Conséquences évolutives de la compétition
La compétition est une forme d’interaction négative entre individus de la même espèce (compétition intraspécifique) ou d’espèces différentes (compétition interspécifique) qui entrent en concurrence pour l’exploitation directe de ressources présentes dans leur environnement ou qui par leurs présences réduisent la capacité des autres individus à disposer de ces ressources.
Ces interactions affectent négativement la fitness des individus entrant en compétition. La compétition entraîne alors l’augmentation des pressions de sélection sur les individus les forçant à évoluer pour réduire ces mêmes pressions. Ces évolutions étant définies par des changements dans le temps des caractères morphologiques et/ou génétiques des individus appartenant aux populations en compétitions.
Un individu en compétition est défini par son habilité compétitive : Elle est composée de deux éléments, l’effet compétitif et la réponse compétitive.
L’effet compétitif est la capacité d’un organisme à réduire le fitness des organismes avec lesquels il intéragit compétitivement. La réponse compétitive est l'habilité à supporter les effets négatifs des voisins[1].
Les mécanismes par lesquels se fait la compétition
Interférence
On observe une interaction directe entre les individus ou populations en concurrence pour l'obtention d'une ressource. Cette interférence peut être active, par exemple sous forme de combats, vols (kleptoparasitisme), ou passive par exemple l'utilisation des métabolites secondaires chez les plantes.
Exploitation
Ce type de compétition se fait sans interaction directe entre les concurrents. Elle vient de l’utilisation de ressources communes par un des concurrents, ce qui diminue la disponibilité des ressources pour l’autre.
Évolution des traits dans une population
La compétition à travers ses différentes formes va contraindre et faire évoluer les traits d’histoire de vie, sélectionnant les individus ayant une meilleure fitness. Les traits comportementaux, physiologiques, anatomiques et morphologiques vont donc évoluer dans la population en fonction de l’environnement et de la compétitivité des autres individus. L'héritabilité des caractères est un point clé dans la sélection naturelle, et cette variation de trait au fur et à mesure des générations est étudié grâce à la génétique quantitative.
Équation du changement du trait moyen de la population de Hadfield[2]:

Avec un trait ressource dépendant dans la population caractérisé par de la variance génétique pour l'acquisition de la ressource sous conditions compétitives,

la compétitivité, le gradient de sélection, A le total de ressource pour N individu en compétition, la variance de l’additivité génétique du trait 2 et la covariance additive positive des traits 1 et 2.




A et N impliquent que si la quantité de ressource augmente ou le nombre de compétiteur diminue alors la moyenne du trait 1 va augmenter.
décrit le changement évolutif du trait 1 attendu comme une conséquence de la sélection direct du trait corrélé génétiquement avec l'habilité compétitive.

Si on part du postulat que seule l‘acquisition de la ressource a un gradient de sélection non nul, alors on peut remplacer ce terme par la covariance de l’additivité génétique entre le trait 1 et la fitness relative, et peut être interprété comme l’identité de Robertson Price :

correspond à la diminution de la variation de causée par la détérioration de l’environnement, car la variance génétique pour l'habilité compétitive implique des effets génétiques indirects sur l’acquisition de la ressource.

Interspécifique
Il y a compétition interspécifique lorsque les organismes de deux espèces différentes entrent en concurrence pour l'obtention d’une même ressource.
Exclusion compétitive
Selon le principe d'exclusion compétitive de Gause (1934): Dans un environnement stable, lorsque deux espèces sont en compétition pour une ressource limitée, celle qui utilise ses ressources le plus efficacement finira par éliminer localement l’autre. Deux espèces peuvent coexister sur un long terme si les ressources ne sont pas limitantes ou que leurs niches écologique différèrent toujours d’une manière ou d’une autre.
Darwin émit le postulat comme quoi la lutte pour l’existence est plus importante entre espèces plus fortement apparentées[3].
Les espèces proches sont plus susceptibles d’avoir des niches similaires et ainsi de subir une plus forte compétition. L’exclusion compétitive d’espèces proches est sans doute la cause de la coexistence fréquente d’espèces distantes dans les communautés écologiques. C'est la sur-dispersion phylogénétique: les espèces cohabitant au sein d’une communauté locale sont en moyenne moins apparentées que des espèces de communautés distinctes[3]. La sur-dispersion phylogénétique est souvent attribuée à l’exclusion compétitive d’espèces proches d’un point de vue phylogénétique (Phylogénétique limiting similarity Hipothesis) même si d’autres facteurs sont sans doute impliqués, surtout dans les communautés naturelles[3].
