Évolution du système nerveux
L'évolution du système nerveux débute avec l'apparition des premiers systèmes nerveux chez les animaux (ou métazoaires). Les neurones, des cellules spécialisés dans la signalisation électrique, apparaissent chez les animaux multicellulaires, en conservant le mécanisme des potentiels d'action déjà présents chez des cellules unicellulaires mobiles et des colonies d'eucaryotes. On retrouve dans le règne animal des réseaux nerveux diffus simples (dépourvus de centralisation ou de céphalisation) comme chez les cnidaires évolués, des cordons nerveux chez les animaux bilatéraux – dont des cordons nerveux ventraux (en) chez les invertébrés et des cordons nerveux dorsaux (en) provenant de la notochorde chez les chordés. Pour finir, la bilatéralisation a conduit à l'apparition du cerveau, par un processus complexe appelé céphalisation.
Précurseurs neuronaux
Les potentiels d'action sont nécessaires pour l'activité neuronale et sont apparus en premier chez les eucaryotes unicellulaires. Ces potentiels « primitifs » utilisent le calcium plutôt que le sodium. Les scientifiques pensent que le rôle des potentiels d'action dans la signalisation neuronale électrique est cependant apparu chez les animaux multicellulaires. Dans certaines colonies d'eucaryotes, tels que les obelias, les signaux électriques ne se propagent pas seulement à travers les réseaux de neurones, mais aussi à travers les cellules épithéliales du système digestif commun à toute la colonie[1].
Éponges
Les éponges ne possèdent pas de cellules reliées par des jonctions synaptiques, ce qui implique l’absence de neurones, et donc de système nerveux. Cependant, elles présentent des homologues de nombreux gènes qui jouent un rôle clé dans le développement des synapses. Des études récentes ont montré que les cellules des éponges expriment un groupe de protéines qui s'assemblent pour former une structure ressemblant à une postsynaptic density (PSD) (localisée dans la partie réceptrice du signal). Cependant, la fonction de cette structure est mal déterminée. Bien que les cellules de l'éponge n'utilisent pas de transmission synaptique, elles communiquent bien les unes avec les autres via des ondes de calcium et diverses impulsions, qui permettent quelques actions simples telle que la contraction générale du corps.
Réseau nerveux diffus
.jpg.webp)
Les cnidaires (comme les méduses ou les hydres[2] - [3]), les cténophores, et autres animaux en relation possèdent un réseau nerveux diffus (nerve nets en anglais) plutôt qu'un système nerveux central. Chez la plupart des méduses le réseau nerveux diffus est réparti plus ou moins uniformément dans le corps; chez les cténophores il est concentré près de la bouche. Ce réseau nerveux diffus est constitué de neurones sensoriels qui réagissent à des signaux chimiques, tactiles et visuelles; de neurones moteurs qui peuvent déclencher des contractions à la surface du corps; et des neurones dits « intermédiaires » qui reconnaissent les schémas d'activité des neurones sensoriels et envoient les signaux appropriés à des groupes de neurones moteurs. Dans certains cas, ces neurones intermédiaires se regroupent sous forme de ganglions.
Chez les radiatas, le développement du système nerveux est relativement peu structuré. Contrairement aux bilateriens, les radiatas ont seulement deux couches de cellules primordiales, l'endoderme et l'ectoderme (absence de mésoderme). Les neurones apparaissent à partir d'un groupe spécial de cellules ectodermiques, qui sert également de précurseurs pour toutes les autres cellules ectodermiques[4].
