Cycle sylvigénétique
Le cycle sylvigénétique (aussi appelé cycle sylvogénétique) est le cycle d'évolution naturelle d'une forêt sauvage (non exploitée par l'homme).
Cette succession écologique inclut des dynamiques successives d'évolution interne de la forêt et de ses milieux, partant d’un stade dit pionnier et tendant à aboutir à un stade dit climacique jusqu’à ce qu'une perturbation (chablis résultant de tempêtes, incendies, glissements de terrain, avalanches…) réintroduise les conditions d’expression du premier stade.

La sylvigenèse ou sylvogenèse désigne l'ensemble des processus dynamiques naturels s'observant au sein d'un écosystème forestier ou d'un massif forestier, relevant de la dynamique cyclique (cycle sylvigénétique) ou de la dynamique linéaire (reconquête forestière, reconstitution après une perturbation ayant ouvert de vastes espaces)[1].
Chez les arbres ; à chaque stade de la succession correspond un type dominant de reproduction, un potentiel d’expression des semences, et des stratégies adaptatives et colonisatrices différentes, mises en œuvre par des groupes fonctionnels d'essences spécifique dominantes.
Plusieurs facteurs écologiques peuvent expliquer les logiques dynamiques des premiers stades juvéniles de toutes les essences forestières ;
- Mise en lumière, modifications hydriques, biochimiques et physiques apportées par la perturbation (ex : apport de charbons de bois après un incendie de forêt[2])
- mécanismes de germination
- type de régénération
- type de réponses à la mise en lumière
...faisant émerger deux catégories d’arbres qu’on regroupe habituellement en
- essences (et espèces associées) de lumière (ou de milieux ouverts) qui ne se développent bien que dans les trouées, clairières et en plein lumière, au moins pour leur stade juvénile.
- essences (et espèces associées) d'ombre qui ne se développent qu’à couvert ou sous une lumière tamisée ; toute leur vie, ou au moins au stade juvénile.
Dans cette optique, on classe parfois les essences et leurs cortèges d’espèces associées selon un « tempérament (ou gradient) photique » juvénile des essences.
Succession spatiotemporelle
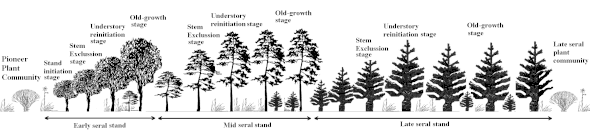
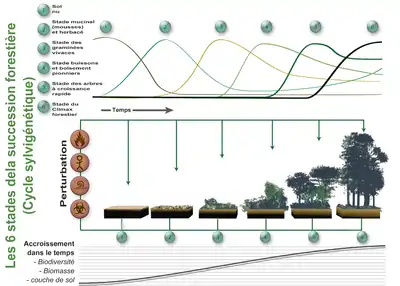
Après un certain temps survient une perturbation qui fait reprendre le "cycle" à son début (ou à un stade intermédiaire si la perturbation est peu importante).
Dans le bas de l'image est représentée l'accroissement de biomasse (sur pied et dans le sol, animale, végétale et fongique..) de biodiversité et d'épaisseur de sol (qui a une importance en termes de puits de carbone). Au fur et à mesure de cette succession, les communautés végétales (et les communautés microbiennes, fongiques et animales qui leur sont associées évoluent) en se remplaçant les unes les autres. Les projets de « forêt comestible » basés sur la foresterie analogue s'inspirent de ce cycle, mais en ciblant principalement des espèces d'intérêt alimentaire
.
La théorie sylvicole veut que différents stades se succèdent cycliquement pour constituer la sylvigenèse. En réalité - à échelle macroscopique - ces stades coexistent dans l'espace et le temps, au gré d'apparition des trouées dans la forêt.
