Succession écologique
La succession écologique est le processus naturel d'évolution et de développement d'un écosystème en une succession de stades : de la recolonisation initiale à un stade théorique dit climacique. Suivant le type de perturbation écologique ayant entrainé la formation d'un néosol, on peut distinguer la succession primaire de la succession secondaire.

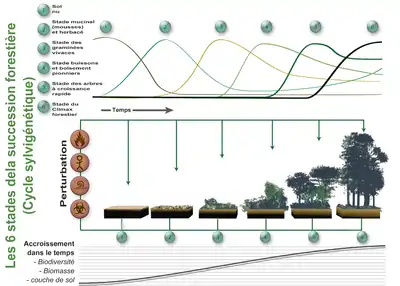
Après un certain temps survient une perturbation qui fait reprendre le "cycle" à son début (ou à un stade intermédiaire si la perturbation est peu importante).
Dans le bas de l'image est représentée l'accroissement de biomasse (sur pied et dans le sol, animale, végétale et fongique..) de biodiversité et d'épaisseur de sol (qui a une importance en termes de puits de carbone). Au fur et à mesure de cette succession, les communautés végétales (et les communautés microbiennes, fongiques et animales qui leur sont associées évoluent) en se remplaçant les unes les autres.
La succession écologique est donc l'ensemble théorique des étapes décrivant — dans le temps et l'espace — un cycle évolutif théorique et complet au sein d'un espace écologique donné. Conséquence évolutive de la compétition, la succession s'apprécie du point de vue de l'écologie du milieu et donc, de manière systémique, en termes d'espèce mais aussi de structure d'occupation de l'espace. Ce cycle correspond aussi à une succession d'habitats et de communautés vivantes (biocénose).
Une sère est un des stades successionnels donnés d'une série de communautés végétales ou animales.
Modèles pour expliquer les successions écologiques
Les botanistes danois Bülow Warming et américain Henry Chandler Cowles développent les modèles de successions végétales dans leur pays respectif. Le biogéographe Henri Gaussen est à l'origine en France de la notion de successions de la végétation, fondatrice de la phytogéographie[1].
Plusieurs modèles sont proposés pour expliquer la succession des espèces dans le processus de colonisation : modèle de la facilitation (création d’un sol ou rétention d'un sol plus épais, protection contre le vent, changement de température, d'ombrage) ; modèle de tolérance (les premiers colonisateurs n'ont ni effet positif ni effet négatif sur l'arrivée des suivants, par exemple des plantes tolérant à l'ombre qui, plus compétitives, éliminent les premiers stades) ; modèle d'inhibition (antériorité de l'espèce compétitive qui empêche le développement d'espèces héliophiles ou plus exigeantes, effet Janzen–Connell)[2]. L'effet Janzen–Connell propose que certaines niches sont délimitées par des rétroactions négatives dans lesquelles des plants adultes, via des interactions intraspécifiques (leur cortège de pathogènes), inhibent le développement de jeunes plants de la même espèce à proximité d'eux. De plus, le développement des pathogènes entraîne un déclin de vigueur des plants adultes qui sont progressivement remplacés par une autre espèce compétitive qui n'a pas les mêmes pathogènes[3] - [4]. « D'une manière générale, les espèces transitoires des successions écologiques seraient frappées de rétrocontrôles négatifs, favorisant ainsi l'arrivée d'autres espèces. À l'opposé, les espèces se développant en fin de succession favoriseraient le développement d'un réseau mycorhizien qui leur serait essentiellement bénéfique[5]. »
Succession autogénique

Ce sont des successions uniquement liées aux interactions entre les organismes sans influence extérieure. C'est un processus biotique.
Succession autogénique primaire
Une succession autogénique primaire est caractérisée par l'établissement de la vie végétale sur un substrat vierge tel qu'une coulée de lave, un sol décapé, des éboulis récents mais aussi un mur en pierre. Les premiers organismes (bactéries, champignons, microflore et microfaune, végétation simple) à s'établir sur un terrain neuf sont alors qualifiées d'espèces pionnières ou de communautés pionnières. Il peut s'agir d'espèces symbiotiques telles que les lichens.
