Espèce réactive de l'azote
Les espèces réactives de l'azote (ERA), dérivés réactifs de l'azote ou espèces azotées réactives, aussi désignées sous le sigle « RNS » (de l'anglais reactive nitrogen species) sont une famille d'espèces chimiques jouant un rôle très important dans les systèmes biologiques et l'environnement (polluant atmosphérique) appartenant au groupe plus vaste des espèces réactives de l'oxygène et de l'azote (RONS, reactive oxygen and nitrogen species).
Source endogène/exogène
Tout comme pour les espèces réactives de l'oxygène, on distingue :
- les ERA d'origine exogène — produits par les UV solaires ou des rayonnements ionisants par exemple, qui en quantité anormale sont des polluants, car dans l'air et/ou dans les cellules, le taux de ces altéragènes chimiques croît significativement sous l'effet de certains stress ou polluants, par exemple chez les plantes sous l'effet de l'exposition aux ultraviolets, notamment dans des contextes tels que ceux du trou de la couche d'ozone et/ou du dérèglement climatique (comme pour les ROS)[2] — et ils peuvent alors endommager les structures cellulaires bactériennes, végétales, fongiques et animales, phénomène dénommé stress oxydant.
- les ERA endogènes, apparaissant comme sous-produits du métabolisme normal de l'azote et jouant alors un rôle majeur dans l'immunité et la lutte contre les microbes, et dans la communication cellulaire et intercellulaire. Ils apparaissent aussi sous l'effet de certains stress (chaleur excessive par exemple, situation qui sera également source de stress oxydatif par les espèces réactives de l'oxygène.
Synergies
Les espèces réactives de l'azote et de l'oxygène agissent synergiquement[2], notamment via la réaction suivante, en tant que polluants et dans les cellules et l'organisme vivant, selon la réaction :
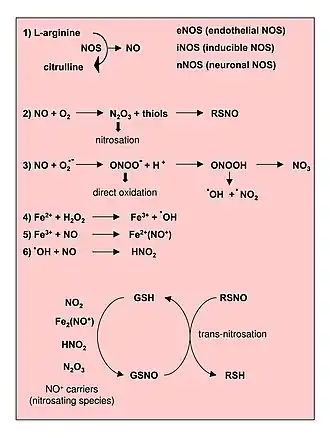
- •NO (oxyde nitrique) + O2•− (superoxyde) → ONOO− (peroxynitrite).
L'anion superoxyde (O2−) est une espèce oxygénée très réactive qui réagit rapidement avec l'oxyde nitrique (NO) dans le système vasculaire. La réaction produit du peroxynitrite qui épuise la bioactivité du NO. Ceci est important car le NO est un médiateur chimique-clé dans de nombreuses fonctions vasculaires importantes, notamment la régulation du tonus musculaire des muscles lisses, la régulation de la pression artérielle, l'activation des plaquettes et la signalisation des cellules vasculaires[3].
Les types d'ERA
Chez l'animal, les ERA sont produites à partir de la réaction de l'oxyde nitrique (•NO) avec le superoxyde (O2•−) pour former du peroxynitrite (ONOO−)[4] - [5]:(• NO) avec le superoxyde (O2 • -)
Le peroxynitrite lui-même est une espèce hautement réactive.
Il peut réagir directement avec diverses cibles biologiques et composants essentiels de la cellule, dont les lipides, les thiols, les résidus d'acides aminés, les bases d'ADN et les antioxydants de faible poids moléculaire[6]. Ces réactions se déroulent toutefois à un rythme relativement lente qui permet aux espèces réactives de l'azote de réagir plus sélectivement dans la cellule.
Remarque : le peroxynitrite peut, dans une certaine mesure, traverser les membranes cellulaires par les canaux anioniques[7].
De plus, le peroxynitrite peut réagir avec d'autres molécules pour former des types supplémentaires d'ERA, notamment le dioxyde d'azote (•NO2) et le trioxyde de diazote (N2O3), ainsi que d'autres types de radicaux libres chimiquement réactifs.
Les réactions importantes impliquant les espèces réactives de l'azote comprennent :
- ONOO− + H+ → ONOOH (acide peroxynitreux) → •NO2 (dioxide d'azote) + •OH (radical hydroxyle) ;
- ONOO− + CO2 (dioxyde de carbone) → ONOOCO2− (nitrosoperoxycarbonate) ;
- ONOOCO2− → •NO2 (dioxide d'azote) + O=C(O•)O− (radical carbonate) ;
- •NO + •NO2 ⇌ N2O3 (trioxyde de diazote).
Cibles biologiques
Le peroxynitrite peut réagir directement avec des protéines contenant des centres de métaux de transition. Il peut ainsi modifier des protéines telles que l'hémoglobine, la myoglobine et le cytochrome c en oxydant l'hème ferreux en ses formes ferriques correspondantes.
Le peroxynitrite peut aussi modifier la structure de protéines en réagissant avec divers acides aminés de la chaîne peptidique, notamment (réaction la plus courante) en oxydant la cystéine. Une autre réaction est la nitration de la tyrosine mais le peroxynitrite ne réagit pas directement avec la tyrosine. La tyrosine réagit avec d'autres ERA produits par le peroxynitrite.
Ces réactions affectent toutes la structure et la fonction de protéines ayant des fonctions majeures. Elles ont donc le potentiel de provoquer des changements dans l'activité catalytique des enzymes, des modifications de l'organisation cytosquelettique et des altérations de la transduction du signal cellulaire[7].
