le plus vieux métier du monde ? ou les progrès de la paléo-parasitologie, pages 6-7.
- (en) A. Egri, M. Blaho, G. Kriska, R. Farkas, M. Gyurkovszky, S. Akesson et G. Horvath, « Polarotactic tabanids find striped patterns with brightness and/or polarization modulation least attractive: An advantage of zebra stripes », Journal of Experimental Biology, vol. 215, no 5, , p. 736–745 (PMID 22323196, DOI 10.1242/jeb.065540)
- (en) Marcella Alsan, « The Effect of the TseTse Fly on African Development », American Economic Review, vol. 105, no 1, , p. 382–410 (ISSN 0002-8282, DOI 10.1257/aer.20130604, lire en ligne, consulté le )
- D. C. Dorward et A. I. Payne, « Deforestation, the Decline of the Horse, and the Spread of the Tsetse Fly and Trypanosomiasis (nagana) in Nineteenth Century Sierra Leone », The Journal of African History, vol. 16, no 2, , p. 239–256 (ISSN 0021-8537, lire en ligne, consulté le )
- Robin Law, « A West African Cavalry State: The Kingdom of Oyo », The Journal of African History, vol. 16, no 1, , p. 1–15 (ISSN 0021-8537, lire en ligne, consulté le )
Annexes
Bibliographie
- F. Rodhain et C. Perez, Précis d'entomologie médicale et vétérinaire, Paris, Maloine, , 458 p. (ISBN 2-224-01041-9), chap. 9 (« Les glossines : systématique, biologie, importance médicale »), p. 225-248.

- Gérard Duvallet (dir.), Philippe Solano et Issa Sidibe, Entomologie médicale et vétérinaire, Marseille, Versailles, IRD - Quae, (ISBN 978-2-7099-2376-7), chap. 15 (« Les glossines (Diptera : Glossinidae) »), p. 367-389.
- Gary Sheffield, La première Guerre mondiale en 100 objets : Ces objets qui ont écrit l'histoire de la grande guerre, Paris, Elcy éditions, , 256 p. (ISBN 978 2 753 20832 2), p. 126-127.
Liens externes
- Ressources relatives au vivant :
- Global Biodiversity Information Facility
- (en) Animal Diversity Web
- (cs + en) BioLib
- (en) EPPO Global Database
- (en) Paleobiology Database
- (mul + en) iNaturalist
- (en) Invasive Species Compendium
- Ressource relative à la santé :
- Notices dans des dictionnaires ou encyclopédies généralistes :
- « Biologie, systématique et répartition des tsé-tsé », sur www.fao.org (consulté le )
- (en) Référence Animal Diversity Web : Glossina (consulté le )
- (en) Référence BioLib : Glossina Wiedemann, 1830 (consulté le )
- (en) Référence Catalogue of Life : Glossina (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023