Colonisation assistée
La colonisation assistée, ou migration assistée, est une stratégie utilisée en biologie de la conservation qui consiste en l’introduction intentionnelle d’espèces animales et végétales dans un milieu se trouvant en dehors de leurs aires de répartition originelles[1].
Ce processus de conservation s’intègre dans un contexte de perte de biodiversité et de changement climatique. La colonisation assistée a pour but de littéralement venir en aide à ces espèces en les déplaçant volontairement dans un nouveau milieu qui correspondra sur le long terme à la niche écologique de l’espèce.
Contrairement à d’autres méthodes de conservation qui ont pour but la réintroduction des espèces au sein de leurs aires de répartition naturelle, la colonisation assistée présente des risques conséquents liés au fait que les espèces ou variétés déplacées n’étaient pas primitivement présentes dans le milieu colonisé. Certaines mesures visent à réduire ces risques, qui peuvent par exemple engendrer des proliférations indésirables (problématiques d’espèces invasives) ou de la compétition due au recouvrement de niches écologiques d’espèces déjà présentes sur le site d’introduction par l’espèce colonisatrice.
La colonisation assistée est aussi parfois utilisée à des fins économiques[2] (cf. exemple sur les langoustes de Tasmanie).
La migration assistée reste majoritairement théorique, à part pour quelques exemples d’applications[3]. Ceci est dû au fait que cette stratégie de conservation est un sujet de débat dans le domaine de l’environnement et de la conservation. Un consensus sur son acceptabilité n’a pas encore été atteint.
Contexte
En raison de l’urbanisation, de la fragmentation des habitats, de l’expansion de l’agriculture, ou encore de pollutions, la capacité des espèces à survivre dans leurs habitats naturels peut être menacée. On observe déjà aujourd’hui de nombreuses modifications dans l’aire de répartition de certaines espèces, cependant toutes ne sont pas capables de disperser ou de s’adapter suffisamment vite pour faire face au changement climatique et aux pressions anthropiques[4].
Prépondérance du changement climatique
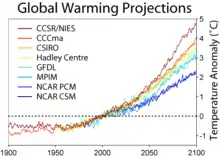
Le moteur de la conceptualisation de la colonisation assistée a d’abord été la prise en compte du changement climatique par les conservationnistes, et l’anticipation des menaces pouvant entraîner certains taxons vers un vortex d'extinction. Le débat persiste sur les capacités de certaines espèces à s’adapter au réchauffement global, notamment en raison des barrières naturelles et anthropiques à la migration.
En effet, celui-ci entraîne des modifications de conditions de vie. Ces modifications affectent souvent les températures, ainsi que les cycles hydrologiques locaux. Par exemple, « au Canada, les limites climatiques de nombreuses essences forestières devraient se déplacer d’environ 300 km vers le nord d’ici 2060[5]. » On s’attend notamment à ce que la composition des forêts québécoises soit modifiée : en effet, selon un mémoire publié par le ministère des Ressources naturelles du Québec[6], 14 des 49 espèces arborescentes présentes dans la province pourraient être désavantagées par la nouvelle réalité climatique d’ici 2100. Le changement climatique est donc en train de déplacer et de réduire la taille des enveloppes climatiques de plusieurs espèces. La question se pose aussi pour les espèces qui suivent la montée en altitude pour compenser l’augmentation de température, et qui se retrouvent parfois en quelque sorte cernées par les reliefs.
Menaces sur les espèces
Le potentiel d’impact se situe de l’ordre du gène et des organismes, à celui des populations, des communautés, et des écosystèmes[7]. Ce phénomène rend les individus, les espèces, et les communautés plus vulnérables aux espèces envahissantes, aux épidémies, et au parasitisme[5]. Ce qui, ensuite, crée des stresses environnementaux pour lesquelles certaines espèces végétales et animales ne sont pas adaptées[5]. De plus, si les stresses perdurent, les espèces vulnérables risqueraient de ne plus être en mesure de se régénérer[5].
À force de stress, les espèces les plus vulnérables risquent d’atteindre des extinctions locales, qui ont le potentiel de devenir globale, privant d’autres espèces de ressources ou de niches, et entraînant des dérèglements dans les dynamiques écologiques de leur écosystème. Ces dérèglements pourraient engendrer une perte de fonctions écologiques, qui affecterait la santé et la productivité des forêts, ainsi que la biodiversité locale[5]. Ces dérèglements mèneraient ainsi à une modification du paysage. Des forets pourraient devenir des savanes[8], qui pourraient finalement devenir des déserts.
Cependant, en parallèle, le changement climatique crée des espaces propices à la survie de ces espèces ailleurs. Ces nouveaux espaces peuvent se situer en bordure de leur espace de distribution habituel, tout comme ils peuvent se trouver à plusieurs kilomètres ou sur d’autres continents, hors de portée pour une dispersion naturelle[7].
