Origine multirégionale de l'homme moderne
Selon la théorie de l'origine multirégionale de l'homme moderne, ou de continuité avec hybridation, proposée et défendue depuis 1984 par l'américain Milford H. Wolpoff, l'évolution de l'homme jusqu'à nos jours se serait faite à partir d'Homo ergaster, apparu il y a environ 2 millions d'années en Afrique, et se serait déroulée au sein d'une espèce humaine unique présente dans les différentes régions du monde sous des formes diverses. Le chemin d'évolution vers l'Homme moderne ne se serait pas fait par sélection de groupe et remplacements successifs de populations, mais par une évolution à foyers multiples diffusée par de nombreux croisements entre populations et jalonnée par la sélection individuelle. Parfois nommée Out of Nowhere, cette théorie propose un « grand métissage » entre les différentes populations humaines de la planète depuis 2 millions d'années[1] - [2].
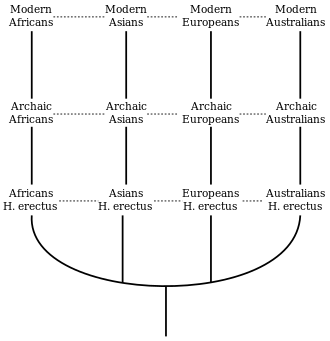
La théorie concurrente, l'origine africaine de l'Homme moderne (également connue sous le nom d'Out of Africa), a émergé comme un quasi-consensus depuis les années 1990[3] - [4].
Définition
L'hypothèse multirégionale est un modèle scientifique proposé par Milford H. Wolpoff en 1984 pour rendre compte du schéma de l'évolution humaine[5] - [6] - [7] - [8]. Cette théorie soutient que l'évolution humaine, du début du Pléistocène il y a 2 millions d'années jusqu'à nos jours, s'est faite au sein d'une seule espèce humaine, dont les différentes populations auraient évolué en parallèle partout dans le monde vers Homo sapiens. Cette espèce unique englobe toutes les formes humaines archaïques, telles qu'Homo erectus et l'Homme de Néandertal, ainsi que l'Homme moderne. La théorie repose sur le mécanisme de cline systématique, avec un équilibre entre la dérive génétique, le flux de gènes et la sélection naturelle tout au long du Pléistocène, permettant ainsi une évolution parallèle des différents groupes humains répartis autour du monde, mais avec le maintien de différences régionales, qu'elles soient extérieurement visibles ou non[9]. Le modèle de l’évolution multirégionale s’appuie sur l’observation de traits jugés communs entre fossiles de différentes époques dans une même région.
Histoire
Vue d'ensemble
L'hypothèse multirégionale a été proposée en 1984 par Milford H. Wolpoff, Alan Thorne (en) et Xinzhi Wu (en)[10] - [11] - [12]. L'hypothèse « polycentrique » des origines de l'homme de Franz Weidenreich (« évolution polycentrique » selon laquelle l'espèce Homo sapiens aurait émergé de façon parallèle et indépendante, à partir de populations d'Homo erectus, dans quatre parties du monde) conceptualisée en 1939, a été une influence majeure pour l’élaboration de l’hypothèse multirégionale, mais Wolpoff l'affine en supposant l'existence d'une unique espèce humaine, qui aurait évolué vers Homo sapiens partout dans le monde. L'humanité actuelle aurait eu plusieurs berceaux, reliés les uns aux autres à la suite du nomadisme des populations (les différentes populations se seraient homogénéisées par le jeu de migrations.
Wolpoff avertit que la théorie polycentrique ne doit pas être confondue avec le polygénisme de Carleton Coon (un des partisans les plus connus de la pensée raciste évolutionniste) qui minimise le flux de gènes[12] - [13] - [14]. Selon Wolpoff, l'hypothèse multirégionale a été mal interprétée par William W. Howells dans ses publications pendant cinquante ans, qui a confondu hypothèse de Weidenreich avec un « modèle de candélabres » polygénique :
« Pourquoi l'hypothèse multirégionale a été accusée d’être une forme de polygénie ? Nous pensons que la confusion des idées de Weidenreich et des nôtres avec Coon en est à l’origine. La raison historique reliant Coon aux idées de Weidenreich sont les interprétations erronées du modèle polycentrique de Weidenreich (Howells, 1942, 1944, 1959, 1993). Ces interprétations rendant par là même le modèle polycentrique beaucoup plus semblable à Coon qu'il ne l'est en réalité »[15].
À cause de l'influence de William W. Howells, beaucoup d'anthropologues et de biologistes ont confondu l’hypothèse du multirégionalisme avec le polygénisme. Alan Templeton note par exemple que cette confusion originale a conduit à la croyance généralisée que le flux génétique entre les différentes populations eut été ajouté à l'hypothèse multirégionale « en réponse aux critiques du modèle », malgré le fait que « l'évolution parallèle n'ait jamais fait partie du modèle multirégional, tandis que le flux de gènes n’a pas été un ajout récent, mais était présent dans le modèle dès le début »[16]. Malgré cela, le multirégionalisme est toujours confondu avec polygénisme, ou modèle de Coon des origines raciales, à partir desquels Wolpoff et ses collègues ont pourtant pris leurs distances[17] - [18]. Wolpoff a également défendu l'hypothèse polycentrique de Weidenreich contre l’étiquetage de polyphylétique. Weidenreich lui-même en 1949 a écrit : « Je cours le risque d'être mal compris, […] je crois en l'évolution polyphylétique de l'homme »[19].
