Moschorhinus
Moschorhinus kitchingi
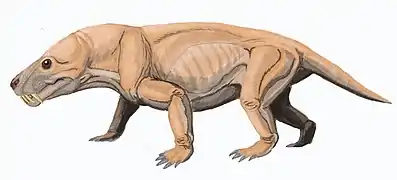
Moschorhinus est un genre éteint et fossile de thérapsides thérocéphales appartenant à la famille également éteinte des akidnognathidés. Une seule espèce est connue, Moschorhinus kitchingi, décrit par le paléontologue Robert Broom dès 1920 et désormais bien connu avec vingt-sept collections de fossiles enregistrées selon Paleobiology Database en 2022.
L'animal possède un museau large et émoussé qui porte de longues canines droites. Moschorhinus semble avoir chassé un peu comme un grand félin actuel et avoir remplacé écologiquement les gorgonopsiens carnivores. La comparaison avec la panthère nébuleuse actuelle a été évoquée scientifiquement.
Bien qu'il soit le plus abondant à la fin du Permien, il survit au début du Trias, donc après l'extinction du Permien, mais en rapetissant en taille. Il vit environ 11 millions d'années, entre 254 et 247 millions d'années avant notre ère, dans ce qui est aujourd'hui l'Afrique du Sud.
Découverte et étymologie
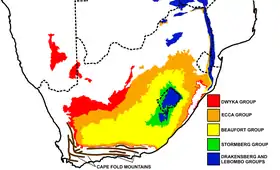
Le spécimen holotype est un crâne découvert par James Kitching dans le supergroupe du Karoo en Afrique du Sud, près du village de Nieu-Bethesda et décrit par le paléontologue Robert Broom en 1920[1].
Le nom de genre Moschorhinus provient du grec ancien μόσχος / moskhos « veau » et ῥινός / rhinós « nez » pour donner littéralement « nez de veau », en référence à la longueur du museau de l'animal. L'épithète spécifique kitchingi fait quant à elle référence à James Kitching, le paléontologue ayant découvert le spécimen holotype[1].
De nos jours, c'est l'un des thérapsides les plus connus et les plus reconnaissables de cette zone[2]. Les restes de Moschorhinus sont trouvables principalement dans le groupe de Beaufort sur une période allant du Permien supérieur au Trias inférieur[2] - [3] - [4].
Description
Description simplifiée
D'après les fossiles découverts en Afrique du Sud, Moschorhinus mesure environ deux mètres de long, faisant de lui l'un des plus grands thérocéphales connus. Le crâne est similaire à celui des gorgonopsiens, avec de grandes fosses temporales et un palais convexe. La taille du crâne est comparable à celle d'un varan et montrent la présence de très grands incisives de forme similaires à celles des célèbres tigres à dents de sabre datant de la fin du Cénozoïque[5] - [6].
Museau
Le museau de Moschorhinus est typiquement court et large. L'extrémité émoussée du museau présente une crête descendant le long de la ligne médiane jusqu'à l'os frontal[5] - [3]. Le museau supérieur dépasse un peu les incisives chez les juvéniles[3].
Les narines sont larges et positionnées vers le bout du museau[3].
Dentition
Moschorhinus a une formule dentaire de I6.C1.M3, c'est-à-dire six incisives, une canine et trois dents postcanines de chaque côté de la mâchoire supérieure[1].
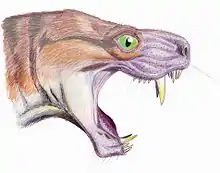
Les incisives sont logées dans les prémaxillaires. Ils sont grands, légèrement incurvés et ont une section transversale en forme de cloche. Ils ont des surfaces de coupe lisses et, contrairement à ceux des autres thérocéphales, manquent de facettes ou de stries résultant de l'abrasion et de l'usure[5].

Les grandes canines en forme de sabre sont maintenues dans les maxillaires et figurent parmi les caractéristiques rapidement identifiables de Moschorhinus. Ces grandes canines sont particulièrement épaisses et résistantes, et de section transversale uniquement circulaire. En longueur, ces sabres sont comparables aux gorgonopsiens. Bien qu'il n'y ait pas de véritable analogue moderne, l'exemple vivant le plus similaire serait celle de la Panthère nébuleuse[5].