Ce cas fut observé chez certaines espèces de Tamia[4], dans des montagnes du Nevada deux populations de Tamia peuvent coexister grâce à des comportements et des aptitudes différentes, bien qu'elles exploitent les mêmes ressources que sont les fruits et graines de genévrier et de pin. La première espèce est Tamias dorsalis qui possède un caractère extrêmement agressif face aux possibles compétiteurs qu'ils soient de la même espèce ou d'une espèce proche, lors d'une confrontation il va poursuivre l'adversaire jusqu'à la fuite de ce dernier. La seconde espèce est Tamias umbrinus qui est une espèce fortement arboricole et plus sociale comparée à Tamias dorsalis, cette espèce vit la plupart du temps dans les arbres, s'il y a danger elle s'y réfugie pour être hors de portée et reste immobile en attendant que le danger passe. Lors de leurs rencontres, Tamias dorsalis agresse Tamias umbrinus, si la zone où l'agression a lieu est dégagée la tactique d'agression de Tamias dorsalis sera avantagée, si l'agression a lieu dans une forêt dense la fuite de Tamias umbrinus sera facilitée et Tamias dorsalis dépensera de l'énergie inutilement pour pourchasser son compétiteur.
Il y a coexistence d’espèces en compétition seulement si elles occupent des niches écologiques différentes. On peut alors observer une différenciation de niche.
Séparation écologique
Deux populations présentes dans un habitat donné peuvent coexister s'il y a un minimum de différences entre leurs niches écologiques. On observe une séparation écologique des espèces concurrentes due à la sélection naturelle qui favorise les individus divergents, à partir du moment où leur comportement s’expriment dans leurs gènes. Dans ce cas, la compétition interspécifique a pour effet le rétrécissement de la niche écologique, ce qui favorise le processus de glissement des niches associé dans le long terme d'un phénomène de déplacement des caractères dû à une évolution des caractères phénotypiques propres à chacune des espèces en présence (spécialisation des espèces)[5].
Sélection directionnelle
Dans un environnement où on observe la présence d’une espèce présentant un polymorphisme qui entraine l’utilisation, par des individus d’une même population, de ressources différentes; si une espèce immigrante arrivant dans l’environnement est une meilleure compétitrice que l’espèce résidente pour une ressource commune; alors l’espèce immigrante va exclure l’espèce résidente d’une partie des ressources exploitables. La population résidente va alors réduire son polymorphisme afin de réduire les pressions entrainées par la compétition, on aura alors disparition ou réduction de certains phénotypes utilisé dans l’acquisition des ressources communes[6].
Ce cas fut observé dans certaines Îles des Galápagos avec des Pinsons de Darwin[6]. Geospiza fortis utilisait deux ressources différentes, des graines de petites et de grandes tailles. Seuls les individus avec de gros becs pouvaient consommer les grosses graines, cela leur donnait un avantage en période de sécheresse où les petites graines se faisaient rares. Lors de l’arrivée de Geospiza magnirostris dont la ressource principale était ces grosses graines, ces derniers sont devenus les compétiteurs dominants pour cette ressource. Chez Geospiza fortis, après plusieurs décennies on a observé de moins en moins d’individus à gros bec, le taux d’individus à petit bec a augmenté jusqu'à quasi-disparition du trait gros bec dans la population à cause des pressions de sélections.
Successions végétales
La compétition entre populations différentes de plantes conduit à l’instauration d’une dynamique de successions végétales au cours du temps[1].
Chez ces plantes soumises à la compétition, leur réponse compétitive se décline en deux processus : une conservation des ressources en stockage et sûreté, la réponse persistante. Ou, une tolérance des voisins en acquérant des ressources plus rapidement qu’eux ou dans des lieux/moments différents c’est le foraging ou réponse d’évasion. On observe souvent un trade-off (compromis évolutif) entre ces deux processus : priorité pour la croissance rapide ou sur la conservation des ressources[1].