Réponses
Face à ces nouvelles conditions, les espèces impliquées devraient assurer leur survie en migrant et colonisant de nouveaux espaces. Mais ce stratagème n’est pas nécessaire pour tous. Les pressions environnementales ont toujours su être le vecteur de l’évolution naturelle, et certaines espèces possèdent le potentiel de s’adapter aux nouvelles conditions climatiques sans avoir à quitter leur écosystème actuel. Ce potentiel varie d’une espèce à une autre. Entre autres, une étude a déterminé que l’évolution de la tolérance à de plus hautes température parmi certaines espèces d’amphibiens et de reptiles permettrait probablement à ces espèces de survivre à une augmentation en température de 3 °C sur 100 années[9]. Cependant, le rythme d’évolution du changement climatique dépasse celui de l’évolution naturelle de la majorité des espèces et des communautés, notamment les espèces végétales[5]. Par exemple, au Québec, le rythme de migration des espèces arborescentes est évalué à 100 à 200 m/an, tandis que la vitesse de déplacement du climat est estimée à 2 000 à 10 000 m/an[10].
Les espèces et les populations d’arbres forestiers devraient donc migrer plus rapidement que leur capacité naturelle. Par conséquent, les stratégies d’adaptation au changement climatique, telles que la migration assistée, ont retenu l’attention depuis 2007[11]. Les espèces peuvent être exportées vers un habitat offrant un avenir climatique plus propice à leur survie[5] ; ou inversement, des espèces sont importées pour leur diversité génétique déjà adaptée à un climat plus chaud, afin de maintenir la productivité forestière locale[10].
La migration assistée est donc une mesure préventive face au changement climatique, qui altère les communautés et les conditions environnementales, et qui se fait à une vitesse trop rapide pour l’écosystème[12].
Des biologistes peuvent aussi implémenter des espèces exotiques adaptées aux nouvelles conditions abiotiques, et capable de remplacer les fonctions écologiques des espèces locales stressées[10]. De cette manière les dynamiques de l’écosystème restent relativement stables, et l’écosystème peut conserver sa valeur ».
Terminologie
Brian J. Keel est le premier à utiliser le terme de « migration assistée » dans sa thèse en 2007. Il plaide en faveur de l’utilisation de la migration assistée comme stratégie de conservation contre le changement climatique rapide :
« In these situations where plants are prevented from migration, human intervention will be necessary to prevent extinction. I call this intervention “assisted migration”, which is the employment of techniques to ensure the maintenance of plant populations in a changing global environment through the intentional creation of populations beyond the boundary of a plant’s present range. »[13]
Cependant, la terminologie du terme employé par Keel est débattue car il peut prêter à confusion. En effet, en écologie le terme « migration » fait référence à un phénomène de déplacement périodique d’une espèce d’un endroit à un autre. Cette définition est en contradiction avec celle de la colonisation assistée, dans laquelle une espèce est déplacée durablement et artificiellement, en dehors de son aire de répartition originelle, afin de fonder une nouvelle population.
Types d’application
La migration assistée a le potentiel d’être appliqué aux espèces végétales et animales. Cependant, elle est plus souvent appliquée aux espèces végétales. Les animaux ont la capacité de se déplacer pour atteindre des espaces plus propices à leur survie, afin de trouver plus de nourriture, et de meilleure niche écologique[9]. Ils peuvent aussi se reproduire plus rapidement, en exprimant des mutations sélectionnant les plus adaptées[9]. Les espaces végétales ne possèdent ces capacités, et leur rythme d’évolution ne peut entrer en compétition avec le rythme de modification des caractéristiques abiotiques et biotiques de leur environnement[9] ; elles ont donc plus de besoins d’assistances.
La migration assistée offre trois types de procédés. La réintroduction, l’introduction, et l’augmentation[14].
Les habitats cibles peuvent être une expansion de leur habitat originel, un habitat duquel ils ont connu une baisse d’effectif ou une extinction locale, ou un nouvel habitat qui n’a jamais soutenu cette espèce[15]. Dans tous les cas, l’habitat cible est caractérisé par des conditions propices à la survie de l’espèce.
L’augmentation
L’augmentation a lieu lorsque la population atteint une densité trop basse pour assurer la survie de l’espèce via la reproduction, tout en risquant divers aléas génétiques tels que la consanguinité et l’homozygotie[14]. De nouveaux individus de la même espèce sont introduits afin d’augmenter la diversité génétique et l’accumulation d’individus.
Cependant, la présence de membres de la même espèce ne garantit pas que le nouvel habitat va convenir aux individus introduits, surtout si le manque d’effectif est causé par une baisse de qualité de l’environnement. Dans ce cas-ci, le nouvel habitat empêcherait l’établissement efficaces des nouveaux individus, et l’augmentation aurait de fortes chances d’être contre-productive[16].
La réintroduction
La réintroduction consiste en l’importation d’une espèce dans un écosystème dans lequel il habita, mais en fut extirpé[16]. Puisque cet environnement l’a supporté dans le passé, on peut émettre l’hypothèse qu’il serait propice à l’établissement de l’espèce. Cependant, comme précédemment, des facteurs sont à considérer[16]. Avant de pratiquer ce stratagème il est important de s’assurer que la cause de l’extinction locale a été traitée. Il est aussi important de s’assurer de l’authenticité des documents décrivant l’espèce comme ayant appartenu autrefois au site de réintroduction. Finalement, il faut aussi s’assurer que l’habitat n’a pas subi de modification depuis le déclin de l’espèce[16].
