Axelrodichthys
Axelrodichthys est un genre éteint de poissons à membres charnus rattaché à la famille des Mawsoniidae, elle-même regroupée au sein des Coelacanthiformes. Plusieurs espèces sont connues, dont les restes ont été découverts dans le Crétacé inférieur (Aptien-Albien) du Brésil[1], d’Afrique du Nord[2], et possiblement du Mexique[3] - [4], ainsi que dans le Crétacé supérieur du Maroc (Cénomanien)[5], de Madagascar (Coniacien-Santonien)[2] et de France (Campanien inférieur – Maastrichtien inférieur)[6] - [7]. Les Axelrodichthys du Crétacé inférieur fréquentaient à la fois les eaux saumâtres et les eaux marines côtières (environnement laguno-côtier) tandis que l’espèce la plus récente vivait exclusivement en eaux douces (lacs et rivières)[6] - [7]. Les spécimens français sont les derniers cœlacanthes d'eau douce connus[7]. Les espèces de ce genre mesuraient en moyenne entre 1 et 2 m de longueur[1]. Axelrodichthys fut nommé en 1986 par John G. Maisey en l’honneur de l’ichtyologiste Américain Herbert R. Axelrod[1].

Espèces de rang inférieur
Description

Comme son cousin Mawsonia, Axelrodichthys est un cœlacanthe au crâne allongé, bas et large, dont les os du toit crânien et de la joue sont caractérisés par une forte ornementation[7]. Il diffère de Mawsonia principalement par une région pariétonasale plus allongée, le développement du processus descendant du supratemporal, et par la forme et la disposition des os de la joue et de la mâchoire inférieure. Sur cette dernière, les deux branches postérieures du dentaire sont de longueur similaire chez Axelrodichthys alors que la branche inférieure de la bifurcation est beaucoup plus longue que la supérieure chez Mawsonia. La surface de contact avec l’angulaire est également plus étendue chez Axelrodichthys[8].
Taxonomie
Plusieurs espèces d’Axelrodichthys ont été décrites. La validité de certaines d’entre elles est discutée et d’autres spécimens sont laissés en nomenclature ouverte car représentés par des fossiles trop incomplets pour une détermination spécifique. Le genre comprend les taxons suivants :
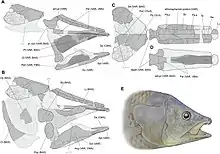
- Axelrodichthys araripensis est l’espèce type du genre. Elle est aussi l’espèce la mieux connue grâce à la découverte de nombreux spécimens entiers exceptionnellement conservés en trois dimensions dans des nodules carbonatés[1] - [8]. Ces fossiles proviennent de la formation de Crato, daté de l’Aptien, et surtout de la formation de Romualdo (anciennement un membre de la formation de Santana, cette dernière ayant été élevée au rang de groupe), à laquelle est attribué un âge albien mais qui pourrait dater de l’Aptien supérieur[9]. Ces deux formations sont situées dans le bassin d’Araripe au nord-est du Brésil (États de Ceará, Pernambouc et Piauí). Il est possible que cette espèce soit également présente dans la formation de Tlayúa (Albien supérieur) dans l’ État de Puebla au Mexique[3]. Cependant, le spécimen mentionné dans la littérature scientifique n'a jamais été décrit et a malheureusement été perdu par la suite[4].
- Axelrodichthys maiseyi provient de la Formation de Codó, datée de l’Albien moyen - supérieur, et située dans le bassin de Grajaú (état de Maranhão) au nord-est du Brésil. L’espèce est nommée d’après John G. Maisey le créateur du genre Axelrodichthys[10]. Toutefois, le statut d’A. maiseyi est discuté par certains auteurs qui doutent de l’interprétation de certaines structures anatomiques et suggèrent de réviser cette espèce[8].