Le long de la succession végétale, on observe une limitation croissante de la luminosité. Cela se traduit par une augmentation de la compétition entre les plantes pour l’accès à la lumière: on peut observer un accroissement de la biomasse et de la quantité de litière au cours du temps, ce qui joue sur l’augmentation de l’humidité du sol[1].
Le trait principal de ces plantes qui est affecté par la compétition est la biomasse totale des plantes, que l’on peut seconder par le diamètre des tiges dans les successions végétales d’herbacées. En effet, elles ont tendance à s’étendre plutôt que de croître en hauteur. Chez les populations de plantes vivant dans des environnements à luminosité réduite on peut de la même manière considérer la taille des plantes[1].
Ces adaptations ont permis de sélectionner les meilleures compétitrices le long des successions et du temps. Il semblerait en effet, selon certaines études, que les espèces des successions avancées soient plus compétitives, et que le milieu des successions primaires provoque plus de compétition. On a eu une sélection dynamique des espèces au cours du temps, de l’espace et au cours de la succession[1].
Intraspécifique
Il y a compétition intraspécifique lorsque des organismes de la même espèce entrent en concurrence pour l'obtention d’une même ressource.
Densité dépendance négative
C'est le mathématicien Pierre François Verhulst qui prit en compte la densité dépendance négative dans son modèle de croissance logistique en 1836. Dans une population la densité peut augmenter jusqu’au point où la compétition intra spécifique réduira la survie et/ou la fécondité individuelle. Cette densité de population maximum possible dans le milieu est appelée la capacité de charge du milieu et on estime que la population atteint un équilibre lorsque chaque individu a assez de ressources pour produire un descendant viable[7].
La compétition intraspécifique est donc densité dépendante négative lorsqu'il y a une relation négative entre la densité de population et son taux de croissance. En effet, une forte densité d’individus aura un impact négatif sur la reproduction et la survie des individus et en conséquence la population croîtra moins rapidement. Dans ce cas, la surface de la courbe de fitness est réduite de façon homogène en proportion avec la densité de population totale. À haute densité la fitness est alors plus faible mais tous les phénotypes étant affectés de la même manière sa pente ne change pas[7].
Fréquence dépendante
La compétition peut aussi être fréquence dépendante. C’est le cas lorsque les différents phénotypes dans une population utilisent des parties différentes des ressources disponibles, ceci étant due à des compromis/trade-off dans l’utilisation de ressources alternatives[7]. La compétition est alors une fonction de la densité des phénotypes identiques plutôt que de la taille de la population. Cependant le fait d’augmenter ou diminuer la taille de la population changera l’intensité de cette compétition. La compétition densité et fréquence dépendante affecte négativement la fitness du phénotype le plus abondant même s’il est adapté à la ressource la plus abondante. Les phénotypes consommateurs rares ont certes moins de ressources disponibles mais aussi moins de compétiteurs avec lesquels les partager; il en résulte que leur fitness totale est relativement élevée[7].
Conséquences évolutives
Quand la compétition interspécifique diminue, la compétition intraspécifique devient une force évolutive potentielle favorisant la variation écologique et génétique.
Extension de niche
La compétition densité et fréquence dépendante peut mener a un agrandissement de la niche écologique d’une espèce et a la diversification des phénotypes associés. Lorsque la population n’utilise qu’une partie des ressources disponibles dans le milieu, cela favorise la diversification en diminuant la valeur sélective des individus qui utilisent la ressource ancestrale jusqu’au point où les ressources qui étaient sous optimales avant permettent d’acquérir une fitness plus importante. Les individus se voient alors obligés d’évoluer vers une utilisation plus large des ressources de la niche afin de diminuer la pression de sélection exercée par la compétition. Le taux de compétition est le facteur qui favorise et accélère cette évolution. En effet, les populations subissant un fort taux de compétition s’adaptent plus rapidement a la nouvelle ressource que celles qui subissent un taux plus faible[8].
On a observé ce phénomène d’extension de niche chez la mouche Drosophila melanogaster qui est intolérante au Cadmium[8]. En effet lorsque ces mouches sont placées dans des cages contenant des fioles avec différentes concentrations de Cadmium, dans des conditions de compétition fréquence dépendante forte et de ressource limitante, elles évoluent rapidement vers des performances accrues sur les milieux avec du Cadmium afin d’utiliser cette nouvelle ressource plus efficacement. Cette résistance élevée au Cadmium en condition de forte compétition étant héritable, le comportement de ponte des mouches adaptées au Cadmium sur un milieu en contenant facilite l’extension de la niche[8].
