Cire épicuticulaire
La cuticule des plantes et des insectes est couverte par une cire épicuticulaire, constituée de composés appartenant à un vaste groupe de substances lipophiles, qui diminue la mouillabilité de la surface et réduit la perte d'humidité. La forme de ces sécrétions cireuses qui donnent à la surface de la cuticule un aspect luisant est un important caractère systématique ; des excroissances cireuses plus massives, appelées pruine, donnent à la cuticule un aspect givré ou poussiéreux.


Chez les végétaux
Composition et analyse
La cire épicuticulaire est constituée principalement d'hydrocarbures aliphatiques à chaîne droite avec une variété de groupes substitués. C'est ainsi qu'on peut observer[3] - [4] :
- des paraffines sur les feuilles de choux et de pois,
- des esters d'alkyle dans les feuilles du palmier à cire, Copernicia prunifera, (cire de carnauba) et de bananier,
- un alcool secondaire asymétrique (10-nonacosanol) dans la plupart des conifères, tels le Ginkgo biloba et l'épinette de Sitka, beaucoup de Ranunculaceae, Papaveraceae et Rosaceae et quelques bryophytes,
- des alcools secondaires symétriques chez des Brassicaceae, notamment Arabidopsis thaliana,
- des alcools primaires (surtout octacosan-1-ol) chez la plupart des graminées , Eucalyptus et des légumineuses,
- des β-dicétones chez de nombreuses graminées, Eucalyptus, Buxus et des Ericaceae,
- des aldéhydes dans des feuilles de hêtre, de jeunes chaumes de canne à sucre et sur des fruits comme le citron,
- des triterpènes dans les cires des fruits des pommiers, pruniers et sur le raisin.
Des composés cycliques, comme les phytostérols, les triterpénoïdes pentacycliques et des flavonoïdes peuvent aussi être présents[5].
Ces composés sont pour la plupart solubles dans des solvants organiques comme le chloroforme et l'hexane, ce qui permet leur analyse chimique, mais chez certaines espèces l'estérification d'acides et d'alcools en étholides ou la polymérisation d'aldéhydes peuvent donner naissance à des composés insolubles. Les extraits obtenus par des solvants contiennent des cires épicuticulaires mélangées à des cires cuticulaires, souvent contaminées par des lipides des membranes des cellules sous-jacentes. La cire épicuticulaire a pu être isolée par des méthodes mécaniques[6] qui ont permis de la distinguer de la cire cuticulaire constituant le polymère cuticule et de démontrer qu'elles sont chimiquement distinctes et organisées en deux couches[7].
Ultrastructure
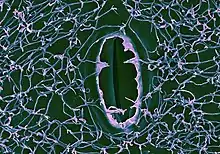
Les cires épicuticulaires se présentent généralement sous la forme de cristalloïdes microscopiques en écailles ou en tubules dont l'orientation et l'ultrastructure sont un important caractère systématique[5]. La forme des cristaux dépend des composés présents : les alcools secondaires asymétriques et les ß-dicétones forment des nanotubes de cire creux, alors que les alcools primaires et secondaires symétriques forment des plaques planes[8].
Ces cires sont responsables des reflets bleutés ou glauques des nombreux épidermes (effet de diffraction sur leur ponctuation cuticulaire) et constituent la pruine des fruits[9].
Rôles
En augmentant la réflexion et la diffusion, les projections cristallines des dépôts cireux protègent la plante du rayonnement incident et réfléchissent les rayonnements UV ; grâce à leur faible mouillabilité, elles augmentent l'imperméabilité et créent une propriété d'auto-nettoyage connue comme l'effet lotus[10] ; enfin elles accroissent la résistance aux agents pathogènes en diminuant le risque de contamination par des poussières, des spores et autres microrganismes[5].
Les cires épicuticulaires étant très hydrophobes, leur destruction augmente l'absorption foliaire[11].
Chez les insectes
Chez les insectes, la cire épicuticulaire est sécrétée principalement par les œnocytes situés sous la cuticule, et connait de grandes variations en composition et en épaisseur au sein de ce groupe. Beaucoup d'autres composants se retrouvent mélangés à cette cire, et reflètent de nombreux rôles secondaires qui sont venus s'ajouter au rôle initial au fil de l'évolution des insectes[12]. Les composés épicuticulaires sont ainsi impliqués dans la communication et dans la défense contre les pathogènes[13].
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Epicuticular wax » (voir la liste des auteurs).
- (en) Wilhelm Barthlott, Matthias Mail, Bharat Bhushan, Kerstin Koch, « Plant Surfaces: Structures and Functions for Biomimetic Innovations », Nanomicro Lett., vol. 9, no 2, , p. 23 (DOI 10.1007/s40820-016-0125-1).
- (en) George Karabourniotis, Georgios Liakopoulos,Panagiota Bresta, Dimosthenis Nikolopoulos, « The Optical Properties of Leaf Structural Elements and Their Contribution to Photosynthetic Performance and Photoprotection », Plants, vol. 10, no 7, , p. 1455 (DOI 10.3390/plants10071455).
- (en) E.A. Baker, « Chemistry and morphology of plant epicuticular waxes », dans D.J. Cutler, K.L. Alvin & C.E. Price, ed., The Plant Cuticle, London, Academic Press, , p. 139-165
- (en) P.J. Holloway et C.E. Jeffree, Epicuticular waxes, coll. « Encyclopedia of Applied Plant Sciences » (no 3), , p. 1190-1204
- Wilhelm Nultsch, Botanique générale, Paris, De Boeck Université, , xvi, 602 (ISBN 2-7445-0022-4)
- (en) H.J. Ensikat, C. Neinhuis et W. Barthlott, « Direct access to plant epicuticular wax crystals by a new mechanical isolation method », International Journal of Plant Sciences, vol. 161, , p. 143-148
- (en) R. Jetter, S. Schäffer et M. Riederer , 23,, « Leaf cuticular waxes are arranged in chemically and mechanically distinct layers: evidence from Prunus laurocerasus L. », Plant, Cell and Environment, vol. 23, , p. 619-628
- C.E. Jeffree, E.A. Baker et P.J. Holloway, « Ultrastructure and recrystallisation of plant epicuticular waxes », New Phytologist, vol. 75, , p. 539-549
- D. Robert, Jean Claude Roland, Biologie végétale : caractéristiques et stratégie évolutive des plantes, Doin, , p. 91
- W. Barthlott et C. Neinhuis, « Purity of the sacred lotus, or escape from contamination in biological surfaces », Planta, vol. 202, , p. 1-8
- Swietlik D. and M. Faust. 1984. Chapter 8: Foliar Nutrition of fruit Crops. Horticultural Reviews 6: 287-353.
- Beutel, Rolf,, Insect morphology and phylogeny : a textbook for students of entomology, , 531 p. (ISBN 978-3-11-026404-3 et 3-11-026404-8, OCLC 900020732, lire en ligne)
- (en) Anna Katarzyna Wrońska, Mieczysława Irena Boguś, Emilia Włóka et Michalina Kazek, « Cuticular fatty acids of Galleria mellonella (Lepidoptera) inhibit fungal enzymatic activities of pathogenic Conidiobolus coronatus », PLOS ONE, vol. 13, no 3, , e0192715 (ISSN 1932-6203, PMID 29518079, PMCID PMC5843172, DOI 10.1371/journal.pone.0192715, lire en ligne, consulté le )