Chacun de ces stades est caractérisé par des types particuliers de « stratégies » et adaptations évolutives lentement mises en place avec la sélection naturelle (ce sont par exemple des stratégies de compétition, exclusion, reproduction et interactions durables..), et par des comportements juvéniles plus ou moins adaptés au manque de disponibilité en eau, nutriments ou lumière (laquelle diminue au sol au fur et à mesure qu’une canopée dense et jointive se reconstitue).
De manière très simplifiée et théorique on présente ces stades comme suit :
Stade 1 : la trouée forestière ou unité de régénération naturelle
Les semis naturels d’espèces pionnières (ex: bouleau en zone tempérée de l'hémisphère nord), profitant du soleil et de l’eau qui n’est plus interceptée par la canopée, ni pompée par les racines, y croissent très vite. La sélection des plants et des espèces semble contrôlée par la compétition interindividuelle, mais de nombreuses symbioses et interactions complexes sont en jeu. Les bois morts commencent à être colonisés et décomposés par les bactéries, les champignons, les insectes et invertébrés qui s’en nourrissent, produisant un sol riche et favorables aux futurs arbres.
Invertébrés fouisseurs, oiseaux, chauve-souris et mammifères jouent un rôle important d’apports, de transport, d'enfouissement ou au contraire de mise à jour de graines enfouies dans le sol. Certaines graines ne germant qu’après être passées dans le tube digestif d’un animal.
À ce stade une première sélection se fait par l’occupation de l’espace aérien et racinaire, avec élimination d’un grand pourcentage des individus issus des graines qui ont pu s’exprimer. Les buissons et de premiers arbres pionniers (dominants et dominés) apparaissent.
Au fur et à mesure que les espèces pionnières (re)colonisent en successions et assemblages plus ou moins complexes[3] l’espace aérien et le sol (saproxylophages, invertébrés dont arthropodes[4], enchytraéidés[5] et lombrics[6] - [7]), l’hygrométrie et la qualité de l’humus évoluent, alors que l’ensoleillement direct du sol diminue, modifiant les conditions de vie ou d’apparition/disparition des espèces. Le tapis herbacé (dont légumineuses) après avoir intensément colonisé (enrichi et stabilisé) le sol, va peu à peu régresser à l’ombre des arbres et buissons.
À ce stade, les espèces consacrent beaucoup d’énergie dans la reproduction (semences abondantes, légères et fortement dispersées par le vent et l’eau, ou les oiseaux) et dans la compétition (via par exemple une maturité sexuelle précoce et une forte fécondité, ou en attirant des espèces animales qui vont favoriser leur dissémination à distance).
Certaines graines d’espèces pionnières ont une dormance levée par la mise en lumière, un incendie (température) ou simplement l’interruption de production d’hormones inhibitrices par les arbres antérieurement présents.
Les espèces puis essences pionnières sont toutes caractérisées par une croissance rapide, une taille plus modeste que celle des plus grands arbres et des durée de vie plus courte, ainsi que par l’héliophilie des semis et une germination facilité dans les milieux ouverts. Les buissons et arbres « pionniers » sont généralement à bois tendres et fragile (saule, bouleau, peuplier tremble, aulne ou cytise et dans certaines conditions le pin sylvestre) voire creux ou à moelle peu dense (ex : Bois-canon sous les tropiques, sureau en zone tempérée), prolixes en graines et pollens. Sous l’action des bactéries et champignons, leur bois mort se décompose très rapidement, contribuant à une production accélérée d’humus pré-forestier. Ces arbres poussent en bouquet qui créent en quelques années un premier microclimat forestier (plus ombreux et frais) favorable à l'installation d'essences dites secondaires, appartenant aux autres groupes fonctionnels. Leurs racines décolmatent et enrichissent les sols.
Stade 2 : espèces et essences secondaires ou post pionnières
Leur bois est un peu plus dense et durable. Elles vivent plus longtemps et nécessitent un moindre ensoleillement, mais croissent plus lentement. Elles sont souvent moins héliophiles ; ce sont par exemple en zone tempérée l’érable, le frêne, l’orme, le tilleul, le chêne, le merisier, le sorbier, le pin, le mélèze. Elles peuvent être accompagnées de lianes et d'épiphytes qui s'élèvent au rythme de la croissance de la forêt.