Comme les autres thérocéphales, Moschorhinus a un nombre réduit de postcanines logées dans le maxillaire. Chez la plupart des thérocéphales, les « dents », ou plutôt la projection en forme de dent (denticulations) des os ptérygoïdiens, sont considérablement réduites ou manquantes, et chez Moschorhinus elles sont absentes[3] - [7].
Toit crânien
Traçant le toit crânien, Moschorhinus possède de petits os préfrontaux situé au-dessus des yeux, suivis de grands os frontaux élargis. Les os pariétaux forment une crête sagittale étroite le long de la ligne médiane du crâne, qui abrite un foramen pinéal très basique[1] - [3]. Des indentations peuvent être observées dans les fosses temporales, des dépressions de chaque côté de la crête, indiquant la présence de nombreux vaisseaux sanguins et nerfs alimentant le cerveau[8].
Orbites
L'os lacrymal est plus gros que le préfrontal réduit et forme la majorité de l'orbite. Le lacrymal a un bossage osseux sur l'orbite et un grand foramen vers sa face interne. Le bord inférieur de l'orbite est formé par l'os jugal et le maxillaire[1]. Le jugal se termine à l'orbite de l'œil et n'est pas convexe, comme chez plusieurs thérocéphales ultérieurs[3].
Palais
Dans l'ensemble, le palais est convexe, avec un large vomer triangulaire, avec des tubercules appariés, des projections arrondies pointant vers le ventral[5] - [3] semblable à d'autres akidnognathidés[1]. Les os palatins, formant l'arrière du toit de la bouche, sont élargis et épais, en particulier sur leurs bords externes où ils sont joints au maxillaire. Sur leurs bords internes, les palatins sont reliés au ptérygoïde et au vomer sur le nez, faisant partie de la circonférence de la cavité nasale. Entre le palatin et le maxillaire, juste derrière les canines, se trouvent de grands foramens, vraisemblablement pour permettre le passage des nerfs. Une crête inclinée le long du milieu du palais soutient vraisemblablement un palais mou, ce qui permet à l'air de circuler entre le nez et les poumons[5].
Les canines en formes de sabres nécessitent que la bouche s'ouvre largement pour être utilisées, ce qui rend l'alimentation difficile. Le genre apparenté Promoschorynchus présente des plis raides (crête choanale) à la frontière du passage nasal et de la gorge, utilisés pour le maintenir ouvert et pour permettre la respiration en mangeant. Le développement d'un palais secondaire dans le crâne a progressivement évolué chez les thérocéphales, et la crête choanale est présente chez tous les thérocéphales ultérieurs[7].
Classification
Moschorhinus est un thérocéphale classé dans la famille des akidnognathidés, des thérapsides carnivores possédants des crânes solides et de grandes canines sur la mâchoire supérieure[5].
Ci-dessous, les résultats simplifiés de l'analyse phylogénétique des Therocephalia basée d'après Liu et Abdala (2022)[9] :
| ◄ Therocephalia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paléobiologie
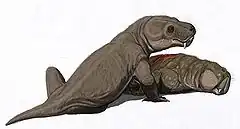
Les paléontologues supposent que Moschorhinus aurait été un superprédateur ayant un mode de vie semblable à celui des félins, capable de percer la peau et de s'accrocher à des proies en difficulté avec ses longues incisives. Il s'agit du premier fossile connu montrant ce type de technique de chasse. Compte tenu de son museau épais et robuste, de ses canines énormes et de ses puissants muscles de la mâchoire, Moschorhinus semble avoir été un prédateur remarquable[5].
Paléoécologie
De nombreux fossiles de vertébrés ont été découverts dans le bassin du Karoo. D'autres thérocéphales du même niveau rocheux sont Tetracynodon et Promoschorhynchus[2]. Les spécimens de Moschorhinus sont les seuls grands thérocéphales trouvé dans cette région[10] - [3].
Moschorhinus reprend la niche autrefois contrôlée par les gorgonopsiens. Les deux groupes sont construits un peu comme des grands félins. À la suite de l'extinction de Moschorhinus durant le Trias, les cynodontes prennent le contrôle d'une niche similaire[5].
Le genre semble s'être éteint au Trias inférieur peu de temps après l'extinction du Permien[11] - [12], les preuves fossiles montrent que les représentants du Trias ont grandi plus rapidement que ceux datant du Permien, ce qui a entraîné une réduction de la taille du corps chez ce dernier, ce qui est largement considéré comme un effet de la variabilité environnementale plus sévère après l'extinction du Permien (effet Lilliput (en))[2] - [3] - [13]. Les crânes fossiles datant du Permien mesurent en moyenne 207 mm de longueur, tandis que celles datant du Trias ne mesure que 179 mm[2]. Néanmoins, malgré la légère différence de taille, Moschorhinus, figure parmi les plus grands thérocéphales de son temps[2] - [10].