- Axelrodichthys megadromos provient de diverses formations géologiques du Crétacé supérieur du sud de la France dont l’âge s’étend du Campanien inférieur au Maastrichtien inférieur. L’espèce est nommée d’après le grec megas, grand, et dromos, allée, et fait référence à la fois à l’arrivée en Europe de ce taxon originaire du Gondwana, et à la construction d’une nouvelle autoroute à proximité de la localité type[6]. A. megadromos est représenté par un crâne partiel et de nombreux os crâniens isolés provenant de plusieurs sites localisés en Provence (Bouches-du-Rhône et Var) et en Occitanie (Aude et Hérault)[7]. Les spécimens les plus anciens, tels le crâne holotypique découvert à Ventabren dans les Bouches-du-Rhône, datent du Campanien inférieur. Le spécimen le plus récent, provenant de la formation des Marnes Rouges Inférieures dans l’Aude, est daté par la magnétostratigraphie du Maastrichtien inférieur, il y a 71,5 millions d’années[6] - [7] - [11]. Cette espèce est donc le dernier représentant connu des Mawsoniidae d'eau douce[7] et l’un des derniers cœlacanthes fossiles avec le genre Megalocoelacanthus d’Amérique du Nord d’âge comparable[12] - [13] (le cœlacanthe fossile le plus récent étant un mawsoniidé fragmentaire trouvé dans les couches marines du Maastrichtien supérieur du Maroc[14]). La présence d'A. megadromos dans ce qui était l’île Ibéro-Armoricaine durant le Crétacé supérieur est la preuve d’un événement de dispersion du genre Axelrodichthys depuis le Gondwana occidental (Afrique + Amérique du Sud) vers l’archipel européen de cette époque[6] - [7].
- ? Axelrodichthys lavocati, nommé d’après le paléontologue français René Lavocat, provient de sédiments de la fin du Crétacé inférieur (Albien) et/ou du début du Crétacé supérieur (Cénomanien) du Maroc et d’Algérie[15] - [16] - [5]. Seulement connue par des os isolés, cette espèce fut d’abord attribuée au genre Mawsonia[15] - [16] avant d’être reversée dans le genre Axelrodichthys en 2019[8]. Le statut de cette espèce reste toutefois incertain car le matériel regroupé sous le nom lavocati pourrait en fait appartenir à la fois aux genres Mawsonia et Axelrodichthys[17].
Paléoécologie
Les Axelrodichthys ont vécu dans des environnements différents selon les espèces et l’époque. Durant le Crétacé inférieur, l’espèce A. araripensis habitait à la fois les eaux saumâtres et les eaux marines côtières du Gondwana occidental. En effet, la formation de Romualdo, d’où provient essentiellement cette espèce, s’est déposée dans une lagune côtière influencée par des cycles de transgressions et régressions marines et un apport variable en eau douce[9]. À la fin du Crétacé supérieur, l’espèce A. megadromos vivait exclusivement en eau douce (lacs et rivières), sur l’île Ibéro-Armoricaine, une terre émergée constituée d’une grande partie de la France et de la péninsule Ibérique[6] - [7] - [18]. Tous les sites ayant livré cette espèce ne présentent aucune influence marine. Les spécimens du Campanien inférieur proviennent de dépôts lacustres, et ceux du Campanien supérieur et du Maastrichtien inférieur furent découverts dans des sédiments de rivières et de plaines inondables[19] - [6] - [7]. Les restes d’Axelrodichthys du Crétacé supérieur du Maroc et de Madagascar proviennent également de sédiments d’eau douce[5] - [2]. L’arrivé du genre Axelrodichthys dans les écosystèmes continentaux du sud-ouest de l’Europe s’est probablement effectué à la suite de connexions terrestres ayant permis des liaisons fluviales entre l’Europe et le Gondwana[18].
Le régime alimentaire des mawsoniidés est encore mal connu. Bien que des dents minuscules soient présentes sur le palais et la partie interne de la mandibule, la bouche de ces poissons est essentiellement édentée. Aussi certains auteurs ont supposé qu’ils devaient se nourrir en aspirant leurs proies à la manière du Latimeria actuel. D’autres chercheurs ont proposé un mode d’alimentation par filtration des nutriments[20]. La description en 2018 d’un spécimen articulé d’A. araripensis ayant avalé un poisson entier semble confirmer le mode d’alimentation par aspiration[21].
