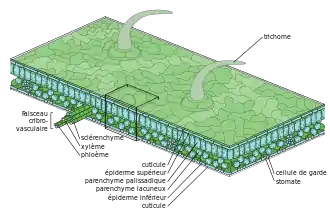
Cuticule (botanique)
La cuticule (du latin cuticula « petite peau ») est la couche externe continue cireuse produite par l’épiderme des organes aériens des plantes terrestres, essentiellement les feuilles et les tiges des plantes vasculaires et de quelques bryophytes. Ce revêtement de la paroi externe des cellules épidermiques n'est pas une couche cellulaire mais une matrice extracellulaire continue constituant un film hydrophobe translucide, formé d'une couche extérieure constituée de cires recouvrant une couche de cutine hydrophobe qui forme la base de la cuticule. Pellicule plus ou moins épaisse (de 0,02 à 20 μm)[5], la cuticule végétale est principalement considérée comme une barrière solide qui a comme fonction principale de protéger les organes aériens de la dessiccation et des rayons ultraviolets (adaptation anatomique à la colonisation de la terre ferme), et a développé de nombreuses fonctions secondaires résistance face à divers stress biotiques et abiotiques).

.jpg.webp)
Ces cires rendent également les feuilles luisantes : elles absorbent la rayonnement photosynthétiquement actif tout en agissant comme un miroir (effet de réflexion spéculaire de la lumière) lorsque la radiation solaire est trop importante[4].
Structure
La cuticule est une matrice lipidique extracellulaire continue biosynthétisée par les cellules épidermiques au niveau de leur face extérieure. Elle est constituée d’un squelette de cutine et d’un mélange complexe de cires. Les parties des plantes comme les racines ou l'écorce n'ont pas de cuticule mais sont protégées par un polymère plus complexe, la subérine, formant un dépôt liégeux sur la paroi primaire de ces tissus[6].
Chez les Bryophytes
Les sporophytes et parfois les gamétophytes des bryophytes montrent une cuticule avec des stomates permettant les échanges gazeux, mais l'évolution et la biochimie de la cuticule des bryophytes, particulièrement difficile à prélever, sont peu connues[7].
Chez les plantes vasculaires
La cuticule est une couche protectrice qui recouvre les organes aériens des plantes vasculaires. Elle est composée de dépôts successifs de cire[8] enrobée dans une couche d'acides gras hydrophobes, la cutine[9] ; la cire intracuticulaire est elle-même souvent recouverte de cire épicuticulaire d'une autre nature chimique[10] - [11]. Les rôles physiologiques principaux de la cuticule sont liés à sa nature hydrophobe : d'une part elle maintient une zone pauvre en eau à la surface du végétal, ce qui protège la plante de pathogènes (notamment de la germination et du développement des spores de champignons), d'autre part elle limite les pertes de la plante en eau, en ions et en solutés polaires (sucres, acides organiques…). L'évapotranspiration se fait par la transpiration stomatique (90 à 95 %) mais aussi par la transpiration cuticulaire (environ 10 % en plus dans les régions tempérées)[12]. Ainsi, une cuticule mince (hygrophytes, tilleul) entretient cette transpiration cuticulaire alors qu'une cuticule épaisse (houx, lierre) est imperméable à l'eau. Dans ce dernier cas, la transpiration est uniquement stomatique. L'absorption de CO2 se fait par l'ouverture variable des stomates, orifices de petite taille présents dans l'épiderme des végétaux, qui permettent des échanges gazeux entre la plante et l'air ambiant. La cuticule cireuse est imperméable aux gaz et ce sont donc les stomates qui exercent un contrôle strict de la diffusion de CO2 dans la feuille[13]. Les cires épicuticulaires sont responsables des reflets bleutés ou glauques des nombreux épidermes (effet de diffraction sur leur ponctuation cuticulaire) en lien avec leur implication dans la résistance à des stress environnementaux (UV, stress hydrique, attaque de pathogènes)[14].