Voir aussi
- Therapsida
- Theriodontia
- Therocephalia
- Akidnognathidae
- Euchambersia, un thérocéphale plus petit appartenant à la même famille, mais suspecté d'être venimeux.
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
Références
- (en) Broom R, « On Some New Therocephalian Reptiles from the Karroo Beds of South Africa », Proceedings of the Zoological Society of London, , p. 351–354 (lire en ligne)
- (en) Huttenlocker AK, Botha-Brink J, « Body size and growth patterns in the therocephalian Moschorhinus kitchingi (Eutheriodontia) before and after the end-Permian extinction in South Africa », Paleobiology, vol. 39, no 2, , p. 253–77 (DOI 10.1666/12020)
- (en) Adam Huttenlocker, « The Paleobiology of South African Therocephalian Therapsids (Amniota, Synapsida) and the Effects of the End-Permian Extinction on Size, Growth, and Bone Microstructure », Université de Washington, (lire en ligne)
- (en) B. S. Rubidge et C. A. Sidor, « Evolutionary Patterns Among Permo-Triassic Therapsids », Annual Review of Ecology and Systematics, vol. 32, , p. 449–480 (DOI 10.1146/annurev.ecolsys.32.081501.114113)
- (en) van Valkenburgh B, Jenkins I, « EVOLUTIONARY PATTERNS IN THE HISTORY OF PERMO-TRIASSIC AND CENOZOIC SYNAPSID PREDATORS », Paleontological Society Papers, vol. 8, , p. 267–88 (lire en ligne [archive du ] [PDF])
- (en) Huttenlocker Adam, « An investigation into the cladistic relationships and monophyly of therocephalian therapsids (Amniota: Synapsida) », Zoological Journal of the Linnean Society, vol. 157, no 4, , p. 865–891 (DOI 10.1111/j.1096-3642.2009.00538.x
 , S2CID 84603632)
, S2CID 84603632) - (en) Maier W, van den Heever J, Durand F, « New therapsid specimens and the origin of the secondary hard and soft palate of mammals », Journal of Zoological Systematics and Evolutionary Research, vol. 34, , p. 9–19 (DOI 10.1111/j.1439-0469.1996.tb00805.x)
- (en) Durand J F, « A revised descripction of the skull of moschorhinus (therapsida, therocephalia) », Annals of the South African Museum, vol. 99, , p. 381–413 (lire en ligne)
- (en) J. Liu et F. Abdala, « The emblematic South African therocephalian Euchambersia in China: a new link in the dispersal of late Permian vertebrates across Pangea », Biology Letters, vol. 18, no 7, , p. 20220222 (ISSN 1744-957X, PMID 35857894, PMCID 9278400, DOI 10.1098/rsbl.2022.0222 )
- (en) Christian A. Sidor, Roger M. H. Smith, Adam K. Huttenlocker et Brandon R. Peecook, « New Middle Triassic Tetrapods from the Upper Fremouw Formation of Antarctica and Their Depositional Setting », Journal of Vertebrate Paleontology, vol. 34, no 4, , p. 793–801 (DOI 10.1080/02724634.2014.837472)
- (en) Peter D Ward, Jennifer Botha, Roger Buik, Michiel O. De Kock, Douglas H. Erwin, Geoffrey H Garrison, Joseph L Kirschvink et Roger Smith, « Abrupt and Gradual Extinction Among Late Permian Land Vertebrates in the Karoo Basin, South Africa », Science, vol. 307, no 5710, , p. 709–714 (PMID 15661973, DOI 10.1126/science.1107068 , CiteSeerx 10.1.1.503.2065)
- (en) Botha J, Smith RM, « Rapid vertebrate recuperation in the Karoo Basin of South Africa following the End-Permian extinction », Journal of African Earth Sciences, vol. 45, nos 4–5, , p. 502–14 (DOI 10.1016/j.jafrearsci.2006.04.006, lire en ligne)
- (en) Richard J Twitchett, « The Lilliput effect in the aftermath of the end-Permian extinction event », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 252, nos 1–2, , p. 132–144 (DOI 10.1016/j.palaeo.2006.11.038, lire en ligne)