L’extension de la niche écologique peut être associée à une augmentation de la variance phénotypique, à l’évolution du polymorphisme ou à la divergence en plusieurs espèces utilisant des ressources différentes[8].
L'extension de niche favorise les espèces euryèces (capables de vivre dans plusieurs niches écologiques). Sur le plan évolutif, ces généralistes varient peu au cours des temps géologiques, elles sont dans une certaine mesure à l'abri d'une pression sélective novatrice, ce qui les oppose aux espèces sténoèces (étroitement dépendantes du milieu dans lequel elles vivent). L'extension caractérise aussi l'homme : confrontée à l'explosion démographique et l'augmentation des besoins alimentaires, l'humanité s'est mise à cultiver des terres de plus en plus marginales, accentuant l'érosion des sols et la désertification[9].
Sélection disruptive/diversifiante
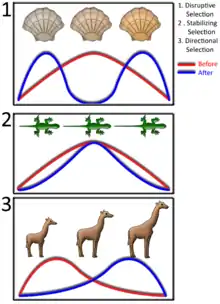
Dans une population les individus ayant un phénoype intermédiaire sont défavorisés par la sélection naturelle par rapport aux individus ayant des phénotypes extrêmes. La compétition intraspécifique lorsqu’elle est fréquence dépendante, et donc quand le phénotype le plus abondant est au minimum de la fitness de la population, peut mener à de la sélection disruptive[7]. Mais cela dépendra de l’importance sélective de cette compétition et du degré de spécialisation individuel nécessaire pour générer de la fréquence dépendance. L'accouplement au hasard, la plasticité phénotypique, les effets d’autres forces sélectives comme la prédation ou la sélection sexuelle, ou la variation du niveau de compétition peuvent atténuer les effets à long terme de la sélection disruptive. La spéciation et le dimorphisme sexuel sont des réponses possibles à la sélection disruptive. Lequel des deux apparaîtra dépend de la covariance génétique entre les traits des mâles et femelles et du potentiel génétique pour l’accouplement d’individus ayant des traits en commun. La sélection disruptive peut aussi aboutir à des polymorphismes ou à une plus grande variation phénotypique[7].
Spéciation sympatrique
La compétition pour les ressources limitées peut restreindre l’utilisation de l’habitat et l’aire géographique de répartition des espèces, et cette compétition peut mener à des déplacements de caractères phénotypiques où des lignées très proches génétiquement ne peuvent plus se reproduire. L’isolement reproductif dans une aire géographique continue et l’acquisition de différences morphologiques s'appelle la spéciation sympatrique[12].
Quand la compétition est la pression de sélection la plus importante et que la capacité de dispersion des individus est limitée, les deux niches écologiques différentes de l’espèce ne seront pas séparés géographiquement.
Les individus les moins compétitifs étant plus limités dans l’acquisition des ressources seront contraints de changer d’habitat dans certains cas. Des évolutions de traits vont apparaître au fur et à mesure des générations afin de développer des adaptations à leur nouvel habitat.
Il va donc y avoir une restriction des flux de gènes et des changements phénotypiques majeurs qui vont entraîner un isolement reproductif entre deux populations d’une même espèce.
Exemple d’un Cichlidae[12] : Telmatochromis temporalis. Les individus de cette espèce de poisson sont en compétition par interaction direct pour leur habitat dans le lac Tanganyika en Afrique de l’Est. Le mécanisme de compétition apparent est la compétition par contest, où les moins compétitifs finissent par être exclus de l’habitat préférentiel. Les pressions de sélection au fur et à mesure des générations vont conduire à l’apparition de deux écomorphes (population de même espèce mais ayant un phénotype lié à leur environnement), avec une population vivant sur des substrats rocheux ayant une taille plus grande, et une population vivant sur un substrat de coquilles, avec des adaptations de traits d’histoire de vie permettant de maximiser leur fitness (par exemple leur taille, leur âge à la maturité et leur nombre de descendant)[12].