En temps normal, les cellules disposent de mécanismes suffisants de défense et réparation contre les dommages causés par les ERA et les ROS, grâce à certaines enzymes. Ces mécanismes sont cependant dépassés en contexte de stress ou de pollution quand ces dérivés réactifs dépassent certains seuils de tolérance, ce qui peut conduire à la mort cellulaire et à des risques accrus de certaines maladies, notamment quand ces espèces chimiques très réactives peuvent en effet négativement interférer avec l'homéostasie cellulaire et de l'organisme, contribuant comme les espèces protéines, l'ADN et les membranes des cellules (attaque des lipides constitutifs par peroxydation lipidique) : c'est une des théories actuelles du vieillissement (sénescence).
Des conséquences génétiques ou épigénétiques sont également possibles dans les cellules germinales sont affectées.
Chez les animaux
Fonctions biologiques
Les macrophages synthétisent et utilisent cette espèce chimique très réactive (tout comme les ROS, espèces réactives de l'oxygène) pour tuer les microbes. Ces molécules sont alors dérivées de l'oxyde nitrique (•NO) et du superoxyde (O2•-) respectivement produites via l'activité enzymatique de l'oxyde nitrique synthase 2 inductible (NOS2) et la NADPH oxydase.
NOS2 est surtout exprimé dans les macrophages après induction par des cytokines et des produits microbiens, notamment l'interféron-gamma (IFN-γ) et le lipopolysaccharide (LPS)[9].
Les espèces azotées réactives agissent facilement synergiquement avec les espèces réactives de l'oxygène (ROS) pour endommager les cellules. On parle alors de « stress nitrosatif ».
Cette synergie fait que les deux espèces sont souvent cirées comme ROS/RNS ou EROA (espèces réactives de l'oxygène et de l'azote) ou RONS pour les anglophones (RONS signifiant Reactive oxygen and nitrogen species).
Chez les végétaux
Des espèces azotées réactives sont également produites en continu dans les plantes en tant que sous-produits du métabolisme aérobie, mais aussi en réponse à un stress[10].
Notes et références
- (en) Erica Novo et Maurizio Parola, « Redox mechanisms in hepatic chronic wound healing and fibrogenesis », Fibrogenesis & Tissue Repair, vol. 1, no 1, , p. 5 (ISSN 1755-1536, PMID 19014652, PMCID PMC2584013, DOI 10.1186/1755-1536-1-5, lire en ligne, consulté le )
- (en) Mudasir Irfan Dar, Mohd Irfan Naikoo, Fareed Ahmad Khan et Farha Rehman, « An Introduction to Reactive Oxygen Species Metabolism Under Changing Climate in Plants », dans Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress, Springer Singapore, , 25–52 p. (ISBN 978-981-10-5253-8, DOI 10.1007/978-981-10-5254-5_2, lire en ligne)
- Guzik TJ, West NE, Pillai R, Taggart DP, Channon KM, « Nitric oxide modulates superoxide release and peroxynitrite formation in human blood vessels », Hypertension, vol. 39, no 6, , p. 1088–94 (PMID 12052847, DOI 10.1161/01.HYP.0000018041.48432.B5)
- Squadrito GL, Pryor WA, « Oxidative chemistry of nitric oxide: the roles of superoxide, peroxynitrite, and carbon dioxide », Free Radical Biology and Medicine, vol. 25, nos 4–5, , p. 392–403 (PMID 9741578, DOI 10.1016/S0891-5849(98)00095-1)
- Dröge W, « Free radicals in the physiological control of cell function », Physiological Reviews, vol. 82, no 1, , p. 47–95 (PMID 11773609, DOI 10.1152/physrev.00018.2001, CiteSeerx 10.1.1.456.6690)
- O'Donnell VB, Eiserich JP, Chumley PH, Jablonsky MJ, Krishna NR, Kirk M, Barnes S, Darley-Usmar VM, Freeman BA, « Nitration of unsaturated fatty acids by nitric oxide-derived reactive nitrogen species peroxynitrite, nitrous acid, nitrogen dioxide, and nitronium ion », Chem. Res. Toxicol., vol. 12, no 1, , p. 83–92 (PMID 9894022, DOI 10.1021/tx980207u)
- Pacher P, Beckman JS, Liaudet L, « Nitric oxide and peroxynitrite in health and disease », Physiol. Rev., vol. 87, no 1, , p. 315–424 (PMID 17237348, PMCID 2248324, DOI 10.1152/physrev.00029.2006)
- (en) Huiying Li, C. S. Raman, Charles B. Glaser et Eric Blasko, « Crystal Structures of Zinc-free and -bound Heme Domain of Human Inducible Nitric-oxide Synthase: Implications for dimer stability and comparison with endothelial nitric-oxide synthase », Journal of Biological Chemistry, vol. 274, no 30, , p. 21276–21284 (ISSN 0021-9258 et 1083-351X, DOI 10.1074/jbc.274.30.21276, lire en ligne, consulté le )
- (en) Nicole M. Iovine, Seema Pursnani, Alex Voldman et Gregory Wasserman, « Reactive Nitrogen Species Contribute to Innate Host Defense against Campylobacter jejuni », Infection and Immunity, vol. 76, no 3, , p. 986–993 (ISSN 0019-9567 et 1098-5522, PMID 18174337, PMCID PMC2258852, DOI 10.1128/IAI.01063-07, lire en ligne, consulté le )
- (en) N. Pauly, « Reactive oxygen and nitrogen species and glutathione: key players in the legume-Rhizobium symbiosis », Journal of Experimental Botany, vol. 57, no 8, , p. 1769–1776 (ISSN 0022-0957 et 1460-2431, DOI 10.1093/jxb/erj184, lire en ligne, consulté le )