En 1998, Wu propose un modèle multirégional spécifique à la Chine appelé « continuité avec hybridation [accidentelle] »[20] - [21]. La variante de Wu applique l'hypothèse multirégionale aux fossiles trouvés en Asie de l'Est, ce qui est populaire parmi les scientifiques chinois[22]. Pour James Leibold, historien politique de la Chine moderne, le soutien apporté au modèle proposé par Wu est largement ancré dans le nationalisme chinois[23]. En 2013, une étude rapportait que l’hypothèse multirégionale avait très peu de soutien au sein de la communauté scientifique en dehors de la Chine[24].
Multirégionalisme faible contre multirégionalisme classique
Christopher Brian Stringer, un des principaux promoteurs de la théorie de l'origine africaine récente, a produit une série de publications tout au long des années 1980 et 1990 à propos du multirégionalisme de Wolpoff et Thorne [25] - [26] - [27] - [28]. D’après lui, l'hypothèse de l'origine multirégionale a été modifiée au fil du temps en une variante plus « faible » qui accorde désormais un rôle plus important à l'Afrique dans l’histoire de l'évolution humaine, y compris la modernité anatomique (et par conséquent moins de continuité régionale que proposé au début)[29].
Stringer distingue le modèle multirégional des origines « classique » ayant existé de 1984 (sa formulation) jusqu'en 2003 et un modèle multirégional « faible » développé après 2003[30] - [31].
Les données fossiles
Clades morphologiques


Les partisans de l'hypothèse multirégionale voient une continuité régionale de certains caractères morphologiques à travers le Pléistocène dans différentes régions du monde. Ce qui constituerait un indice que le modèle unique de remplacement à partir de l'Afrique serait erroné. En général, trois grandes régions sont distinguées : Europe, Chine et Indonésie (comprenant souvent l'Australie)[32] - [33] - [34]. Wolpoff prend la précaution de nommer la continuité de certaines caractéristiques du squelette dans ces régions « clades morphologiques » et de ne pas considérer de contexte racial. Il définit ces clades morphologiques comme un ensemble de traits qui « caractérisent de façon unique une région géographique »[35]. Wolpoff et Thorne écrivent (1981): « Nous ne considérons pas chaque clade morphologique comme une lignée unique et nous ne croyons pas nécessaire de créer une taxonomique particulière pour ces clades »[36].
Les critiques du multirégionalisme ont rapporté qu'aucun trait humain n’est propre à une région géographique (c.-à-d. confiné à une seule population et ne se retrouvant dans aucune autre). Ce à quoi Wolpoff et al. (2000) répondirent que le modèle de continuité régionale n’identifie que la présence de combinaisons de caractères et non pas des traits individuels. Un point qu’ils comparent d'ailleurs à l'identification médico-légale d'un squelette humain :
« « Le modèle de continuité régional [...] ne postule pas que telles ou telles caractéristiques soient exclusives à une région et ne se retrouvent pas ailleurs. La structure génétique de l'espèce humaine rend par ailleurs une telle possibilité extrêmement peu probable. Il peut cependant y avoir une originalité au sens d’unicité dans des combinaisons de traits. Mais pas un seul trait n’est susceptible d'avoir été unique dans une partie du monde mais s’il semble l’être, probablement en raison du faible nombre de fossiles humains connus ». »
Les combinaisons de caractéristiques sont « uniques » au sens de n’avoir été trouvées que dans une seule région, ou à haute fréquence dans une région et très rarement dans les autres. Wolpoff souligne que la continuité régionale fonctionne en incluant des échanges génétiques entre les populations. La continuité régionale à long terme de certains traits morphologiques est alors expliquée par le modèle d'Alan Thorne « Centre et Edge »[37]. Ce modèle d’évolution génétique de population résout le paradoxe de Weidenreich « comment des populations qui ne se mélangent pas géographiquement, évoluent ensemble ? ». Par exemple en 2001, Wolpoff et ses collègues ont publié une analyse de crânes fossiles d’hommes modernes trouvés en Australie et en Europe centrale. Ils concluent que la diversité des caractéristiques morphologiques de ces crânes humains ne peut « résulter exclusivement de la dispersion de la fin du Pléistocène », ce qui implique une double ascendance pour chaque région, impliquant le croisement avec des ancêtres venant d’Afrique et des humains locaux[38].
Indonésie, Australie
Dans ses publications, Alan Thorne décrit une continuité régionale en Indonésie et en Australie pour un clade morphologique[39] - [40]. Ce clade débute avec l'Homme de Sangiran, se poursuit avec l'Homme de Solo, et inclut pour finir les aborigènes d'Australie préhistoriques et contemporains. En 1991, Andrew Kramer teste les 17 caractéristiques morphologiques proposé du clade. Il constate que : « huit des dix-sept caractéristiques […] relient Sangiran aux Australiens modernes » et que ces liens « sont révélateurs d’une continuité morphologique. Ce qui implique la présence d'un continuum génétique en Australasie remontant au moins à un million d'années »[41]. Dans une contre-étude, Colin Groves critique la méthodologie de Kramer, soulignant que la polarité des caractéristiques n'ayant pas été testée, l'étude ne peut pas être concluante[42]. Phillip Habgood a également découvert que les caractéristiques supposés être uniquement présentes dans la région de l'Australasie étudiée par Thorne, seraient en fait plésiomorphes :
« « ... il est évident que tous les caractères proposés ... pour constituer les « caractéristiques du clade » reliant l’Homo erectus indonésien aux crânes aborigènes australiens sont en réalité présents chez de nombreux Homo erectus et Homo sapiens archaïques. Beaucoup sont aussi communément trouvées sur les crânes et mandibules d'Homo sapiens anatomiquement modernes provenant d'autres zones géographiques et particulièrement sur le matériel squelettique du mésolithique d'Afrique du Nord »[43]. »
Cependant, en dépit de ces critiques, Habgood (2003) reconnait une continuité régionale limitée en Indonésie et en Australie, reconnaissant quatre caractéristiques dont la combinaison est unique : un os frontal plat, une constriction postorbitaire minime, un certain prognathisme facial, et des tubérosités zygomaxillaires[44]. Cette combinaison, selon Habgood, a une « certaine australinité ».