Certaines racines présentent aussi un épiderme recouvert d'une mince cuticule[15].
Fonctions
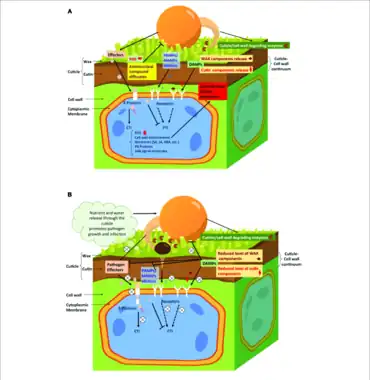
Légende : éliciteurs généraux du pathogène PAMPs/MAMPs, éliciteurs spécifiques ou effecteurs issus des pathogènes spécifiques reconnus par les récepteurs immunitaires membranaires et intracellulaires (protéines de résistance R), intervenant dans le système immunitaire inné PTI[17] et ETI[18] ; Les réponses défensives aux stress biotiques (pathogènes) au cours de l'ETI impliquent la production de molécules de signalisation SAR, la synthèse de composés antimicrobiens (ROS, protéines PR), la production d'hormones de défense (SA, JA, ABA) et le renforcement des parois cellulaires (dépôts de callose, de lignine, de subérine).
La fonction principale est d'être une barrière physique étanche qui empêche les pertes en eau massives, permettant ainsi aux stomates d’effectuer un contrôle dynamique des échanges gazeux et de la transpiration végétale[19].
Elle a comme fonctions secondaires[20] : d'être une barrière physique et chimique[21] constituant la première ligne de défense chez les plantes contre les agressions causées par des ravageurs et des agents phytopathogènes (la plupart des champignons phytopathogènes ont cependant acquis la capacité de traverser cette barrière en secrétant des enzymes spécifiques, les cutinases[22]) ; de maintenir la surface propre (auto-nettoyage par l’effet lotus grâce aux cristaux de cires épicuticulaires faisant glisser les gouttes d’eau) via les gouttes qui se chargent des spores de pathogènes (réduisant leurs attaques) et des poussières (exploitation au maximum du rayonnement lumineux) ; d'avoir un effet photoprotecteur contre les rayons UV ; d'intervenir dans les propriétés biomécaniques (conservation post récolte en limitant la dessiccation) et les aspects des fruits (brillance, pruine constituée par les cires épicuticulaires)[23].
Les adjuvants contenus dans les biocides ont entre autres pour rôle d'annuler ponctuellement cette protection afin de permettre la pénétration de principes actifs, en utilisant par exemple du Gasoil ou des détergents.
Notes et références
- Cet article est partiellement ou en totalité issu de l'article intitulé « Cuticule » (voir la liste des auteurs).
- (en) EJH Corner, The life of plants, University of Chicago Press, , p. 103-119.
- Cette cuticule recouvre les sporanges de rhyniophytes il y a 425 Ma, à la fin du Silurien. Cf (en) D. Edwards, G. Abbott, J. Raven, « Cuticles of early land plants: a palaeoecophysiological evaluation », dans G. Kersteins, Plant Cuticles: an integrated functional approach, Oxford, BIOS Scientific Publishers, , p. 1–31.
- (en) Wilhelm Barthlott, Matthias Mail, Bharat Bhushan, Kerstin Koch, « Plant Surfaces: Structures and Functions for Biomimetic Innovations », Nanomicro Lett., vol. 9, no 2, , p. 23 (DOI 10.1007/s40820-016-0125-1).
- (en) Markus Riederer, Caroline Muller, Biology of the Plant Cuticle, Wiley, , p. 5.
- L'épaisseur et la structure varient en fonction de l'espèce végétale, de l'organe de la plante, du stade de développement, et même des feuilles d'une même plante en fonction de leur orientation et leur position dans la canopée. Cf (en) CE Jeffree, « Structure and ontogeny of plant cuticles », dans G. Kersteins, Plant Cuticles: an integrated functional approach, Oxford, BIOS Scientific Publishers, , p. 37-43.