L’introduction
Dans le cas de l’introduction, une espèce est introduite dans un écosystème dans lequel elle n’a jamais habité. Ce procédé peut être exercé pour plusieurs raisons, tels que le biocontrôle d’une espèce envahissante[14], la perpétuité de l’espèce, ou le remplacement d’une espèce disparue localement[16]. Ce procédé est plus sensible, car l’impact de l’implantation est beaucoup plus difficile à prévoir. Si le sujet est mal étudié, il peut entraîner des désastres écologiques pour l’écosystème hôte[16].
Enjeux
Ce concept offre des bénéfices intéressants, tels que la prévention de l’extinction d’espèces ou de génotypes, la préservation de services écosystémiques, ou la préservation de certaines industries, comme la foresterie[7]. Mais : « l’invasion d’essences déplacées dans un nouvel écosystème, l’introduction de nouveaux organismes nuisibles et maladies, l’échec de la translocation et la collecte excessive de semences ou de boutures provenant de populations sources »[5] posent problèmes. D’autres négateurs soutiennent qu’il devrait y avoir une limite à l’implication humaine, qui a causé des catastrophes dans le passé[15].
Échec de projet
La stratégie serait considérée comme un échec dans le cas d’un projet, si l’espèce introduite ne parvient pas à survivre dans le nouvel écosystème. Cela pourrait être dû à une mauvaise planification, ou à un stress non-considéré[15]. Il n’est pas facile d’acquérir toutes les connaissances concernant une espèce et ses interactions avec son habitat, ce qui rend l’étude des stress d’une migration difficile à estimer. Il pourrait être causé par le transport des individus, une désorientation durant la mise en liberté, ou un délai entre la mise en liberté et les premières périodes de reproductions[15].
Une autre cause d’échec consisterait en un effort effectué inutilement à la suite d'une mauvaise interprétation des prévisions d’évolutions des niches climatiques. En effet, le fait qu’une espèce soit considéré comme étant vulnérable au changement climatique à cause d’une perte de surface de distribution ne signifie pas que la migration assistée est la stratégie propice à son cas. Les conditions géographiques doivent d’abord être prises en considérations pour évaluer la combinaison des pertes et des gains en surface[3]. Une espèce qui ne perd pas d’habitat adéquats ne nécessite pas d’implication humaine. Une espèce qui ne possédera pas de nouveaux espaces adéquats hors de son champ de distribution actuelle ne nécessitera pas d’aide migratoire. Seule une espèce qui gagnerait, par le changement climatique, une quantité non négligeable de territoires adéquats, tout en perdant une quantité importante de territoires actuel, et en ayant des difficultés de dispersions est éligible à une stratégie de migration assistée[3].
Espèces envahissantes
Un des contre-arguments les plus fréquemment associés à la migration assistée, est le fait que les espèces implémentées ont le potentiel de se comporter comme des espèces envahissantes. Une espèce envahissante est une espèce exotique venant d’un écosystème, et s’implémentant dans un nouveau. Les espèces envahissantes ont la caractéristique de « trop bien s’adapter », et de devenir dominantes dans le nouvel écosystème, au détriment des espèces natives[15].
Cependant, les espèces envahissantes ont souvent été la cause d’une introduction accidentelle. Un plan de stratégie incluant la migration assistée devrait faire l’objet d’une étude exhaustive, et prendre en compte ce risque en particulier. De ce fait, ce risque devrait présenter un potentiel minime avant d’entreprendre un tel projet[15]. De plus, une étude a mentionné que moins de 1 % des espèces ayant été introduites dans un nouvel environnement sont devenues envahissantes[17].
Hybridation
L’hybridation représente un autre contre-argument. L’hybridation a lieu lorsque l’espèce introduite est assez proche d’au moins d’une des espèces locales pour former des hybrides[1]. Une répercussion de cette hybridation est la dilution génétique et l’assimilation, qui causerait l’extinction de certaines espèces. Il est estimé que l’hybridation serait responsable d’au moins 31 % des extinction d’espèces de poissons d’eau douce en Amérique du Nord[1].
Le poids des risques et des bénéfices
Le changement climatique a le potentiel de causer la perte de plusieurs espèces, et que les méthodes habituelles de conservation pourraient ne pas protéger les espèces d’un déclin éventuel[3]. Les environnementalistes craignant les risques de la migration assistée proposent des méthodes alternatives, tels que la trame verte, ou le couloir écologique, afin de connecter différents habitats[18]. Cependant, la vitesse de ce changement étant grandissante, des habitats connectés pourraient ne pas être suffisants face à cette menace. Le principe de précaution est aussi appelé face à la stratégie de la migration assistée[15]. Cependant, la situation est trop délicate pour se baser sur les stratégies actuelles. Il est donc nécessaire de considérer la migration assistée, mais uniquement sous des conditions bien étudiées, et avec un plan de suivi sur plusieurs années.