Wolpoff, au départ sceptique vis-à-vis des déclarations de Thorne, se laisse ensuite convaincre par la reconstruction du crâne d’Homo erectus Sangiran 17 d'Indonésie. Il est particulièrement frappé alors par la similarité de l’angle formé par la voute crânienne et le visage chez l'Homo sapiens australien trouvé sur le site archéologique de Kow Swamp 1 et notamment son prognathisme important. Durband (2007) en revanche affirme que les « caractéristiques » montrant la continuité entre Sangiran 17 et Kow Swamp 1 ont disparu dans la nouvelle reconstruction plus orthognathique de ce fossile qui a été récemment achevée »[45]. Baba et al., qui avaient restauré le visage de Sangiran 17, concluent : « la continuité régionale en Australasie est beaucoup moins évidente que Thorne et Wolpoff le font valoir »[46].
Chine

Xinzhi Wu a plaidé en faveur d'un clade morphologique en Chine enjambant le Pléistocène, caractérisé par une combinaison de dix caractéristiques[47] - [48]. La séquence commencerait avec l'homme de Lantian et l'homme de Pékin puis se poursuivrait avec l’Homme de Dali, des spécimens du Pléistocène (par exemple, Liujiang) jusqu’aux chinois modernes. Habgood en 1992 a critiqué la liste de Wu, soulignant que la combinaison de la plupart des dix fonctionnalités apparaissait régulièrement sur des fossiles trouvés à l’extérieur de la Chine[49]. Cependant, il note que la combinaison des trois caractéristiques suivantes : une racine nasale non-déprimé, des os du nez non saillant orientés perpendiculairement et une planéité du visage est unique à la région chinoise dans le registre fossile et peut être une preuve de continuité régionale limitée. Toutefois, selon Chris Stringer, l'étude de Habgood souffre de ne pas avoir inclus suffisamment d'échantillons de fossiles d'Afrique du Nord, dont beaucoup présenteraient la combinaison considérée comme spécifique à la Chine[28].
La planéité du visage comme une caractéristique de clade morphologique a été rejetée par de nombreux anthropologues car il se trouve sur de nombreux fossiles des premiers Homo erectus africains, et est donc considéré plésiomorphe[50], mais Wu a répondu que la forme de la planéité du visage dans le registre fossile chinoise apparaît distincte d'autres formes. Toetik Koesbardiati dans sa thèse de doctorat « sur la pertinence des continuités régionales du visage en Asie de l'Est » a également constaté qu'une forme de planéité du visage est unique à la Chine (c.-à-d. présente à haute fréquence et très rare ailleurs), mais nuance en notant que c’est la seule preuve disponible pour la continuité régionale: « Seules deux caractéristiques semblent conformes au modèle multirégional : la planéité à la face supérieure exprimée par un angle naso-frontal obtus et la planéité à la partie moyenne de la face exprimée par un angle zygomaxillaire obtus ».
Les incisives en forme de pelle sont souvent citées comme preuve de la continuité régionale en Chine[51] - [52]. Stringer (1992) a cependant constaté que les incisives en forme de pelle sont présentes sur plus de 70 % des échantillons de fossiles de Wadi Halfa datés du début de l'holocène, et sont courantes ailleurs[53]. Frayer et al. (1993) ont critiqué la méthode de Stringer d’identification des incisives en forme de pelle. Ils insistent sur le fait qu'il existe différents degrés de « pelle » par exemple trace (+), semi (++), et marqué (+++), mais Stringer trompeusement à regrouper tout ensemble : «... combinant les catégories d’incisives en forme de pelle de cette manière est biologiquement un non-sens et trompeur. La statistique ne peut pas être correctement comparée avec les très hautes fréquences d’incisives en forme de pelle marqué (+++) trouvées chez les Asiatiques de l'Est »[33]. Le paléoanthropologue Fred H. Smith (2009) souligne également que : « c’est la catégorie d’incisives en forme de pelle qui permet d’identifier une caractéristique régionale d’Asie de l'Est, et pas seulement l'apparition d’incisives en forme de pelle de toute sorte »[54]. Les partisans de la théorie du multirégionalisme affirment que les d’incisives en forme de pelle marquées (+++) apparaissent seulement en Chine à une fréquence élevée, et ont moins de 10 % d'occurrence ailleurs.
La découverte en Chine, dans le Guangxi et le Yunnan, de fossiles humains semblant appartenir à une espèce archaïque, l'Homme de Maludong, mais datés de 11 à 14 000 ans avant le présent, semble montrer la coexistence de formes humaines archaïques et modernes pendant des périodes prolongées, ce qui va à l'encontre du multirégionalisme[55].