Polymorphisme de ressource
Le polymorphisme de ressource est un changement de traits phénotypiques en fonction de l’utilisation de l’habitat sans isolement reproductif.
La compétition intraspécifique pour une ressource limitante entre les individus au sein d’une même espèces peut entraîner un changement de niches. Les individus les moins compétitifs étant réduits à un habitat de moindre préférence. On constate en fonction des différentes niches occupées par l'espèce que des variations morphologiques apparaissent, montrant des adaptations différentes en fonction de la ressource afin de compenser la fitness.
Les exemples de ce polymorphisme de ressource sont bien connu dans certaines espèces comme les gerbilles, les colibris et certaines espèces de poisson[13].
La perche Perca fluviatilis et le gardon Rutilus rutilus vivent soit dans la zone littorale des lacs soit dans la zone pélagique en fonction de la compétitivité des individus. Ces deux habitats sont différents du point de vue des ressources : zooplanctons en zone pélagique, puis des poissons et macro-invertbrés en zone littorale. On constate qu’en fonction de l’habitat, un polymorphisme de ressource est visible, les perches attrapées au niveau du littoral ayant un corps plus épais et une bouche plus large que celle de la zone pélagique et idem pour les gardons.
Modifications du sexe ratio
Le sex-ratio est le rapport des sexes (soit la proportion de mâles, soit la proportion de femelles) dans une population donnée.
Il peut être étudié à différents stades de développements des individus. On distingue le sexe ratio primaire à la conception, le sexe ratio secondaire à la naissance et le sexe ratio tertiaire à maturité. Ceci, selon le taux de mortalité aux différents stades et sa discrimination selon les sexes dans la population.
Malgré l’anisogamie des gamètes de la reproduction sexuée, on observe souvent des sexes ratio équilibrés dans les populations.
Théorie du 50/50, Düsing et Fisher[14]: On pose comme hypothèses:
- Qu’il n’y a pas de différence dans le coût des descendants selon leur sexe.
- Que le gain de fitness des descendants ne dépend pas de leur sexe.
- Qu’il y a toujours un équilibre quand la production de mâles et femelles n’apporte pas d'avantage.
On observe alors une répartition du sexe ratio sous forme fréquence dépendance.
Or ces hypothèses ne sont pas toujours vérifiées dans le monde vivant.
Si le coût ou le bénéfice est supérieur pour l’un des deux sexes on observe une manipulation adaptative du sexe ratio. Par exemple, chez le cerf élaphe[14] ou Red deer on observe une augmentation du sexe ratio en faveur des mâles avec le statut dominant des mères. On peut également observer un investissement préférentiel des mères pour le sexe qui a un meilleur taux de reproduction et qui est souvent le sexe mâle.
La compétition a également des impacts sur le déterminisme du sexe. La LRC : local resource competition (compétition locale pour les ressources) est impliquée dans la variation du sexe ratio dans plusieurs études chez les mammifères[14].
L’élevage des mâles est souvent plus onéreux pour les femelles que celui de leur fille à cause du dimorphisme sexuel. Ils demandent plus d’investissement et de ressources, comme le coût de la gestation et de la lactation chez le cerf élaphe[14].
Également, lorsque les mères sont en mauvaise condition physique par manque de ressources, elles auront tendance à privilégier le sexe dispersant pour réduire l’exploitation future des ressources du milieu. Des femelles en bonne condition, vivant dans un milieu riche, privilégierons le sexe non dispersant et seront capables de pourvoir correctement à son développement. Ce dernier point a été démontré dans une étude sur les chevreuils, particulièrement adaptés pour cette démonstration. Le sexe dispersant étant chez ses animaux le sexe mâle, quand le milieu se dégrade et que les ressources disponibles se font rares, les mères ont tendance à privilégier le sexe mâle sur le sexe femelle[14].
Pour éviter la LRC, on privilégie le sexe non philopatric (qui reste sur place).
Compétition spermatique
La sélection sexuelle résulte des différences de succès reproducteurs liées à la compétition pour l’obtention d’un partenaire. La sélection intrasexuelle se fait entre mâles qui sont en compétition pour l’accès aux femelles. En effet en raison de l'anisogamie, les femelles représentent une ressource rare. Une des composantes de cette sélection sexuelle est la compétition spermatique qui a lieu après l’accouplement.