Trois points de vue principaux sur l’utilisation de la colonisation assistée existent au sein de la communauté scientifique:
- La colonisation assistée est nécessaire et doit inclure un déplacement extensif d’espèces au-delà de leurs distributions actuelles. Les partisans de cette approche soutiennent que la colonisation assistée est une action nécessaire pour lutter contre le changement climatique et les perturbations dues à l’activité humaine.
- La méthode précédente est trop agressive. Les partisans prônent une approche permettant d’équilibrer risques et avantages de la colonisation assistée.
- La colonisation assistée doit être évitée. Les défenseurs de cette approche considèrent que les connaissances scientifiques actuelles et la limitation des technologies disponibles ne permettent pas de prédire correctement l’impact et les effets à moyen terme de l’assistance à la colonisation. Les défenseurs de cette thèse estiment que les risques de mise en place de la colonisation assistée sont trop élevés compte tenu des difficultés à prévoir les régions cibles, du manque de données disponibles qui permettraient de modéliser les enveloppes climatiques de la plupart des espèces, et des incertitudes dans les prévisions climatiques.
Conditions et méthodes d’application
Selon les directives IUCN[19] - [20], la colonisation assistée ne doit être utilisée qu’en dernier recours, par exemple dans des cas extrêmes afin d’éviter l’extinction d’une espèce ou lorsqu’il ne reste plus aucune zone habitable dans l’aire de répartition naturelle de l’espèce. En conséquence, seules les espèces dont la probabilité d’extinction est importante, ou les espèces clefs de voûte (celles ayant un rôle écologique essentiel) ainsi que les espèces peu résilientes sont concernées par cette méthode.
Aujourd’hui, une approche relevant du champ de l’écologie fonctionnelle tend à s’imposer. En effet la colonisation assistée peut recevoir une priorité plus élevée dans les stratégies d’adaptation au changement climatique si les taxons déplacés remplissent des fonctions écologiques essentielles, plutôt que d’avoir peu d’avantages collatéraux, ou même des effets néfastes. (Cf exemples)
La gestion des populations déplacées après la colonisation nécessitera un suivi régulier, mais l’intervention humaine est la première étape pour aider les espèces à se déplacer à mesure que le climat change.
Outre les effets physiologiques directs sur les organismes et les modifications de la condition physique, le changement climatique affectera de nombreuses espèces par le biais d’impacts indirects sur la structure, les fonctions et les processus de l’écosystème. Les changements dans l’abondance des espèces dominantes, des espèces de base et des espèces clés modifieront ces processus, ce qui affectera de nombreux organismes associés.
L’examen des stratégies d’adaptation aux changements climatiques fournit un large éventail d’approches pour le maintien de processus écologiques, telles que le cycle des éléments nutritifs, l’hydrologie, les interactions entre espèces, la fourniture d'habitat, la dispersion et les perturbations. Des écologistes du paysage et de la restauration ont ainsi proposé que la végétation soit rétablie et restaurée aux niveaux locaux et régionaux pour améliorer les processus et les fonctions écologiques susceptibles de maintenir la biodiversité. Mais le réchauffement global altère aussi les communautés végétales. La relocalisation des taxons peut alors être entreprise pour assurer des fonctions écologiques directement affectées par le changement climatique ou lorsqu’il exacerbe d’autres causes de déclin, telles que la fragmentation ou la salinisation.
Par exemple, considérons une espèce d’arbre qui est en déclin et qui fournit des nids à une communauté d’oiseaux. L’approche fonctionnelle vise à trouver quelles espèces d’arbres pourraient persister dans la région à l’avenir pour fournir un habitat de nidification aux oiseaux, et pourrait inclure l’option d’introduire plusieurs espèces, si plusieurs espèces sont capables de remplir la fonction souhaitée.
Néanmoins, il est nécessaire de comprendre les stratégies adaptatives pour les espèces confrontées à des changements de climat, par des expériences de transplantation à grande échelle. Ces expériences prennent du temps et sont coûteuses, mais elles seront essentielles à la mise en place d’un programme efficace de migration assistée.
L’idiosyncrasie de la biologie des espèces empêche l’élaboration de plans de migration assistée adaptés à chacune des nombreuses espèces susceptibles d’être menacées par le changement climatique. Par conséquent, les gestionnaires seront obligés de généraliser les stratégies prévues en fonction de grandes catégories de caractéristiques du cycle vital. On peut imaginer que les futurs « biologistes de la migration assistée » se retrouvent dans une position similaire à celle des biologistes des espèces envahissantes actuels : rechercher des généralisations théoriques et lutter avec des idiosyncrasies imprévues dans la pratique[21] - [22] - [23] - [24].
Plusieurs protocoles ont été mis au point lors d’étude portant sur l’application responsable de le migration assistée, afin de guider des décisions à propos des situations et des périodes adéquates pour la migration assistée, ainsi que pour la planification du projet s’il est décrété faisable[3].