Europe
Dans les années 1990, David W. Frayer décrivait ce qu'il considérait comme un clade morphologique en Europe[56] - [57] - [58]. La séquence commence avec les plus anciens spécimens néandertaliens trouvés (crânes de Krapina et de Saccopastore), datant du Pléistocène moyen. Elle se poursuit avec la grotte de Vindija, et Cro Magnon ou avec les Européens récents de la fin du Paléolithique supérieur.
Frayer et al. (1993) considéraient qu'il y avait au moins quatre caractéristiques qui combinées sont uniques chez les fossiles européens : un foramen mandibulaire horizontal en forme ovale, un tubercule antérieure mastoïde, une fosse suprainiaque, et un rétrécissement de la largeur nasale associée à la réduction de la taille des dents. En ce qui concerne ce dernier, Frayer observe une continuité dans le rétrécissement nasal qui commence avec les Néandertaliens, se poursuit à la fin du Paléolithique supérieur et se retrouve sur les crânes de l'Holocène (Mésolithique). Ses conclusions sont contestées[59], mais ont reçu le soutien de Wolpoff, qui considère la forme nasale des spécimens des derniers néandertaliens comme « transitoire » entre les premiers Néandertaliens et les hommes de Cro-Magnon[60]. Sur la base d'autres similitudes crâniennes, Wolpoff et al . (2004) plaident en faveur d'une contribution importante de Neandertal aux Européens modernes[61].
On a trouvé en Europe quelques cas isolés d'hybridation entre Homo sapiens et l'Homme de Néandertal. Des fossiles hybrides présentant des traits anatomiques néandertaliens et modernes indiquent que le métissage restait possible en Europe[62] - [63] - [64]. Ces cas incluent l'enfant de Lapedo trouvé au Portugal[65] (qui est cependant contesté par certains[66]) et les fossiles de Peștera cu Oase en Roumanie[67].
Données génétiques
ADN mitochondrial
Une analyse de l'ADN mitochondrial de 147 personnes du monde entier réalisée pour la première fois en 1987 par Cann et al. a montré que leurs lignées mitochondriales étaient toutes issues d’un dernier ancêtre commun africain ayant vécu entre 140 000 et 200 000 ans AP[68]. L'analyse suggérait une expansion mondiale de l'homme moderne à partir du continent africain, remplaçant les hommes primitifs vivant hors d'Afrique. Un tel scénario de remplacement récent n’était pas compatible avec l'hypothèse multirégionale et les résultats de l'ADNmt ont conduit à donner beaucoup de crédit à la théorie du remplacement (out of Africa)[69] - [70] - [71].
Les conclusions tirées de l'analyse de l'ADN mitochondrial humain ont été néanmoins beaucoup débattues jusqu'au milieu des années 2000, époque à partir de laquelle les études ont pris en compte l'ADN nucléaire, moins sujet à contestations.
ADN nucléaire
Une analyse de 15 sites non codants sur le chromosome X a relevé ce qui pourrait ressembler à des incohérences. L'analyse a révélé une distribution multimodale des temps de coalescence du dernier ancêtre commun de ces sites. La coalescence semblait remonter pour certains gènes à près de 2 millions d'années (Ma), ce qui suggèrerait une séparation ancienne de populations. Alors que la plupart des sites du chromosome X montrent une plus grande diversité en Afrique, conformément aux origines africaines, quelques-uns des sites montrent une plus grande diversité en Asie plutôt qu'en Afrique. Pour quatre des 15 sites de gènes qui ne montrent pas une plus grande diversité en Afrique, la diversité variable des sites par région ne pourrait pas être expliquée par une simple migration depuis l'Afrique[72].
Des analyses supplémentaires des chromosomes X et de l'ADN autosomique ont allongé la liste des sites avec des temps de coalescence anciens[73] - [74] - [75] - [76] - [77]. La trop grande diversité génétique des sites fragiliserait la théorie d'une expansion récente depuis l'Afrique[78]. Des sites présentent également les deux caractéristiques en même temps : temps de coalescence trop ancien et diversité non compatible[79] - [80]. Par exemple, les analyses d'une région de RRM2P4 (ribonucléotide réductase M2 sous-unité pseudogène 4) a montré un temps de coalescence d'environ 2 millions d’années, avec une origine claire en Asie[81] - [82].
En 2001, une étude de l'ADN de plus de 12 000 hommes de 163 régions d’Asie de l'Est a montré que tous portaient une mutation apparue en Afrique il y a 35 000 à 89 000 ans. Ces « données ne confirment pas une contribution des hominidés archaïques in situ dans l’origine des humains modernes en Asie de l'Est »[83].
ADN ancien
Les analyses d'ADN prélevé directement sur des spécimens néandertaliens fossiles indiquent qu'ils ou leurs ancêtres ont contribué au génome de tous les êtres humains en dehors de l'Afrique, indiquant qu'il y a eu un certain degré de croisement avec les Néandertaliens avant leur remplacement[84]. Il a également été montré que l'Homme de Denisova avait contribué à l'ADN des Mélanésiens et des Australiens[85].
En 2010, le séquençage détaillé de l'ADN de plusieurs spécimens néandertaliens trouvés en Europe a montré que les Néandertaliens partageaient avec les humains non-africains modernes 1,8 % de leur génome, contre 0 % pour les Africains sub-sahariens[86] - [87]. Fin 2010, l’analyse de l'ADN dénisovien montre qu’il partage avec les mélanésiens d’aujourd’hui 3 % de son génome, contre environ 0,2 % en Asie orientale et 0 % ailleurs[88] - [89].