Stade 3 : apparition des dryades (phases de maturité)
Ce groupe apparaît lentement et devient dominant après un siècle ou deux, voire beaucoup plus en forêt tropicale et équatoriale.
Il réunit des arbres à grande longévité, souvent à racines profondes dans les forêts où l’eau manque, et à bois beaucoup plus dense et dur, et souvent sciaphiles (appréciant l'ombre et une humidité et fraîcheur relative), au moins au stade juvénile, bien que pouvant s’épanouir comme arbres dominants sur la canopée, parfois après plusieurs siècles ou plus d’un millénaire de lente croissance sous le couvert d’autres arbres.
Les principales dryades en Europe sont : sapin, épicéa dans les milieux froids, acides et ou de montagne, le hêtre, le houx et l'if en plaine[8]. Les dryades se comptent par milliers dans les forêts tropicales où ils dépassent très souvent 500 ou 700 ans.
En Amazonie, à Bornéo, Java et Sumatra, probablement plus d'une centaine d'arbres émergents (ils dépassent et dominent le niveau moyen de la canopée) font partie de ces dryades, mais il a récemment été montré que seuls quelques-uns (une douzaine d'espèce en forêt brésilienne atlantique) résistent à la fragmentation forestière, et uniquement loin des lisières ou au cœur des parcelles les plus grandes[9]. ces dernières espèces peuvent donc être considérées comme bioindicatrices de problèmes de fragmentation écologique des forêts[9].
Le groupe des arbres nomades
Ce sont des arbres opportunistes classés dans le groupe précédent des essences post-pionnières, comme le frêne, l'érable, le merisier, le sorbier, le chêne, le pin et le mélèze en zone tempérée de l'hémisphère nord ; ou parfois des dryades (l'Épicéa en Europe), qui peuvent apparaître directement au stade pionnier à partir de graines ou du recru (plants apparus sous le couvert forestier, au stade précédent), et, si les conditions leur conviennent, persister jusqu’au stade « climacique », parfois en devenant rapidement dominantes.
Exceptions
Quelques essences très adaptées aux conditions extrêmes (froid, sécheresse, climat montagnard, salinité élevée) ont une dynamique qui – en conditions optimales pour elles - les rend dominantes dès ce premier stade, au moins pour la strate forestière supérieure ; ce sont par exemple le hêtre ou sapin en zone tempérée, l’acacia en zone chaude et sèche, L’épinette en zone froide, le palétuvier en zone littorale chaude…
Enjeux
La compréhension de ces mécanismes et de la manière dont ils interfèrent fonctionnellement avec la biodiversité et la ressource en eau est importante pour la conservation de la nature, et pour une gestion forestière plus durable et efficace parce que tenant mieux compte des processus naturels, mais aussi pour mieux anticiper sur les modifications climatiques et leurs effets. Leur bon déroulement implique notamment que les flux écologiques puissent normalement s’exprimer, ce qui implique aussi de mieux comprendre et réparer des problèmes tels que la perte de biodiversité (en particulier perte de diversité génétique), la fragmentation écologique en général et la fragmentation écologique croissante des forêts ou leur insularisation écologique en particulier.
Références
- Vocabulaire forestier. Écologie, gestion et conservation des espaces boisés, Forêt privée française, , p. 495.
- Zackrisson O, Nilsson MC & Wardle DA (1996) Key ecological function of charcoal from wildfire in the boreal forest. Oikos 77, 10-19
- Vedder B, Kampichler C, Bachmann G Bruckner A & Kandeler E (1996) Impact of faunal complexity on micr obial biomass and N turnover in field mesocosms from a spruce forest soil. Biology & Fertility of Soils 22, 22 - 30
- Webb, D.P., 1977. Regulation of deciduous forest litter decomposition by soil arthropod feces. In: Mattson, W.J. (Ed.), T he Role of Arthropods in Forest Ecosystems. Springer, New York, pp. 57 - 69.