Conditions géographiques
Selon la répartition des espaces d’origines, et des espaces potentiels, le potentiel d’action de la migration assistée peut varier. En effet, si les changements climatiques induisent une faible ou forte perte d’espace aux conditions adéquates dans l’espace d’origine, ainsi qu’un faible gain d’espace potentiel, les espèces ne gagneraient pas à être transportées ailleurs[3]. De même, si les changements climatiques causent une faible perte d’espace aux conditions adéquates dans l’espace d’origine, ainsi qu’un fort gain d’espace potentiel, la migration assistée ne devrait pas être appliquée[3].
Cependant, si une perte importante d’espace aux conditions adéquates dans l’espace d’origine et d’espaces propices est estimée, la migration assistée devient un candidat de stratégie de conservation d’espèce[3]. L’étude d’Hällfors (2017)[3] présente une figure représentant bien ces conditions.
Protocole
Une étude[17] sur la propagation d’espèces végétales à graines a élaboré un protocole à suivre avant de déterminer la possibilité d’appliquer la migration assistée à un projet de conservation. Le protocole est le suivant.
Développement d’une liste d’espèces de base
Lors de cette étape, l’importance est portée sur la collecte d’une banque de donnée sur les graines présentes sur le site, ainsi que leur localisation. Plusieurs banques de données ont déjà été établies. Le Millennium Seed Bank Project des Jardins botaniques royaux de Kew a pour but de collectionner et mettre en banque les graines de 35 % des plantes mondiales[17].
Cela commence par la compilation d’une liste des espèces porteuses de graines sur le lieu investit, suivi de l’élimination d’espèces hybrides ou non-native, l’élimination d’espèce à graines récalcitrantes, et la hiérarchisation des espèces selon leur importance dans la communauté.
La compilation nécessite un protocole de collection de graine assez minutieux et précis qui comprend :
- la capture de 95 % de la diversité génétique de chaque espèce ;
- collection restreinte, qui laisse des graines sur le terrain qui pourront germer et produire les générations futures ;
- collection sur plusieurs années, afin d’avoir un minimum de 3 000 graines, en gardant un but optimal de 30 000 ;
- collection de graines toujours matures ;
- collection représentant les gradients de densité du site ;
- les coordonnées GPS associées à chaque collection.
Espèces importantes à la restauration
Au cours de cette étape, on détermine quelles espèces sont importantes pour la restauration du site. En utilisant la liste résultant de la première étape, on identifie les espèces pionnières non-agressive qui sont utilisées en restauration, les espèces les plus conservatives qui sont souvent utilisées dans des projets de restauration, et les espèces non-agressives qui sont aux lisières de l’espace.
Espèces avec un bénéfice potentiel pour la migration assistée
Après la deuxième étape, on priorise la liste selon les espèces qui ont :
- des risques géographiques ;
- des contraintes dans leur histoire de vie ;
- des limites de dispersion ;
- un taxon très conservateur ;
- des distributions très fragmentées ;
- une abondance restreinte à l’emplacement étudié.
Détermination des priorités de collection au niveau de la population.
Au cours de cette étape, on crée un modèle de distribution à l’aide d’un système d’information géographique pour espèces dans les lisières du site, dans les zones en dehors des futures estimations climatiques, et dans les zones non-protégées.
Implémentation de la stratégie de migration assistée
Au cours de cette etape, on peut entreprendre un protocole de correspondance d’habitat à l’aide du système d’information géographique.
Exemples
Exemples de succès écologiques
En tant que méthode de conservation controversée, la colonisation assistée n’est encore qu’à ses débuts, de nombreuses études sont menées par la communauté scientifique, dont voici quelques exemples. Cette liste non exhaustive reprend des études menées chez différents taxons et avec des motivations écologique ou économique.
Les Pins du Mexique
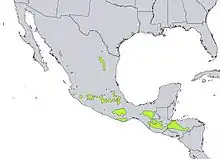
Les changements climatiques globaux vont perturber l’équilibre établi entre les populations forestières et le climat pour lequel ils sont adaptés, pouvant induire une détérioration progressive des forêts engendrant ainsi des conséquences écologiques et économiques.
Afin d’anticiper ce phénomène, en 2015, une équipe de recherche a expérimenté la colonisation assistée sur trois espèces de Pins — Pinus pseudostrobus, Pinus leiophylla et Pinus devoniana — dans les montagnes du Mexique[26].
Naturellement, ces trois espèces sont réparties le long d’un gradient d’altitude entre 2 900 m et 1 650 m, avec la dominance, de haut en bas, de P. pseudostrobus, puis de P. leiophylla et enfin de P. devoniana[26]. À la suite d'une colonisation assistée ascendante de 300 mètres d’altitude (plantation de pins entre 3 200 et 1 950 m), la survie moyenne chez les trois espèces est de 79,6 %, avec une différenciation génétique notable, majoritairement due à la pression sélective exercée par les variations de températures. Cette diversité génétique traduit l’adaptation des pins à leur nouvel environnement, par sélection naturelle.
Les Papillons d’Angleterre

Entre 2000 et 2008, une étude a été menée sur la colonisation assistée de deux espèces de papillons britanniques : Melanargia galathea et Thymelicus sylvestris au nord de l’Angleterre[27].