Les partisans de l'hypothèse multirégionale pensent que la combinaison de continuités régionales à l’intérieur et à l’extérieur de l’Afrique ainsi que les transferts latéraux de gènes entre les régions autour du monde sont des arguments forts en faveur de l’hypothèse multirégionale. Cependant, les promoteurs de la théorie « Out of Africa » pensent qu'il est possible d'expliquer ces résultats par le fait que les changements génétiques se produisent à une échelle régionale plutôt que continentale, et qu'ainsi les populations proches les unes des autres sont plus susceptibles de partager certains caractéristiques régionales spécifiques (polymorphisme d'un seul nucléotide - SNPs), tout en ayant la plupart des autres gènes en commun[90].
Notes et références
- Donald Johanson, Origins of Modern Humans : Multiregional or Out of Africa ?
- Discover : Not Out of Africa, Alan Thorne's challenging ideas about human evolution
- (en) Timothy D Weaver, « New developments in the genetic evidence for modern human origins », Evolutionary Anthropology: Issues, News, and Reviews, Wiley-Liss, vol. 17, no 1, , p. 69–80 (DOI 10.1002/evan.20161, lire en ligne)
- (en) NJ Fagundes, « Statistical evaluation of alternative models of human evolution », Proc Natl Acad Sci USA, vol. 104, no 45, , p. 17614–9 (PMID 17978179, PMCID 2077041, DOI 10.1073/pnas.0708280104, lire en ligne)
- Yves Coppens, Histoire de l'homme et changements climatiques, Fayard, 2006
- Jo Godefroid, Psychologie: science humaine et science cognitive, De Boeck Supérieur, 2001, p. 143
- (en) MH Wolpoff, « Multiregional, not multiple origins », Am J Phys Anthropol, vol. 112, no 1, , p. 129-36 (lire en ligne)
- (en) MH Wolpoff, « Modern Human Origins », Science, vol. 241, no 4867, , p. 772-4 (DOI 10.1126/science.3136545, lire en ligne)
- (en) Wolpoff M. H., « Modern human origins », Science, vol. 241, no 4867, , p. 772-774 (DOI 10.1126, lire en ligne)
- (en) Wolpoff, M. H., Wu, X. Z., & Alan, G. (86). G. Thorne, ‘Modern Homo Sapiens Origins : A General Theory of Hominid Evolution Involving the Fossil Evidence from east Asia’. The Origins of Modern Humans, New York, Liss, , p. 411-483
- (en) M. H. Wolpoff, J. N. Spuhler, F. H. Smith et J. Radovcic, « Modern human origins », Science, vol. 241, , p. 772–774 (ISSN 0036-8075 et 1095-9203, PMID 3136545, DOI 10.1126/science.3136545, lire en ligne, consulté le )
- Wolpoff, MH; Hawks, J; Caspari, R, « "Multiregional, not multiple origins" », American Journal of Physical Anthropology, no 112 (1), , p. 129-136 (DOI 10.1002/(SICI)1096-8644(200005)112:1<129::AID-AJPA11>3.0.CO;2-K, lire en ligne)
- (en) John Hawks et Milford H. Wolpoff, « Sixty Years of Modern Human Origins in the American Anthropological Association », American Anthropologist, vol. 105, , p. 89–100 (ISSN 1548-1433, DOI 10.1525/aa.2003.105.1.89, lire en ligne, consulté le )
- (en) R. B. Eckhardt, M. H. Wolpoff et A. G. Thorne, « Multiregional Evolution », Science, vol. 262, , p. 973–974 (ISSN 0036-8075 et 1095-9203, DOI 10.1126/science.262.5136.973-b, lire en ligne, consulté le )
- (en) R. Caspari et M. H. Wolpoff, « Weidenreich, Coon, and multiregional evolution », Human Evolution, vol. 11, , p. 261–268 (ISSN 0393-9375 et 1824-310X, DOI 10.1007/BF02436629, lire en ligne, consulté le )
- (en) Alan R. Templeton, « Genetics and Recent Human Evolution », Evolution, vol. 61, , p. 1507–1519 (ISSN 1558-5646, DOI 10.1111/j.1558-5646.2007.00164.x, lire en ligne, consulté le )
- (en) Wolpoff, M. H. and R. Caspari., Race and human evolution : A fatal attraction, New York, Simon and Schuster,
- (en) Wolpoff, M.H., and R. Caspari., « "The Many Species of Humanity" », Przegląd Antropologiczny (Anthropological Review), no 63 (1), , p. 3-17 (lire en ligne)
- (en) Weidenreich, F., « "Interpretations of the fossil material". In: Early Man in the Far East: Studies in Physical Anthropology », Studies in Physical Anthropology, Detroit, American Association of Physical Anthropologists, no 1, , p. 149-157
- (en) Wu, X., « "Origin of modern humans of China viewed from cranio-dental characteristics of late Homo sapiens" », Acta Anthropologica Sinica, no 17, , p. 276-282
- (en) Rosenberg, K. R., Wu, X., "A River Runs through It : Modern Human Origins in East Asia". In : The Origins of Modern Humans : Biology Reconsidered, Smith, F. H (ed). Wiley-Blackwell, , p. 89-122
- (en) Liu, L., Chen, X., He Archaeology of China : From the Late Paleolithic to the Early Bronze Age, Cambridge, Cambridge University Press, , p. 14
« "the majority of Chinese archaeologists and palaeontologists support[s] the multiregional development model, proposing a hypothesis of regional continuity with hybridization between immigrants and indigenous populations in the evolution from H. erectus to H. sapiens in East Asia." »