Classification phylogénique
Position du genre Axelrodichthys de la famille éteinte des Mawsoniidae[22] :
Une analyse phylogénétique de la famille publiée en 2020 a trouvé une polytomie regroupant les genres crétacés “Lualabaea”, Axelrodichthys, Mawsonia, ainsi que le genre marin du Jurassique Trachymetopon. “Lualabaea” pourrait être congénérique avec Axelrodichthys[7].
| |||||||||||||||||||||||||||||||||||||||||||||||||
Références taxinomiques
- (en) Référence Paleobiology Database : Axelrodichthys Maisey, 1986
Notes et références
- (en) J.G. Maisey, « Coelacanths from the Lower Cretaceous of Brazil », American Museum Novitates, vol. 2866, , p. 1-30 (lire en ligne)
- (en) M.D. Gottfried, Rogers, R.R. et Curry Rogers, K., « First record of Late Cretaceous coelacanths from Madagascar », dans Arratia, G., Wilson, M..H., Cloutier, R., Recent Advances in the Origin and Early Radiation of Vertebrates, München, Verlag Dr. Friedrich Pfeil, (ISBN 978-3-89937-052-2), p. 687–691
- (en) L. Espinosa-Arrubarrena, S.P. Applegate et K. González-Rodríguez, « The first Mexican record of a coelacanth (Osteichthyes: Sarcopterygii) from the Tlayua quarries near Tepexi de Rodríguez, Puebla, with a discussion on the importance of this fossil: Sixth North American Paleontological Convention, Abstracts of Papers », Paleontological Society Special Publication, vol. 8, , p. 116 (DOI 10.1017/S2475262200001180)
- (en) K.A. González-Rodríguez, Ch. Fielitz, V.M. Bravo-Cuevas et R.E. Baños-Rodríguez, « Cretaceous osteichthyan fish assemblages from Mexico », New Mexico Museum of Natural History and Science Bulletin, vol. 71, , p. 107-119
- (en) L. Cavin, L. Boudad, H. Tong, E. Läng, J. Tabouelle et R. Vullo, « Taxonomic composition and trophic structure of the continental bony fish assemblage from the Early Late Cretaceous of Southeastern Morocco », PLoS ONE, vol. 10(5), , e0125786 (PMID 26018561, PMCID 4446216, DOI 10.1371/journal.pone.0125786)
- (en) L. Cavin, X. Valentin et G. Garcia, « A new mawsoniid coelacanth (Actinistia) from the Upper Cretaceous of Southern France », Cretaceous Research, vol. 62, , p. 65-73 (DOI 10.1016/.cretres.2016.02.002)
- (en) L. Cavin, E. Buffetaut, Y. Dutour, G. Garcia, J. Le Loeuff, A. Méchin, P. Méchin, H. Tong, T Tortosa, E. Turini et X. Valentin, « The last known freshwater coelacanths: New Late Cretaceous mawsoniids remains (Osteichthyes: Actinistia) from Southern France », PLoS ONE, vol. 15(6), , e0234183 (PMID 32502171, PMCID 7274394, DOI 10.1371/journal.pone.0234183)
- (en) L.G.C. Fragoso, P. Brito et Y. Yabumoto, « Axelrodichthys araripensis Maisey, 1986 revisited », Historical Biology, vol. 31(10), , p. 1350-1372 (DOI 10.1080/08912963.2018.1454443)
- (en) M.A. Custódio, F. Quaglio, L.V. Warren, M.G. Simões, F.T. Fürsich, J. A. Perinotto et M.L. Assine, « The transgressive-regressive cycle of the Romualdo Formation (Araripe Basin): Sedimentary archive of the Early Cretaceous marine ingression in the interior of Northeast Brazil », Sedimentary Geology, vol. 359, , p. 1-15 (DOI 10.1016/j.sedgeo.2017.07.010)
- (en) M.S.S. de Carvalho, V. Gallo et H.R.S. Santos, « New species of coelacanth fish from the Lower Cretaceous (Albian) of the Grajaú Basin, NE Brazil », Cretaceous Research, vol. 46, , p. 80-89 (DOI 10.1016/j.cretres.2013.09.006)