Les deux espèces ont été introduites, respectivement, à 65 km et 35 km au nord de leur aire de répartition naturelle, dans des zones respectant leur niche écologique respective. Le suivi des populations au sein des sites d’introduction a permis de montrer que, chez les deux espèces, le taux de croissance de la population assistée était équivalent à celui d’une population naturelle. De plus, entre 2000 et 2006, les individus se sont dispersés, augmentant l’aire de distribution de M. galathea de 7,2 à 17,8 ha et celle de T. sylvestris de 0,17 à 3,64 ha[27].
Ainsi, dans ce cas d’étude, la colonisation assistée semble être un succès, avec le maintien des populations après huit générations (2000-2008) et l’élargissement de leur aire de répartition.
Les Langoustes de Tasmanie
Entre 2005 et 2007, une expérience a été menée dans le sud de la Tasmanie, visant à augmenter la production issue de la pêcherie de la langouste australe : Jasus edwardsii, tout en augmentant la résilience de ces populations face aux changements climatiques[2].

Ainsi, 5 747 langoustes femelles matures ont été déplacées depuis Maatsuyker Islands (en) vers deux sites à des latitudes supérieures : Riedle Bay (ceb) et Taroona Reserve (en), respectivement à 185 km et 120 km de Maatsuyker Island. Ce changement d’environnement (température, salinité de l’eau), mais également le type, l’abondance et la qualité des proies ainsi que la disponibilité et le type de refuges ont affecté le cycle de vie des langoustes.
Ainsi, en réponse au stress environnemental, les langoustes ont optimisé l’allocation d’énergie à la croissance au détriment de la reproduction (notion de trade-off). En effet, la première année, la croissance de J. edwardsii est passée de 3 à 6 mm par an tandis que 50 % ne se sont pas reproduit cette année[2]. À partir de la deuxième année, 93 % des femelles qui ne se sont pas reproduites la première année, ont recommencé à se reproduire, tandis que leur taux de croissance a régressé jusqu’à revenir identique à celui des langoustes de Maatsuyker Islands. Ces femelles ont augmenté leur production d’œufs de 65 000 œufs par an.
Cette expérience montre le potentiel de la colonisation assistée comme outil pour améliorer la production, la durabilité et la résilience de la pêche. En effet, malgré une maturité sexuelle tardive, en réponse à un stress environnemental, la croissance plus importante des langoustes permet une production d'œufs, plus importante.
Exemples d’effets indésirables
Malgré l’objectif de la conservation de la biodiversité, la connaissance partielle d’un écosystème et l’insuffisante prise en compte de toutes les interactions écologiques au sein de l’écosystème peut entraîner de nombreux effets indésirables sur la biodiversité. Ainsi, la colonisation comporte de nombreux risques comme apporter de nouveaux agents pathogènes, créer une nouvelle compétition pour les ressources, bouleverser les interactions écologiques ou encore diminuer la diversité locale en favorisant l’hybridation[28].
Par précaution, lorsque l’écosystème n’est pas compris dans sa globalité et que tous les potentiels effets d’une colonisation assistée ne sont pas connus, il est aujourd’hui conseillé d’utiliser les mesures de conservation existantes et d’atténuer l’impact de l’activité humaine sur l’écosystème[28], afin d’éviter tout effet indésirable pouvant amener à des catastrophes écologiques.
Les Crevettes d’Amérique du Nord

Mysis relicta (en) est une crevette d’eau douce, d’une taille comprise entre 1 et 2 centimètres de long, originaire d’un nombre limité de lacs d’Amérique du nord et de Suède. En 1949, M.relicta été introduite dans un lac du Canada (Kootenay Lake) afin d’augmenter les populations de truites arc-en-ciel (Oncorhynchus mykiss), en déclin[29]. L’expérience fut un succès, entraînant une importante augmentation de la taille et du nombre de truites.
Ces résultats ont encouragé, entre 1968 et 1975, les gestionnaires de la faune du Flathead Lake (Montana, USA) à introduire M. relicta afin d’augmenter les populations d’une autre espèce de saumon (Oncorhynchus nerka) en améliorant leur régime alimentaire. Malheureusement, la crevette vit la journée au fond du lac et remonte la nuit dans les eaux de surface, tandis que le saumon se nourrit la journée dans les eaux peu profondes[29]. Contrairement au lac du Canada (Kootenay Lake), l’absence de courants verticaux (upwelling) rend les crevettes inaccessibles pour le saumon.
.jpg.webp)
À défaut de devenir des proies, les crevettes sont devenues des concurrentes pour les ressources alimentaires du lac, entraînant l’effondrement de la population de saumon rouge.
La disparition du saumon rouge a eu nombreuses répercussions sur tout le réseau trophique. Ainsi, les populations d’aigles (Haliaeetus leucocephalus), de coyotes (Canis latrans), de visons (Neovison vison), de loutre de rivière (Lutra canadensis), de grizzly (Ursus arctos) et même de cerf de Virginie (Docoileus virginianus) ont été affectées par cette perte de ressource alimentaire.
Il est également notable que la dégradation de la biodiversité a eu un impact économique, puisque le nombre de visiteurs annuel est passé de 46 500 en 1983, à moins de 1 000 en 1989[29].