- (en) Olga Serra, Subhasish Chatterjee, Mercè Figueras, Marisa Molinas, Ruth E Stark, « Deconstructing a plant macromolecular assembly: chemical architecture, molecular flexibility, and mechanical performance of natural and engineered potato suberins », Biomacromolecules, vol. 15, no 3, , p. 799-811 (DOI 10.1021/bm401620d).
- Raven, Evert et Eichhorn 2007, p. 348
- Nultsch 1998, p. 165
- Raven, Evert et Eichhorn 2007, p. 25
- La composition précise des cires intracuticulaires est encore incertaine en raison de la difficulté rencontrée pour les isoler indépendamment des cires épicuticulaires.
- Jetter, Schäffer et Riederer 2000
- Pierre Peycru, Didier Grandperrin et Christiane Perrier, Biologie BCPST 1re année tout-en-un, Dunod, , p. 173
- Raven, Evert et Eichhorn 2007, p. 5, 668-669
- D. Robert, Jean Claude Roland, Biologie végétale : caractéristiques et stratégie évolutive des plantes, Doin, , p. 91
- Raven, Evert et Eichhorn 2007, p. 534
- (en) Carmit Ziv, Zhenzhen Zhao, Yu Gary Gao, Ye Xia, « Multifunctional Roles of Plant Cuticle During Plant-Pathogen Interactions », Frontiers in Plant Science, vol. 9, , p. 1088 (DOI 10.3389/fpls.2018.01088).
- PTI=PAMPs-triggered immunity ou immunité déclenchée par les PAMPs.
- ETI=Effector-triggered immunity ou immunité déclenchée par les effecteurs (en).
- (en) Gerhard Kerstiens, « Cuticular water permeability and its physiological significance », Journal of Experimental Botany, vol. 47, no 12, , p. 1813–1832 (DOI 10.1093/jxb/47.12.1813)
- (en) Markus Riederer, Caroline Muller, Biology of the Plant Cuticle, John Wiley & Sons, (lire en ligne), p. 2-6
- La dégradation de la cuticule par des ravageurs produit des composés spécifiques, les éliciteurs agissant comme signaux intracellulaires pour activer les défenses chimiques de la plante contre ces ravageurs. Cf (en) Mario Serrano, Fania Coluccia, Martha Torres, Floriane L’Haridon, Jean-Pierre Métraux, « The cuticle and plant defense to pathogens », Frontiers in Plant Science, vol. 5, no 274, (DOI 10.3389/fpls.2014.0027).
- (en) James A. Sweigard, Forrest G. Chumley et Barbara Valent, « Cloning and analysis of CUT1, a cutinase gene from Magnaporthe grisea », Molecular and General Genetics, vol. 232, no 2, , p. 174-182 (DOI 10.1007/BF00279994).
- (en) Trevor H Yeats, Jocelyn K C Rose, « The formation and function of plant cuticles », Plant Physiology, vol. 163, no 1, , p. 5–20 (DOI 10.1104/pp.113.222737).
Voir aussi
Bibliographie
- (en) R. Jetter, S. Schäffer et M. Riederer, « Leaf cuticular waxes are arranged in chemically and mechanically distinct layers: evidence from Prunus laurocerasus L. », Plant, Cell and Environment, vol. 23, , p. 619-628
- Wilhelm Nultsch (trad. Roger Miesch et Yves Sell), Botanique générale [« Allgemeine botanik »], Bruxelles, De Boeck Université, , 10e éd., 602 p. (ISBN 2-7445-0022-4)
- Peter H. Raven, Ray F. Evert et Susan E. Eichhorn (trad. Jules Bouharmont), Biologie végétale, Bruxelles, De Boeck Université, , 7e éd., 733 p. (ISBN 978-2-7445-0102-9 et 2-7445-0102-6)