Cordons neuraux

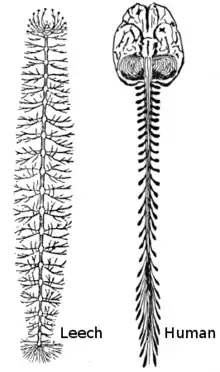
La grande majorité des animaux sont bilatériens, avec un côté gauche et droit qui sont approximativement images l'un de l'autre dans un miroir. Tous les bilatériens ont pour ascendant commun un ancêtre vermiforme apparu durant la période édiacarienne (allant de -550 à -600 millions d'années). La forme fondamentale des bilatériens adultes est celle d'un tube possédant une cavité abdominale allant de la bouche à l'anus et un cordon nerveux avec des élargissements (des « ganglions ») pour chaque segment du corps, avec un plus grand ganglion à l'avant qualifié de « cerveau ». Le cordon nerveux (structure « primitive » définitive) ne doit pas être confondu avec le tube neural (structure embryonnaire transitoire) même si les deux ont un lien très proche. Pour aider à la représentation de ces cordons neuraux, chez l'humain le cordon neural principal peut être grossièrement associé à la moelle épinière.
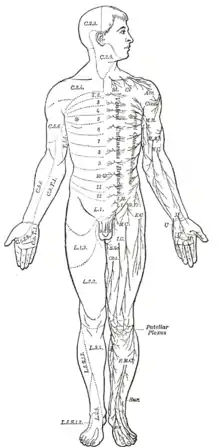
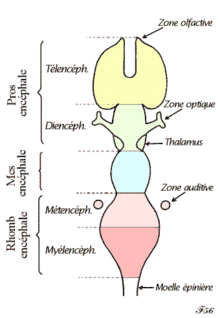
Même les mammifères, y compris les humains, montrent une segmentation bilatérienne du corps (notamment au niveau du système nerveux). La moelle épinière contient une série de ganglions spinaux d'organisation segmentaire, chacun donnant lieu à des nerfs sensitifs et moteurs qui innervent une partie superficielle du corps ainsi que la musculature sous-jacente. Cette organisation pour les membres suit un modèle complexe, sauf pour le tronc dont les zones innervées apparaissent comme une succession de bandes étroites. Les trois premiers segments appartiennent au cerveau, donnant naissance au prosencéphale, au mésencéphale et au rhombencéphale[5].
Les bilatériens peuvent être divisés sur la base des événements qui se produisent très tôt dans le développement embryonnaire, en deux groupes (phylum) appelés les protostomiens et les deutérostomiens.
- Les deutérostomiens incluent les vertébrés ainsi que les échinodermes, les hémichordés, et les xenoturbellidiens.
- Les protostomiens, le groupe le plus diversifié, incluent les arthropodes, les mollusques et de nombreux types de vers.
Il y a une différence fondamentale entre les deux groupes concernant la mise en place du système nerveux : les protostomiens possèdent un cordon nerveux sur la face ventrale (généralement en bas), tandis que chez les deutérostomiens le cordon est dorsal (généralement en haut). En fait, de nombreux aspects de l'organisme sont inversés entre les deux groupes, notamment les zones d'expression de plusieurs gènes participant au gradient dorso-ventral. La plupart des anatomistes contemporains considèrent que le corps des protostomiens et des deutérostomiens est « retourné » l'un par rapport à l'autre, une hypothèse proposée pour la première fois par Geoffroy Saint-Hilaire entre insectes et vertébrés. Ainsi les insectes ont, par exemple, les cordons neuraux le long de la ligne médiane ventrale du corps, tandis que chez tous les vertébrés la moelle épinière court le long de la ligne médiane dorsale.
Vers (protostomien)
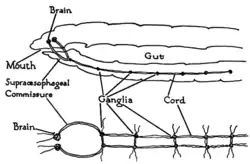
Les vers sont les animaux bilatériens les plus simples, et comprennent une structure nerveuse bilatérienne basique des plus directes[6]. Par exemple, les vers de terre ont un double cordon neural qui suit la longueur du corps et fusionne au niveau de la queue et de la bouche. Ces cordons neuraux sont reliées par des nerfs transverses comme les barreaux d'une échelle. Ces nerfs transverses aident à coordonner les deux côtés de l'animal. Deux ganglions situés à la tête (portion terminale) ont une fonction semblable à celle d'un cerveau primitif. Les photorécepteurs à l'avant captent des informations sensorielles lumineuses[7].