- Wolters V (1988) Effects of Mesenchytraeus glandulosus (Ol igochaeta, Enchytraeidae) on decomposition processes. Pedobiologia 32, 387 - 398
- Bernier N, Ponge JF (1994) Humus form dynamics during the sylvogenetic cycle in a mountain spruce forest. Soil Biology & Biochemistry 26 : 183-220
- Staaf H (1987) Foliage turnover and earthworm populations in three beech forests of contrasting soil and vegetation types. Oecologia 72, 58 - 64.
- Annick Schnitzler-Lenoble, « En Europe, la forêt primaire », La Recherche, no 290, , p. 70
- M.A. Oliveira, A.M.M. Santos, M. Tabarelli, “Profoundimpoverishment of the large-treestand in ahyper-fragmentedlandscape of the Atlanticforest” ; Forest Ecology and Management ; Volume 256, Issue 11, 20 November 2008, Pages 1910–1917 (Résumé)
Voir aussi
- Forêt par pays, forêt boréale et forêt amazonienne (les plus grandes forêts vierges au monde)
- Essence forestière
Articles connexes
- Succession végétale
- Espèce pionnière
- écologie du paysage
- Résilience écologique
- Succession écologique
- Perturbation écologique
- Gestion prosilva
- Sylviculture
- régénération
- Gestion durable des forêts
- Forêt modèle
- Forêt primaire
- Paysage forestier intact (PFI)
- Les insectes dans le milieu forestier
- Scolyte
- Agro-sylviculture
- Agrosylvopastoralisme
Bibliographie
- (fr) Courrier de l’environnement de l’INRA ; Forêt (dossier no 15) Prise en compte des cycles sylvigénétiques naturels pour une gestion conservatoire des forêts tempérées. L’exemple des Basses Vosges gréseuses (A. Schnitzler) ; (Une gestion durable des forêts et la conservation de leur diversité biologique - de leur « naturalité » plutôt - passent par deux actions complémentaires : la re-création de réserves intégrales de surface compatible avec le maintien de toutes les espèces inféodées à l'écosystème forestier et la définition de critères de sylviculture aussi proches que possible de modèles sylvigénétiques naturels pour toutes les forêts gérées. Les forêts des Vosges du Nord, artificialisées, ont vu s’éteindre le Grand Tétras et la Gélinotte. Il faut leur appliquer sans tarder une gestion conservatoire, sur la base de l’étude de l’évolution du milieu à la suite de perturbations naturelles. Et en respectant le paysage, en favorisant les essences autochtones, en laissant vieillir les arbres, en maintenant du bois mort et en réprimant le grand gibier herbivore).
- Bornebusch CH (1930) The fauna of forest soil. Det Forstlige Forsøgsvaesen 11, 1-158
- Zackrisson O, Nilsson MC & Wardle DA (1996) Key ecological function of charcoal from wildfire in the boreal forest. Oikos 77, 10-19.
- Sprugel DG (1991) Disturbance, equilibrium, and environmental variability: what is 'natural' vegetation in a changing environment? Biological Conservation 58, 1-18
Liens externes
- Article / Indicateurs de la biodiversité dans les inventaires forestiers (Unasylva, FAO) (fr)
- Mémento / Protection des forêts (2004, D. Vallauri, WWF)(fr)
- Rapport sur la forêt européenne(ONU/UNEP World Conservation Monitoring Centre Cambridge, UK, 2000, en collaboration avec WWF)(en)
- Ressources naturelles * Canada (fr)
- Types de forêts et de gestions en Europe (Rapport European forest types 2006, avec cartes, 8826 KB). (en)
- Revue FAO Ressources génétiques forestières (2003)
- Succession forestière et biodiversité en milieu tempéré, vidéo de 13:13 par Marion Gosselin, 2015,