Succession autogénique secondaire
Par opposition, le deuxième type de succession écologique appelé succession autogénique secondaire est caractérisé par l'établissement d'espèces végétales de plus en plus complexes, dans un biotope ayant déjà accueilli la vie mais ayant subi une perturbation écologique telle qu'une inondation qui a affecté l'écosystème mais sans l'éliminer entièrement.
Dans les successions végétales, la construction progressive de l'écosystème débute typiquement par une strate herbacée de plantes annuelles, puis de vivaces. Elles modifient le milieu et le préparent pour accueillir la strate buissonnante (buissons, formation végétale constituée essentiellement des arbrisseaux et sous-arbrisseaux)[6], la strate arbustive (arbustes et jeunes arbres) et enfin la strate arborée, chacune de ces strates intégrant les précédentes et remaniant les contraintes, notamment microclimatiques.
« Les traits caractéristiques des espèces de début de succession sont par exemple un taux de croissance rapide, une hiérarchie faible du système ramifié, une durée de vie courte, une petite taille à l’âge adulte, une production importante de graines de petite taille, pouvant se disperser à des distances considérables grâce au vent ou aux petits animaux, leur conférant des avantages compétitifs dans les milieux ouverts (voir Huston et Smith, 1987 pour revue détaillée[7] ; Diaz et al., 2016[8]). Les plantes des stades plus avancés de la succession ont des comportements caractérisés par des traits tels qu’un taux de croissance lent, une forte hiérarchie du système ramifié, une durée de vie longue, une grande taille (en) à l’âge adulte, une faible production de grosses graines se dispersant sur de courtes distances grâce à la gravité ou à des animaux de grande taille (mammifères), leur conférant un avantage dans les milieux fermés (voir Hutson et Smith, 1987 pour revue détaillée ; Diaz et al., 2016). Ces combinaisons de traits covarient avec les formes de croissance des plantes : plutôt herbacées, annuelles ou pérennes en début de succession, puis progressivement lianescentes, arbustives et arborées en fin de succession (Clements, 1916[9] ; Horn, 1974[10] ; Whittaker, 1975[11] ; Küppers, 1989[12])[13]. »
Succession allogénique
Ce sont des successions qui ne sont pas liées aux relations entre les organismes mais à des facteurs externes (incendie, homme, cataclysme, pollution).
Série régressive
On passe généralement d’un climax à un système simplifié : la simplification est d’autant plus importante que la perturbation est forte.
Succession secondaire
La série peut amener à un dysclimax : à la suite de la perturbation, l’écosystème est dans l’incapacité de récréer le climax d’origine ; il y a alors formation d’un climax moins complexe.
Succession cyclique
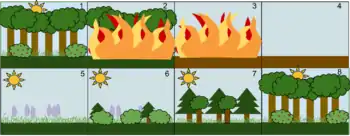
Les successions cycliques sont relativement rares. Dans une telle succession, quelle que soit la perturbation il y a retour au climax et non à un dysclimax. L'exemple typique est les landes bretonnes avec les incendies : l'incendie fait disparaître la lande mésophile arborée, on a un sol nu puis un groupement pionnier muscino-lichenique suivi d'une pelouse ouverte, vient ensuite une pelouse fermée ou lande pionnière puis une lande mésophile moyenne, à laquelle succède une lande mésophile à ajoncs puis enfin un retour à la lande mésophile.
Processus complexe
La succession est caractérisée par une série d'étapes mais aussi en fond par des processus constants de recyclage de la nécromasse par les espèces nécrophages, détritivores et saproxylophages. Certains auteurs estiment que nombre de ces processus sont mis en péril par les activités humaines avec par exemple l'élimination de l'environnement terrestre :
- de la matière organique et de l'humus au profit de sols de plus en plus minéraux, dégradés et instables ;
- des embâcles naturels ;
- des cadavres de la grande faune mammifères (privant la faune nécrophage d'une grande partie de sa nourriture et l'empêchant de redistribuer les sels minéraux et nutriments qu'ils y récupéraient autrefois ;
- de l'équivalent pour le monde végétal des cadavres de grands mammifères ; les gros et très gros bois-morts.