La compétition spermatique est la compétition entre les spermatozoïdes de plusieurs mâles pour la fécondation des ovules d’une femelle. Elle intervient donc pour les espèces chez lesquelles les femelles sont polyandres. Cette compétition est une force évolutive qui a permis la mise en place de traits et comportements apportant un avantage évolutif du point de vue de la reproduction.
Chez les mammifères, les mâles des espèces où les femelles sont polyandres ont un nombre de traits reproductifs qui sont le résultat des pressions de sélection relatives à la compétition spermatique[15]. Leurs testicules sont plus larges en relation avec la taille de leur corps et elles ont plus de tubes séminifères, ils produisent plus de spermatozoïdes dont les flagelles sont plus longs que chez les espèces monogames et une plus grande partie de ceux-ci est mobile. Sachant qu’il y a une corrélation positive entre la taille du spermatozoïde et sa vitesse chez les mammifères, les spermatozoïdes des espèces polyandres atteignent donc plus vite l’ovule et ont plus de chance de le fertiliser[15]. Les mâles faisant face à la compétition spermatique peuvent augmenter leur production de spermatozoïdes en augmentant le nombre de spermatozoïdes par éjaculât ou en augmentant le nombre d’éjaculâts, d'accouplements. De plus, il y a une corrélation positive entre le nombre d’accouplements et la taille des spermatozoïdes qui augmente[15].
Références
- (en) Kazakou, E., Navas, M.L., « Variation in intensity of competition along a Mediterranean successional gradient », Proceedings of the 10th MEDECOS Conference,
- (en) Wilson AJ, « Competition as a source of constraint on life history evolution in natural populations », Heredity, (DOI 10.1038/hdy.2013.7)
- (en) Violle C. et al, « Phylogenetic limiting similarity and competitive exclusion », Ecology Letters, no 14, , p. 782-787
- (en) James H. Brown, « Mechanisms of Competitive Exclusion Between Two Species of Chipmunks », Ecology, no 52, , p. 305-311
- François Ramade, Éléments d'écologie, Dunod, , p. 333.
- (en) Peter R. Grant et B. Rosemary Grant, « Evolution of Character Displacement in Darwin’s Finches », Science, no 313, , p. 224-226.
- (en) Bolnick, D.I, « Can intraspecific competition drive disruptive selection? An experimental test in natural populations of sticklebacks; », Evolution, no 58, , p. 608-618.
- (en) Bolnick, D. I, « Intraspecific competition favours niche width expansion in Drosophila melanogaster. », Nature, no 410, , p. 463–466.
- François Ramade, Éléments d'écologie, Dunod, , p. 332.
- Sélection disruptive : les coquilles blanches sur des rochers recouverts de anatifes ou de pouces-pieds, également blancs, ou sur des fonds sableux blancs, sont moins victimes de prédateurs qui ont du mal à les distinguer. Sur les roches nues et de couleur sombre ou sur les fonds marins foncés, les coquilles sombres ont le même avantage. Par contre, les prédateurs repèrent facilement les coquillages à coloration intermédiaire qui se distingue facilement sur des fonds clairs ou sombres. Sélection stabilisatrice : les lézards plus grands que la moyenne sont plus facilement repérés par les prédateurs que les lézards de taille moyenne ; les lézards plus petits que la moyenne fuient moins vites et sont plus facilement capturés par les prédateurs que les lézards de taille moyenne. Sélection directionnelle : chez les girafes, cette sélection naturelle provoque un déplacement de la population dans la direction du phénotype privilégié, celui du cou long.
- Ou le nombre d'individus, corrélé à cette fitness.
- (en) Winkelmann, K et al., « Competition-driven speciation in cichlid fish », Nature, (DOI 10.1038/ncomms4412)
- (en) Svanbäck, R, Eklöv, P, Fransson, R et Holmgren, K, « Intraspecific competition drives multiple species resource polymorphism in fish communities », Oikos, no 117, , p. 114-124
- (en) Hewison AJM, Gaillard JM, « Birth-sex ratios and local resource competition in roe deer, Capreolus capreolus », Behav Ecol, no 7, , p. 461–464
- (en) Gomendio, M et ERS Roldan, « Sperm competition influences sperm size in mammals. », Proceedings of the Royal Society of London B, no 243, , p. 181-185.