- (en) Leibold, J., "'Filling in the Nation : The Spatial Trajectory of Prehistoric Archaeology in Twentieth-Century China," in Transforming History : The Making of a Modern Academic Discipline in Twentieth Century China, Hong Kong, eds. Brian Moloughney and Peter Zarrow (Chinese University Press), , p. 333-371
- (en) Begun, DR., "The Past, Present and Future of Palaeoanthropology". In : A Companion to Paleoanthropology, Wiley-Blackwell, , p. 8
« "It needs to be noted, however, that this [Multiregional model] is a minority view among paleoanthropologists, most of whom support the African replacement model." »
- (en) Chris Stringer et Günter Bräuer, « Methods, Misreading, and Bias », American Anthropologist, vol. 96, , p. 416–424 (ISSN 1548-1433, DOI 10.1525/aa.1994.96.2.02a00080, lire en ligne, consulté le )
- (en) C. B. Stringer et P. Andrews, « Genetic and fossil evidence for the origin of modern humans », Science, vol. 239, , p. 1263–1268 (ISSN 0036-8075 et 1095-9203, PMID 3125610, DOI 10.1126/science.3125610, lire en ligne, consulté le )
- (en) Stringer, C. B., "Replacement, continuity and the origin of Homo sapiens". In : Continuity or replacement? Controversies in Homo sapiens evolution., Rotterdam: Balkema., F. H. Smith (ed)., , p. 9-24
- (en) Bräuer, G., & Stringer, C., "Models, polarization, and perspectives on modern human origins". In : Conceptual issues in modern human origins research., New York, Aldine de Gruyter, , p. 191-201
- (en) Stringer, C., « "Modern human origins—distinguishing the models" », Afr. Archaeol. Rev, no 18, , p. 67-75
- (en) Chris Stringer, « Modern human origins: progress and prospects », Philosophical Transactions of the Royal Society of London B: Biological Sciences, vol. 357, , p. 563–579 (ISSN 0962-8436 et 1471-2970, PMID 12028792, PMCID 1692961, DOI 10.1098/rstb.2001.1057, lire en ligne, consulté le )
- Chris Stringer, « Why we are not all multiregionalists now », Trends in Ecology & Evolution, vol. 29, , p. 248–251 (DOI 10.1016/j.tree.2014.03.001, lire en ligne, consulté le )
- (en) Wolpoff, M. H., Human evolution at the peripheries : the pattern at the eastern edge. Hominid Evolution : past, present and future, , p. 355-365
- (en) David W. Frayer, Mieford H. Wolpoff, Alan G. Thorne et Fred H. Smith, « Theories of Modern Human Origins: The Paleontological Test », American Anthropologist, vol. 95, , p. 14–50 (ISSN 1548-1433, DOI 10.1525/aa.1993.95.1.02a00020, lire en ligne, consulté le )
- (en) Wolpoff, M.H., A.G. Thorne, F.H. Smith, D.W. Frayer, and G.G. Pope, Multiregional Evolution : A World-Wide Source for Modern Human Populations. In : Origins of Anatomically Modern Humans, New York, M.H. Nitecki and D.V. Nitecki. Plenum Press, p. 175-199
- (en) Wolpoff, M. H., Multiregional evolution : the fossil alternative to Eden. In "The human revolution : behavioural and biological perspectives on the origins of modern humans", , p. 1: 62-108
- (en) Alan G. Thorne et Milford H. Wolpoff, « Regional continuity in Australasian Pleistocene hominid evolution », American Journal of Physical Anthropology, vol. 55, , p. 337–349 (ISSN 1096-8644, DOI 10.1002/ajpa.1330550308, lire en ligne, consulté le )
- (en) Thorne, A.G., "The Centre and the Edge : The significance of Australian hominids to African Palaeoanthropology". Proceedings of the 8th Pan-African Congress of Prehistory (Nairobi), Nairobi, National Museums of Kenya, , p. 180-181
- (en) Milford H. Wolpoff, John Hawks, David W. Frayer et Keith Hunley, « Modern Human Ancestry at the Peripheries: A Test of the Replacement Theory », Science, vol. 291, , p. 293–297 (ISSN 0036-8075 et 1095-9203, PMID 11209077, DOI 10.1126/science.291.5502.293, lire en ligne, consulté le )
- (en) « Abstracts of papers to be presented at the fifty-third annual meeting of the American association of physical anthropologists Philadelphia, Pennsylvania April 11–14, 1984 », American Journal of Physical Anthropology, vol. 63, , p. 133–238 (ISSN 1096-8644, DOI 10.1002/ajpa.1330630203, lire en ligne, consulté le )
- (en) Alan G. Thorne et Milford H. Wolpoff, « The Multiregional Evolution of Humans », Scientific American, vol. 266, , p. 76–83 (DOI 10.1038/scientificamerican0492-76, lire en ligne, consulté le )
- (en) Andrew Kramer, « Modern human origins in Australasia: Replacement or evolution? », American Journal of Physical Anthropology, vol. 86, , p. 455–473 (ISSN 1096-8644, DOI 10.1002/ajpa.1330860403, lire en ligne, consulté le )
- (en) Groves, C. P., "Thinking about Evolutionary Change The Polarity of Our Ancestors". In : Conceptual issues in modern human origins research, Transaction Publishers,
- (en) Habgood, P.J., "The origin of anatomically modern humans in Australasia". In : P. Mellars and C.B. Stringer (eds.) The Human Revolution : Behavioural and Biological Perspectives in the Origins of Modern Humans, , p. 245-273