- (en) V. Fondevilla, J. Dinarès-Turell, B. Vila, J. Le Loeuff, R. Estrada, O. Oms et A. Galobart, « Magnetostratigraphy of the Maastrichtian continental record in the Upper Aude Valley (northern Pyrenees, France): Placing age constraints on the succession of dinosaur-bearing sites », Cretaceous Research, vol. 57, , p. 457-472 (DOI 10.1016/j.cretres.2015.08.009)
- (en) D.R. Schwimmer, J.D. Stewart et G.D. Williams, « Giant fossil coelacanths of the Late Cretaceous in the eastern United States », Geology, vol. 22(6), , p. 503-506 (DOI 10.1130/0091-7613(1994)022<0503:GFCOTL>2.3.CO;2)
- (en) H. Dutel, J.P. Maisey, D.R. Schwimmer, P. Janvier, M. Herbin et G. Clément, « The giant Cretaceous coelacanth (Actinistia, Sarcopterygii) Megalocoelacanthus dobiei Schwimmer, Stewart & Williams, 1994, and its bearing on Latimerioidei interrelationships », PLoS ONE, vol. 10(5), , e49911 (PMID 23209614, DOI 10.1371/journal.pone.0049911)
- (en) P.M. Brito, D.M. Martill, I. Eaves, R.E. Smith et S.L.A. Cooper, « A marine Late Cretaceous (Maastrichtian) coelacanth from North Africa », Cretaceous Research, vol. 122, , p. 104768 (DOI 10.1016/j.cretres.2021.104768)
- (en) L. Cavin et Forey, P.L., « New mawsoniid coelacanth (Sarcopterygii: Actinistia) remains from the Cretaceous of the Kem Kem beds, SE Morocco », dans Arratia, G. & Tintori, A., Mesozoic Fishes III, Systematics, Palaeoenvironments and Biodiversity, München, Verlag Dr. Friedrich Pfeil, (ISBN 978-3-89937-053-9), p. 493–506
- (en) Y. Yabumoto et T. Uyeno, « New Materials of a Cretaceous Coelacanth, Mawsonia lavocati Tabaste from Morocco », Bulletin of the Natural Science Museum, Tokyo, Series C, vol. 31, , p. 39-49 (lire en ligne)
- (en) L. Cavin, C. Cupello, Y. Yabumoto, L.G.C. Fragoso, U. Deesri et P.M. Brito, « Phylogeny and evolutionary history of mawsoniid coelacanths », Bulletin of the Kitakyushu Museum of Natural History and Human History, Series A, vol. 17, , p. 3-13 (lire en ligne)
- (en) Z. Csiki-Sava, E. Buffetaut, A. Ősi, X. Pereda-Suberbiola et S.L. Brusatte, « Island life in the Cretaceous-faunal composition, biostratigraphy, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago », ZooKeys, vol. 469, , p. 1-161 (DOI 10.3897/zookeys.469.8439, lire en ligne)
- (en) L. Cavin, P.L. Forey, H. Tong et E. Buffetaut, « Latest European coelacanth shows Gondwanan affinities », Biology Letters, vol. 1(2), , p. 176-177 (DOI 10.1098/rsbl.2004.0287)
- (en) L. Cavin, « Histoires évolutives des poissons dulçaquicoles », dans Cavin, L., Les poissons d’eau douce : 250 millions d’années d’histoire évolutive, Croydon, Iste, (ISBN 978-1-78405-398-7, BNF 45515862), p. 69–75
- (en) F.J. Meunier, C. Cupello, Y. Yabumoto et B.M. Brito, « The diet of the Early Cretaceous coelacanth Axelrodichthys araripensis Maisey, 1986 (Actinistia: Mawsoniidae) », Cybium, vol. 42(1), , p. 105-111 (DOI 10.26028/cybium/2018-421-011)
- (en) Gaël Clément, « A New Coelacanth (Actinistia, Sarcopterygii) from the Jurassic of France, and the question of the closest relative fossil to Latimeria », Journal of Vertebrate Paleontology, vol. 25, No. 3 (sep. 30, 2005), pp. 481-491, https://www.jstor.org/stable/4524470?seq=1#page_scan_tab_contents