Cette étude démontre, avec du recul (colonisation assistée dans les années 1970), que chaque écosystème est unique et doit être considéré comme tel. Dans ce cas d’étude, un succès de restauration (Kootenay Lake) s’est transformé en désastre écologique (Flathead Lake) à cause d’un seul facteur non considéré (absence de courants verticaux).
Les Écureuils de Terre-Neuve


Afin de sauver la population de martre d’Amérique (Martes americana) en déclin sur l’île de Terre-Neuve, l’écureuil roux d’Amérique (Tamiasciurus hudsonicus) a été introduit en 1963. L’objectif de cette colonisation assistée étant de favoriser la survie et la reproduction du martre en enrichissant son régime alimentaire[28].
Mais, les effets ne furent pas ceux escomptés, puisque la martre est toujours menacée sur l’île de Terre-Neuve[30].
En revanche, la population d’écureuils roux a grandement proliféré au sein des forêts de conifères. La densité d’écureuils sur l’île est deux fois supérieure à la densité d’écureuils continentaux. Ceci provoque une forte compétition à la faune locale pour les ressources alimentaires : en dévorant les cônes de sapin baumier (Abies balsamea), l’écureuil a provoqué la disparition, en 1988, du bec-croisé des sapins (Loxia curvirostra) qui se nourrissait également de ces cônes[28].
Voir Aussi
https://www.rncan.gc.ca/forets/changements-climatiques/13084
https://www.rncan.gc.ca/forets/changements-climatiques/adaptation/13122
Notes et références
- (en) Anthony Ricciardi et Daniel Simberloff, « Assisted colonization is not a viable conservation strategy », Trends in Ecology & Evolution, vol. 24, no 5, , p. 248–253 (ISSN 0169-5347, PMID 19324453, DOI 10.1016/j.tree.2008.12.006, lire en ligne, consulté le )
- (en) Peter J. Hawthorne, Adrian Linnane, Caleb Gardner et Bridget S. Green, « The Good, the Bad and the Recovery in an Assisted Migration », PLOS ONE, vol. 5, no 11, , e14160 (ISSN 1932-6203, PMID 21151965, PMCID PMC2996890, DOI 10.1371/journal.pone.0014160, lire en ligne, consulté le )
- (en) « Quantifying the need and potential of assisted migration », Biological Conservation, vol. 205, , p. 34–41 (ISSN 0006-3207, DOI 10.1016/j.biocon.2016.11.023, lire en ligne, consulté le )
- « Assisted Colonization and Rapid Climate Change », sur online.nwf.org (consulté le )
- J. H. Pedlar, D. W. McKenney et I. Aubin, « Comment la migration assisté pourrait-elle faciliter l’adaptation des forêts aux changements climatiques? Nouvelles Express 63. », Nouvelles Express, (lire en ligne, consulté le )
- Périé, C., S. de Blois, M.-C. Lambert et N. Casajus. 2014. Effets anticipés des changements climatiques sur l'habitat des espèces arborescentes au Québec. Gouvernement du Québec, ministère des Ressources naturelles, Direction de la recherche forestière. Mémoire de recherche forestière n° 173. 46 p. En ligne : https://mffp.gouv.qc.ca/publications/forets/connaissances/recherche/Perie-Catherine/Memoire173.pdf (consulté le 18 janvier 2022)
- I. Aubin, C.M. Garbe, S. Colombo et C.R. Drever, « Why we disagree about assisted migration: Ethical implications of a key debate regarding the future of Canada's forests », The Forestry Chronicle, vol. 87, no 06, , p. 755–765 (ISSN 0015-7546 et 1499-9315, DOI 10.5558/tfc2011-092, lire en ligne, consulté le )
- (en) « ‘Tipping points’ for the Amazon forest », Current Opinion in Environmental Sustainability, vol. 1, no 1, , p. 28–36 (ISSN 1877-3435, DOI 10.1016/j.cosust.2009.07.003, lire en ligne, consulté le )
- (en) DAVID K. SKELLY, LIANA N. JOSEPH, HUGH P. POSSINGHAM et L. KEALOHA FREIDENBURG, « Evolutionary Responses to Climate Change », Conservation Biology, vol. 21, no 5, , p. 1353–1355 (ISSN 0888-8892 et 1523-1739, DOI 10.1111/j.1523-1739.2007.00764.x, lire en ligne, consulté le )
- Lamhamedi, M. S, « L’écophysiologie, un Atout Pour Réussir la Migration Assistée de Sources Génétiques D’épinette Blanche », Avis de Recherche forestière,
- (en) Mary I. Williams et R. Kasten Dumroese, « Preparing for Climate Change: Forestry and Assisted Migration », Journal of Forestry, vol. 111, no 4, , p. 287–297 (lire en ligne, consulté le )
- (en) S. Trumbore, P. Brando et H. Hartmann, « Forest health and global change », Science, vol. 349, no 6250, , p. 814–818 (ISSN 0036-8075 et 1095-9203, PMID 26293952, DOI 10.1126/science.aac6759, lire en ligne, consulté le )