Processus itératif
Si ce processus peut être théoriquement décrit à une échelle locale comme un processus régulier, il est dans la réalité régulièrement interrompu par différents aléas (perturbations anthropiques et/ou naturelles du milieu). Il est donc itératif.
Ceci explique qu'aux échelles paysagères ou supérieures, dans un même milieu (forestier par exemple), divers habitats naturels (ou semi-naturels) et stades écologiques coexistent, généralement dans une structure « en mosaïque » à divers stades d'évolution du cycle (sylvigénétique dans le cas de la forêt), ce qui explique la coexistence de strates écologiques variées, dans laquelle se côtoient des espèces caractéristiques des différents stades de la succession évoluant à différents stades de maturité[14]. Selon l'ensemble de leurs stratégies adaptatives, les essences sont classées en trois catégories : essences pionnières au stade pionnier[15], post-pionnières au stade transitoire[16] et dryades au stade terminal ou optimal[17]. Les nomades ou espèces opportunistes[18] issues des deux derniers types, peuvent s'installer directement en phase pionnière dans certaines conditions (stratégie de tolérance)[19].
Importance et utilité de la notion de « succession »
Le processus est évolutif, et consiste en une série d'étapes devant se succéder chronologiquement dans un ordre fonctionnellement contraint.
Lorsque l'Homme imite les processus de résilience écologique en voulant les hâter (reboisement accéléré ou reforestation par plantation ou régénération naturelle, génie écologique, génie végétal, etc.), l'omission d'une seule étape peut empêcher le bon déroulement des étapes ultérieures. En particulier le stade pionnier a une grande importance pour la restauration ou apparition du sol et de l'ancrage des végétaux, la capacité du milieu à stocker l'eau, etc.
Gestion des espaces naturels
La majorité des habitats naturels est vouée à évoluer naturellement vers le boisement s'ils ne sont pas contraints par un usage (agriculture, dépendances vertes…) et une valorisation humaine. En effet, la succession végétale conduit généralement à une fermeture des milieux, c'est-à-dire la colonisation progressive des milieux ouverts dominés par la strate herbacée par des espèces végétales arbustives puis arborescentes. Des habitats ouverts et espèces associées peuvent être amenés à disparaître au fil des années et des changements de conditions (facteurs abiotiques et biotiques). Beaucoup d'élus ruraux, agriculteurs, aménageurs et gestionnaires de la biodiversité condamnent cette fermeture des milieux. « Dès lors, tous les outils sont bons pour dégager les moyens financiers nécessaire à l'entretien des espaces ouverts protégés contre la dynamique spontanée : programme Life, Fonds européen agricole pour le développement rural (Feader), Fonds européen de développement régional (Feder), contrat Natura 2000, mesures agri-environnementales, aides des collectivités territoriales, notamment au travers de la politique des espaces naturels sensibles ou des réserves naturelles régionales »[20].
Annexes
Articles connexes
- Conséquences évolutives de la compétition
- Résilience écologique
- Cycle sylvigénétique
- Hydrosère
- Biocénose
- Cycle biogéochimique
- Équilibre écologique
- Stabilité écologique
- Écosystème, biotope
- Niche écologique
- Écologie du paysage
- Histoire environnementale
- Écologie rétrospective
- Évolution
- Perturbation écologique
- Incendie de forêt
- Tempête
- Sélection et adaptation durant les successions végétales
- Inondation
- Érosion des sols
- Succession écologique lors de feu de forêt
- Végétation naturelle potentielle
Références
- Claude Tassin, Paysages végétaux du domaine méditerranéen, IRD Éditions, , p. 44
- (en)Joseph H. Connell & Ralph O. Slatyer, « Mechanisms of succession in natural communities and their role in community stability and organization », The American Naturalist, vol. 111, n°982, 1977, p.1119-1144.