- (en) Habgood, P.J., A Morphometric Investigation into the Origins of Anatomically Modern Humans, British Archaeological Reports, International Series 1176, Oxford England, Archaeopress,
- (en) Durband, A., « The view from down under: a test of the multiregional hypothesis of modern human origins using the basicranial evidence from Australasia », Collegium antropologicum, no 31 (3), , p. 651-659
- (en) Baba, H.; Aziz, F.; Narasaki, S., « Restoration of the face of Javanese Homo erectus Sangiran 17 and re-evaluation of regional continuity in Australasia », Acta Anthropologica Sinica, no 19, , p. 34-40
- (en) Wu, X., « The evolution of humankind in China », Acta Anthropologica Sinica, no 9 (4), , p. 312-321
- (en) Wu, X., Poirier, F. E., Human evolution in China : a metric description of the fossils and a review of the sites, New York, Oxford University Press, (lire en ligne)
- (en) Habgood, P.J., The origin of anatomically modern humans in east Asia Continuity or Replacement : Controversies in Homo sapiens evolution, G. Bräuer, and F.H. Smith (eds.), , p. 273-288
- (en) Groves, C. P., « A regional approach to the problem of the origin of modern humans in Australasia », The Human Revolution, Princeton, Princeton University Press,
- (en) Woo, R, « Chinese human fossils and the origin of Mongoloid racial group », Anthropos (Brno), no 23, , p. 151-155
- (en) Wu, X., « Evidence of Multiregional Human Evolution Hypothesis from China », Quaternary Sciences, no 26 (5), , p. 702-770
- (en) Stringer, C. B., « Replacement, continuity and the origin of Homo sapiens », Continuity or replacement? Controversies in Homo sapiens evolution, Rotterdam, , p. 9-24
- (en) Cartmill, M & Smith, F. H., The Human Lineage, Wiley-Blackwell, , 450 p.
- Darren Curnoe, Ji Xueping, Andy I. R. Herries et Bai Kanning, « Human Remains from the Pleistocene-Holocene Transition of Southwest China Suggest a Complex Evolutionary History for East Asians », PLoS ONE, vol. 7, , e31918 (PMID 22431968, PMCID 3303470, DOI 10.1371/journal.pone.0031918, lire en ligne, consulté le )
- (en) Frayer, D. W., « The persistence of Neanderthal features in post-Neanderthal Europeans », Continuity or Replacement: Controversies in Homo sapiens Evolution, Rotterdam, Balkema, , p. 179-188
- (en) Frayer, D. W., « Evolution at the European edge: Neanderthal and the Upper Paleolithic relationships », Préhistoire Européenne, no 2, , p. 9-69
- (en) Frayer, D. W., « Perspectives on Neanderthals as ancestors », Conceptual Issues in Modern Human Origins Research, New York, Aldine de Gruyter, , p. 220-235
- Nathan E. Holton et Robert G. Franciscus, « The paradox of a wide nasal aperture in cold-adapted Neandertals: a causal assessment », Journal of Human Evolution, vol. 55, , p. 942–951 (DOI 10.1016/j.jhevol.2008.07.001, lire en ligne, consulté le )
- (en) Wolpoff, M. H., « The place of Neanderthals in human evolution », The Emergence of Modern Humans: Biocultural Adaptations in the Later Pleistocene, Trinkaus, E (ed.). Cambridge University Press, , p. 97-141
- Milford Wolpoff, Bruce Mannheim, Alan Mann et John Hawks, « Why not the Neandertals? », World Archaeology, vol. 36, , p. 527–546 (ISSN 0043-8243, DOI 10.1080/0043824042000303700, lire en ligne, consulté le )
- (en) Erik Trinkaus, « European early modern humans and the fate of the Neandertals », Proceedings of the National Academy of Sciences, vol. 104, , p. 7367–7372 (ISSN 0027-8424 et 1091-6490, PMID 17452632, PMCID 1863481, DOI 10.1073/pnas.0702214104, lire en ligne, consulté le )
- « http://www.sciencedaily.com/releases/2007/04/070423185434.htm », sur www.sciencedaily.com (consulté le )
- Fred H. Smith, Ivor Janković et Ivor Karavanić, « The assimilation model, modern human origins in Europe, and the extinction of Neandertals », Quaternary International, vol. 137, , p. 7–19 (DOI 10.1016/j.quaint.2004.11.016, lire en ligne, consulté le )
- (en) Cidália Duarte, João Maurício, Paul B. Pettitt et Pedro Souto, « The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia », Proceedings of the National Academy of Sciences, vol. 96, , p. 7604–7609 (ISSN 0027-8424 et 1091-6490, PMID 10377462, DOI 10.1073/pnas.96.13.7604, lire en ligne, consulté le )
- (en) Ian Tattersall et Jeffrey H. Schwartz, « Hominids and hybrids: The place of Neanderthals in human evolution », Proceedings of the National Academy of Sciences, vol. 96, , p. 7117–7119 (ISSN 0027-8424 et 1091-6490, PMID 10377375, DOI 10.1073/pnas.96.13.7117, lire en ligne, consulté le )
- (en) Erik Trinkaus, Oana Moldovan, Ştefan Milota et Adrian Bîlgăr, « An early modern human from the Peştera cu Oase, Romania », Proceedings of the National Academy of Sciences, vol. 100, , p. 11231–11236 (ISSN 0027-8424 et 1091-6490, PMID 14504393, PMCID 208740, DOI 10.1073/pnas.2035108100, lire en ligne, consulté le )