- (en) Brian J. Keel (Chapter 3 Assisted Migration - 28 p.), Assisted Migration as a Conservation Strategy for Rapid Climate Change: Investigating Extended Photoperiod and Mycobiont Distributions for Habenaria Repens Nuttall (Orchidaceae) as a Case Study, Antioch University New England, , 160 p. (lire en ligne)
- (en) Andrew R. Weeks, Carla M. Sgro, Andrew G. Young et Richard Frankham, « Assessing the benefits and risks of translocations in changing environments: a genetic perspective », Evolutionary Applications, vol. 4, no 6, , p. 709–725 (ISSN 1752-4571, PMID 22287981, PMCID PMC3265713, DOI 10.1111/j.1752-4571.2011.00192.x, lire en ligne, consulté le )
- (en) Bridget S. Green, Caleb Gardner, Adrian Linnane et Peter J. Hawthorne, « The Good, the Bad and the Recovery in an Assisted Migration », PLoS ONE, vol. 5, no 11, , e14160 (ISSN 1932-6203, PMID 21151965, PMCID PMC2996890, DOI 10.1371/journal.pone.0014160, lire en ligne, consulté le )
- (en) Philip J. Seddon, « From Reintroduction to Assisted Colonization: Moving along the Conservation Translocation Spectrum », Restoration Ecology, vol. 18, no 6, (ISSN 1526-100X, DOI 10.1111/j.1526-100x.2010.00724.x@10.1111/(issn)1526-100x.2525thanniversaryvi, lire en ligne, consulté le )
- (en) « Assisted migration of plants: Changes in latitudes, changes in attitudes », Biological Conservation, vol. 143, no 1, , p. 18–27 (ISSN 0006-3207, DOI 10.1016/j.biocon.2009.08.015, lire en ligne, consulté le )
- (en) MEADE KROSBY, JOSHUA TEWKSBURY, NICK M. HADDAD et JONATHAN HOEKSTRA, « Ecological Connectivity for a Changing Climate », Conservation Biology, vol. 24, no 6, , p. 1686–1689 (ISSN 0888-8892, DOI 10.1111/j.1523-1739.2010.01585.x, lire en ligne, consulté le )
- « Guidelines for Reintroductions and Other Conservation Translocations », sur portals.iucn.org (consulté le )
- (en) « IUCN Guidelines For Re-introduction », sur portals.iucn.org (consulté le )
- « Canadian Institute of Forestry », sur pubs.cif-ifc.org (DOI 10.5558/tfc2011-092, consulté le )
- (en) « Using assisted colonisation to conserve biodiversity and restore ecosystem function under climate change », Biological Conservation, vol. 157, , p. 172–177 (ISSN 0006-3207, DOI 10.1016/j.biocon.2012.08.034, lire en ligne, consulté le )
- (en) C. D. Thomas, H. P. Possingham, C. Parmesan et D. B. Lindenmayer, « Assisted Colonization and Rapid Climate Change », Science, vol. 321, no 5887, , p. 345–346 (ISSN 1095-9203 et 0036-8075, PMID 18635780, DOI 10.1126/science.1157897, lire en ligne, consulté le )
- (en) Jason S. McLACHLAN, Jessica J. Hellmann et Mark W. Schwartz, « A Framework for Debate of Assisted Migration in an Era of Climate Change », Conservation Biology, vol. 21, no 2, , p. 297–302 (ISSN 1523-1739, DOI 10.1111/j.1523-1739.2007.00676.x, lire en ligne, consulté le )
- Elbert L. Little, Atlas of United States trees., U.S. Dept. of Agriculture, Forest Service,, (lire en ligne)
- (en) D. Castellanos-Acuña, R. Lindig-Cisneros et C. Sáenz-Romero, « Altitudinal assisted migration of Mexican pines as an adaptation to climate change », Ecosphere, vol. 6, no 1, , art2 (ISSN 2150-8925, DOI 10.1890/ES14-00375.1, lire en ligne, consulté le )
- (en) Stephen G. Willis, Jane K. Hill, Chris D. Thomas et David B. Roy, « Assisted colonization in a changing climate: a test-study using two U.K. butterflies », Conservation Letters, vol. 2, no 1, , p. 46–52 (ISSN 1755-263X, DOI 10.1111/j.1755-263X.2008.00043.x, lire en ligne, consulté le )
- (en) Daniel Simberloff et Anthony Ricciardi, « Assisted colonization is not a viable conservation strategy », Trends in Ecology & Evolution, vol. 24, no 5, , p. 248–253 (ISSN 0169-5347, PMID 19324453, DOI 10.1016/j.tree.2008.12.006, lire en ligne, consulté le )
- Craig N. Spencer, B. Riley McClelland et Jack A. Stanford, « Shrimp Stocking, Salmon Collapse, and Eagle Displacement », BioScience, vol. 41, no 1, , p. 14–21 (ISSN 0006-3568 et 1525-3244, DOI 10.2307/1311536, lire en ligne, consulté le )
- « Rapport COSEPAC : Martre d’Amérique - Population de Terre-Neuve », (consulté le )
- Cet article est partiellement ou en totalité issu de l'article intitulé « Migration assistée » (voir la liste des auteurs).