- (en) Brian D. Fath, Encyclopedia of Ecology, Elsevier, , p. 597-598.
- Marc-André Selosse, Jamais seul. Ces microbes qui construisent les plantes, les animaux et les civilisations, Actes Sud Nature, , p. 87.
- Marc-André Selosse, Franck Richard, Pierre-Emmanuel Courty, « Plantes et champignons : l'alliance vitale », La Recherche, no 411, , p. 60.
- En sous-bois, les fougères, les ronces, le houx, les genêts, le cornouiller, le genévrier, l'alisier font du taillis un fourré. Le prunellier, l'aubépine et l'ajonc forment des fourrés dans les clairières. Toutes ces espèces pionnières composent des haies pionnières, « berceaux des arbres ». Cf Cogneaux Christian et Gambier Bernard, Plantes des haies champêtres, Rouergue, , p. 47
- (en) Michael Huston & Thomas Smith, « Plant Succession: Life History and Competition », The American Naturalist, vol. 30, no 2, , p. 168-198 (DOI 10.1086/284704)
- (en) S. Diaz et al., « The global spectrum of plant form and function », Nature, vol. 529, no 7585, , p. 167-171 (DOI 10.1038/nature16489)
- (en) F.E. Clements, Plant indicators: the relation of plant communities to process and practice, vol. 290. Carnegie Institution of Washington. 1916
- (en) Henry S. Horn, « The ecology of secondary succession. », Annu. Rev. Ecol. Syst., vol. 5, , p. 25-37 (DOI 10.1146/annurev.es.05.110174.000325).
- (en) Robert H. Whittaker, Communities and Ecosystems. Macmillan, Macmillan, , 385 p..
- (en) M. Küppers, « Ecological significance of above-ground architectural patterns in woody plants: A question of cost-benefit relationships », Trends in Ecology & Evolution, vol. 4, no 12, , p. 375-379 (DOI 10.1016/0169-5347(89)90103-1).
- Mathieu Millan. Analyse de la variabilité des traits architecturaux des formes de croissance dans les communautés végétales. Botanique. Université de Montpellier, 2016, p. 64
- Eugénie Cateau, Laurent Larrieu, Daniel Vallauri, Jean-Marie Savoie, Julien Touroult, Hervé Brustel, « Ancienneté et maturité : deux qualités complémentaires d’un écosystème forestier », Comptes Rendus Biologies, vol. 338, no 1, , p. 58-73 (DOI 10.1016/j.crvi.2014.10.004).
- Essences de pleine lumière, anémochores (à fort pouvoir de dispersion), à forte croissance juvénile mais à faible longévité, à croissance juvénile rapide mais à faible longévité, colonisant les milieux ouverts, perturbés ou instables (exemples : bouleaux, saules, aulnes, peuplier tremble).
- Essences de plus grande taille, plus longévives et à croissance plus lente, s’installant en général après les pionnières, et restant des essences de lumière dans le jeune âge (exemples : chênes, charmes, ormes, tilleuls, érables, sorbiers, pins, mélèze, frêne commun).
- Essences d'ombre arrivant en fin de cycle, de grande longévité et à croissance lente (hêtres, épicéas, sapins, if).
- Espèces se développant lorsqu'un blocage stationnel local arrête la succession. C'est le cas, par exemple, des forêts sur éboulis instables. Exemples : Pins, Épicéa commun, Chêne pédonculé, Chêne tauzin, Chêne pubescent, ormes, érables.
- Gérard Dumé, Jean-Claude Rameau, Christian Gauberville, Dominique Mansion, Flore forestière française, Institut pour le développement forestier, , p. 22.
- Annik Schnitzler et Jean-Claude Génot, La France des friches. De la ruralité à la féralité, éditions Quæ, (lire en ligne), p. 142.