Subérine
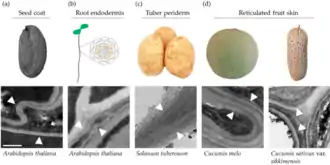
La subérine (du latin suber, « liège ») est un biopolymère lipidique imperméable qui double intérieurement les parois des cellules végétales. Cette substance cireuse est déposée dans différents tissus de plantes vasculaires pendant leur croissance et en fonction de l'espèce végétale (dépôts unicellulaires dans le tégument de graines ou l'endoderme de racines, dépôts pluricellulaires dans les péridermes aériens et souterrains, dans les zones d'abscission, les zones subéreuses du rhizoderme, l'épiderme de fruits roussis, l'épicarpe réticulé de péponides[3], le liège des écorces…)[2]. Présente de manière constitutive dans ces tissus, elle est également déposée en réponse à des blessures ou à des attaques de phytopathogènes, soulignant ainsi son rôle important dans la protection de ces plantes (notamment comme barrière contre les pertes d'eau), dans leur résistance et dans leurs relations avec l'environnement. Très stable, elle contrôle la décomposabilité et biodégradabilité des racines et joue à ce titre un rôle encore mal compris dans le cycle du carbone[4].



La subérine et la cutine sont des composés hautement hydrophobes qui entravent l'activité enzymatique des décomposeurs. Ces biopolymères sont proches l'un de l'autre par leur origine (biosynthèse qui partage des voies métaboliques communes, celle de la synthèse des acides gras et des phénylpropanoïdes (en)), leur constitution et leurs propriétés. La compréhension de leurs structures chimiques, leur biochimie et leurs fonctions soulève toujours des questions fondamentales non résolues[5].
Structure
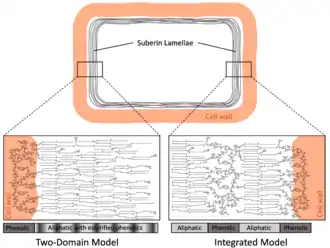
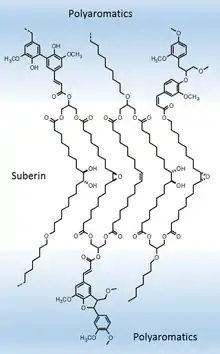
Composée de longues chaînes de polyesters d'acides gras et de composés phénoliques liés aux chaînes lipidiques par des liens esters et éthers, la subérine est le constituant principal du liège ou suber (45 % environ) et elle tient d'ailleurs son nom du Quercus suber, le chêne-liège. Elle est issue de la copolymérisation de lipides et de dérivés phénoliques[6].
Fonctions
C'est une substance originale très complexe, imperméabilisante, ignifugeante et présentant certaines propriétés biocides, dont les constituants chimiques sont assez bien connus, ce qui permet de la considérer comme un polymère lipide, mais dont la structure macromoléculaire n'est pas encore établie de façon définitive.
Avec d'autres hétéropolymères plus connus (comme les lignines et les tanins), elle permet aux plantes de lutter contre les stress biotiques et abiotiques (sécheresse, salinité, hypoxie, attaques d'agents phytopathogènes, blessures) et de maximiser les fonctions racinaires d'absorption des nutriments et de l'eau[4] - [7]. Selon une étude récente (2017) le réchauffement climatique altère la qualité de la subérine du tissu racinaire de certaines plantes (graminées en C3 ou C4, telles que Bouteloua gracilis (C4) et Hesperostipa comata (C3) [4]. Les racines de B. gracilis exposées à un taux élevé de CO2 et au réchauffement du sol augmentent leur teneur en subérine (et en lignine)[4] et moindrement celles de H. comata. Cet accroissement du taux de subérine pourrait conduire à une moindre dégradabilité des racines mortes et augmenter le taux de carbone du sol (sans que ce carbone soit biodisponible, dans un premier temps). De plus le réchauffement climatique pourrait favoriser les espèces dont les racines produisent plus de subérine, laquelle pourrait donc jouer un rôle croissant pour le réservoirs ou puits de carbone que constitue le sol[4].
Constituant principal du suber, la subérine participe aux différentes fonctions du liège.
Notes et références
- (en) Kathlyn N. Woolfson, Mina Esfandiari, Mark A. Bernards, « Suberin Biosynthesis, Assembly, and Regulation », Plants, vol. 11, no 4, , p. 555 (DOI 10.3390/plants11040555)

- (en) Gal Nomberg, Ofir Marinov, Gulab Chand Arya, Ekaterina Manasherova, Hagai Cohen, « The Key Enzymes in the Suberin Biosynthetic Pathway in Plants: An Update », Plants (Basel), vol. 11, no 3, , p. 392 (DOI 10.3390/plants11030392).
- Chez quelques variétés de Cucurbitacées (melon, concombre), le développement du fruit très volumineux induit la formation de fissures. En réponse à ces blessures, certaines cellules de l'hypoderme prolifèrent, se subérifient et provoquent l'éclatement de l'épiderme, formant en surface des verrues ou des réticulations caractéristiques. Cf (en) Hagai Cohen, Yonghui Dong, Jedrzej Szymanski, Justin Lashbrooke, Sagit Meir, Efrat Almekias-Siegl, Viktoria Valeska Zeisler-Diehl, Lukas Schreiber, Asaph Aharoni, « A Multilevel Study of Melon Fruit Reticulation Provides Insight into Skin Ligno-Suberization Hallmarks », Plant Physiology, vol. 179, no 4, , p. 1486–1501 (DOI 10.1104/pp.18.01158).
- Suseela, V., Tharayil, N., Pendall, E., Rao, A. M., & Volder, A. (2017). Warming and elevated CO2 alter the suberin chemistry in roots of photosynthetically divergent grass species. AoB PLANTS, 9(5).
- (en) Mike Pollard, Fred Beisson, Yonghua Li, John B Ohlrogge, « Building lipid barriers: biosynthesis of cutin and suberin », Trends Plant Sci., vol. 13, no 5, , p. 236-246 (DOI 10.1016/j.tplants.2008.03.003).
- (en) Beisson F, Li-Beisson Y, Pollard M., « Solving the puzzles of cutin and suberin polymer biosynthesis », Curr Opin Plant Biol., vol. 15, no 3, , p. 329-337 (DOI 10.1016/j.pbi.2012.03.003)
- (en) Lukas Schreiber, « Transport barriers made of cutin, suberin and associated waxes », Trends Plant Sci., vol. 15, no 10, , p. 546-553 (DOI 10.1016/j.tplants.2010.06.004).
Voir aussi
Articles connexes
Bibliographie
- Armas-Herrera, C. M., Dignac, M. F., Rumpel, C., Arvelo, C. D., & Chabbi, A. (2017, April). Composition and dynamics of cutin and suberin biomarkers in plants and soils under agricultural use. In EGU 2017, European Geophysical Union General Assembly 2017 (p. np)|résumé.
- Fedi F, O’Neill C.M, Menard G, Trick M, Dechirico S, Corbineau F, etc. & Penfield S (2017) Awake1, an ABC-type transporter, reveals an essential role for suberin in the control of seed dormancy Plant physiology, 174(1), 276-283.
- Camille Delude, La subérine chez Arabidopsis thaliana : Mécanisme d’export et contribution des alcools gras, thèse de doctorat en biologie végétale, Université de Bordeaux, 2015