Le système nerveux d'un très petit ver, le ver rond Caenorhabditis elegans, a été cartographié jusqu'au niveau synaptique. Chaque neurone avec sa descendance cellulaire a été enregistré et toutes les connexions neuronales sont connues. Dans cette espèce, le système nerveux entraîne un dimorphisme sexuel ; les systèmes nerveux des deux sexes, mâles et hermaphrodites, ont des neurones spécifiques et différents responsables des fonctions sexuelles. Chez Caenorhabditis elegans, les mâles ont exactement 383 neurones, tandis que les hermaphrodites en ont exactement 302.
Arthropodes (protostomien)
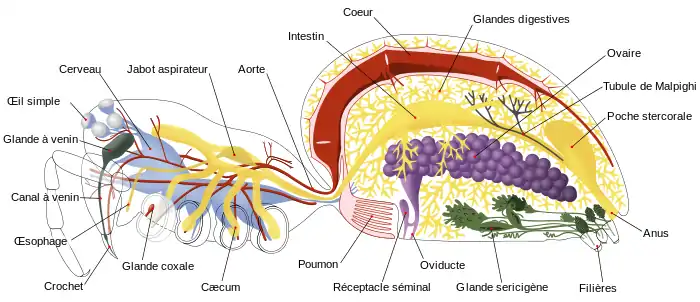
Les arthropodes, tels que les insectes et les crustacés, ont un système nerveux constitué d'une série de ganglions, reliés par un cordon nerveux ventral constitué de deux jonctions parallèles[8]. En règle générale, chaque segment du corps est lié à un ganglion de chaque côté, bien que certains ganglions fusionnent pour former le cerveau et des ganglions plus gros. Le segment crânial contient le cerveau, aussi connu sous le nom de ganglion supraesophageal (voir en anglais). Chez les insectes, le cerveau est anatomiquement divisé en protocérébrum, deutocérébrum, et tritocérébrum. Juste derrière le cerveau se trouve le ganglion subesophageal (voir en anglais), qui est composé de trois paires de ganglions fusionnés. Il contrôle les zones buccales, les glandes salivaires et certains muscles. De nombreux arthropodes ont développé des organes sensoriels comme des yeux pour la vision, et des antennes pour l'olfaction et la reconnaissance des phéromones. Les informations sensorielles provenant de ces organes sont traitées par le cerveau.
Chez les insectes, de nombreux neurones ont leur corps cellulaire positionné à la périphérie du cerveau et sont électriquement passifs — le corps cellulaire ne sert qu'à assurer les fonctions métaboliques et ne participe pas à la signalisation. Une fibre protoplasmique part du corps cellulaire et crée de nombreux branchements avec quelques pièces responsables de la transmission des signaux et d'autres pièces responsables de la réception des signaux. Ainsi la plupart des parties du cerveau des insectes possèdent des neurones passifs en périphérie, tandis que les neurones dont le rôle est le traitement du signal sont rassemblés dans un enchevêtrement de fibres protoplasmiques (axone et dendrite) appelé neuropile[9].
Amphibiens
L'iode et le T4 stimulent l'apoptose (mort cellulaire programmée) chez les cellules larvaires des bronches, de la queue et des nageoires, et stimulent aussi l'évolution du système nerveux transformant un têtard (presque toujours) végétarien aquatique en une grenouille carnivore terrestre avec des meilleures capacités neurologiques, visuo-spatiales et olfactives pour la chasse.
À l'opposé de la métamorphose des amphibiens, la thyroïdectomie et l'hypothyroïdie des mammifères peuvent être considérées comme une sorte de régression phylogénétique, métabolique et neurologique vers un stade reptilien primitif. Ainsi, de nombreux troubles qui affectent les patients atteints de problème d'hypothyroïdie montrent des caractéristiques reptiliennes, telles qu'un ralentissement général des réflexes nerveux; une léthargie du métabolisme, de la digestion et de la fréquence cardiaque; une hypothermie; et une peau sèche, glabre, écailleuse et froide[10] - [11].
Évolution des comportements innés
Des comportements tels que le "tail-flip" escape reaction des crustacés (comme les écrevisses et les homards) sont des schèmes d’action spécifique qui peuvent avoir évolué à partir d'anciennes habitudes ancestrales.
Évolution du système nerveux central
Évolution du cerveau humain
Il y a eu une augmentation progressive du volume cérébral tout au long de l'histoire évolutive de la lignée humaine, d'environ 600 cm3 chez Homo habilis à environ 1 600 cm3 chez l'Homme de Néandertal. À taille du corps égale, il existe une corrélation brute entre volume du cerveau et intelligence[12]. La dernière phase de l'évolution humaine montre cependant un schéma plus complexe : l'Homme moderne a un cerveau plus petit (1 350 cm3 en moyenne) que celui des Néandertaliens.
Iain Davidson résume la loi régissant le volume du cerveau par l'expression « aussi grand que nécessaire et aussi petit que possible »[13].
L'évolution du cerveau peut être étudiée à l'aide de la technique de l'endocaste, un moulage de l'intérieur de la boite crânienne, utilisée en paléoneurologie.
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Evolution of nervous systems » (voir la liste des auteurs).
- (en) Gary G. Matthews, Neurobiology : molecules, cells, and systems, Wiley-Blackwell, , 592 p. (ISBN 0-632-04496-9, lire en ligne), « Evolution of nervous systems », p. 21
- (en) THOMAS L. LFNTZ et RUSSFLL J. BARRNETT, « FINE STRUCTURE OF THE NERVOUS SYSTEM OFHYDRA », American Zoologist, vol. 5, no 3, , p. 341–356 (ISSN 1540-7063, DOI 10.1093/icb/5.3.341, lire en ligne, consulté le )
- (en) Allison L. Burnett et Norma A. Diehl, « The nervous system of hydra. I. Types, distribution and origin of nerve elements », Journal of Experimental Zoology, vol. 157, no 2, , p. 217–226 (ISSN 1097-010X, DOI 10.1002/jez.1401570205, lire en ligne, consulté le )
- (en) ouvrage collectif (?), Development of the nervous system, Academic Press, , 3–4 p. (ISBN 978-0-12-618621-5)
- (en) Ghysen A, « The origin and evolution of the nervous system », Int. J. Dev. Biol., vol. 47, nos 7–8, , p. 555–62 (PMID 14756331, lire en ligne)
- « Ediacaran fauna worms », Walter Jahn (consulté le ) – Suny Orange
- ADEY WR, « The nervous system of the earthworm Megascolex », J. Comp. Neurol., vol. 94, no 1, , p. 57–103 (PMID 14814220, DOI 10.1002/cne.900940104)
- Chapman RF, The insects : structure and function, Cambridge University Press, , 533–568 p. (ISBN 978-0-521-57890-5, lire en ligne), « Ch. 20: Nervous system »
- Chapman, p. 546
- Venturi, Sebastiano, « Evolutionary Significance of Iodine », Current Chemical Biology-, vol. 5, no 3, , p. 155–162 (ISSN 1872-3136, DOI 10.2174/187231311796765012)
- Venturi, Sebastiano, « Iodine, PUFAs and Iodolipids in Health and Disease: An Evolutionary Perspective », Human Evolution-, vol. 29 (1-3), , p. 185–205 (ISSN 0393-9375)
- Kwang Hyun Ko, « Origins of human intelligence : The chain of tool-making and brain evolution », Anthropological Notebooks, vol. 22, no 1, , p. 5–22 (lire en ligne)
- Iain Davidson, « As large as you need and as small as you can. Implications of the brain size of Homo floresiensis », Une-au.academia.edu