- (en) Rebecca L. Cann, Mark Stoneking et Allan C. Wilson, « Mitochondrial DNA and human evolution », Nature, vol. 325, , p. 31–36 (DOI 10.1038/325031a0, lire en ligne, consulté le )
- (en) William A. Haviland, Harald E. L. Prins, Dana Walrath et Bunny McBride, Essence of Anthropology, Cengage Learning, , 408 p. (ISBN 978-0-495-59981-4, lire en ligne)
- (en) Renée Hetherington et Robert G. B. Reid, The Climate Connection : Climate Change and Modern Human Evolution, Cambridge University Press, (ISBN 978-1-139-48436-7, lire en ligne)
- (en) Martin Meredith, Born in Africa : The Quest for the Origins of Human Life, PublicAffairs, (ISBN 978-1-58648-663-1, lire en ligne)
- Michael F. Hammer, Daniel Garrigan, Elizabeth Wood et Jason A. Wilder, « Heterogeneous patterns of variation among multiple human x-linked Loci: the possible role of diversity-reducing selection in non-africans », Genetics, vol. 167, , p. 1841–1853 (ISSN 0016-6731, PMID 15342522, PMCID 1470985, DOI 10.1534/genetics.103.025361, lire en ligne, consulté le )
- (en) Toshiyuki Hayakawa, Ikuko Aki, Ajit Varki et Yoko Satta, « Fixation of the Human-Specific CMP-N-Acetylneuraminic Acid Hydroxylase Pseudogene and Implications of Haplotype Diversity for Human Evolution », Genetics, vol. 172, , p. 1139–1146 (ISSN 0016-6731 et 1943-2631, PMID 16272417, PMCID 1456212, DOI 10.1534/genetics.105.046995, lire en ligne, consulté le )
- (en) Eugene E. Harris et Jody Hey, « X chromosome evidence for ancient human histories », Proceedings of the National Academy of Sciences, vol. 96, , p. 3320–3324 (ISSN 0027-8424 et 1091-6490, PMID 10077682, DOI 10.1073/pnas.96.6.3320, lire en ligne, consulté le )
- Hie Lim Kim et Yoko Satta, « Population genetic analysis of the N-acylsphingosine amidohydrolase gene associated with mental activity in humans », Genetics, vol. 178, , p. 1505–1515 (ISSN 0016-6731, PMID 18245333, PMCID 2278054, DOI 10.1534/genetics.107.083691, lire en ligne, consulté le )
- Daniel Garrigan, Zahra Mobasher, Sarah B. Kingan et Jason A. Wilder, « Deep haplotype divergence and long-range linkage disequilibrium at xp21.1 provide evidence that humans descend from a structured ancestral population », Genetics, vol. 170, , p. 1849–1856 (ISSN 0016-6731, PMID 15937130, PMCID 1449746, DOI 10.1534/genetics.105.041095, lire en ligne, consulté le )
- (en) Rosalind M. Harding, « More on the X files », Proceedings of the National Academy of Sciences, vol. 96, , p. 2582–2584 (ISSN 0027-8424 et 1091-6490, PMID 10077551, DOI 10.1073/pnas.96.6.2582, lire en ligne, consulté le )
- Audrey Sabbagh, André Langaney, Pierre Darlu et Nathalie Gérard, « Worldwide distribution of NAT2 diversity: implications for NAT2 evolutionary history », BMC genetics, vol. 9, , p. 21 (ISSN 1471-2156, PMID 18304320, PMCID 2292740, DOI 10.1186/1471-2156-9-21, lire en ligne, consulté le )
- Etienne Patin, Luis B. Barreiro, Pardis C. Sabeti et Frédéric Austerlitz, « Deciphering the ancient and complex evolutionary history of human arylamine N-acetyltransferase genes », American Journal of Human Genetics, vol. 78, , p. 423–436 (ISSN 0002-9297, PMID 16416399, PMCID 1380286, DOI 10.1086/500614, lire en ligne, consulté le )
- (en-US) « Variation in NAT1 and NAT2 », sur john hawks weblog (consulté le )
- (en) Molecular Biology and Evolution Society, American Society of Naturalists et Society for the Study of Evolution, « Molecular biology and evolution. », Molecular biology and evolution., (ISSN 0737-4038, lire en ligne, consulté le )
- Murray P. Cox, Fernando L. Mendez, Tatiana M. Karafet et Maya Metni Pilkington, « Testing for archaic hominin admixture on the X chromosome: model likelihoods for the modern human RRM2P4 region from summaries of genealogical topology under the structured coalescent », Genetics, vol. 178, , p. 427–437 (ISSN 0016-6731, PMID 18202385, PMCID 2206091, DOI 10.1534/genetics.107.080432, lire en ligne, consulté le )
- (en) Yuehai Ke, Bing Su, Xiufeng Song et Daru Lu, « African Origin of Modern Humans in East Asia: A Tale of 12,000 Y Chromosomes », Science, vol. 292, , p. 1151–1153 (ISSN 0036-8075 et 1095-9203, PMID 11349147, DOI 10.1126/science.1060011, lire en ligne, consulté le )
- Vania Yotova, Jean-Francois Lefebvre, Claudia Moreau et Elias Gbeha, « An X-linked haplotype of Neandertal origin is present among all non-African populations », Molecular Biology and Evolution, vol. 28, , p. 1957–1962 (ISSN 1537-1719, PMID 21266489, DOI 10.1093/molbev/msr024, lire en ligne, consulté le )
- David Reich, Nick Patterson, Martin Kircher et Frederick Delfin, « Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania », American Journal of Human Genetics, vol. 89, , p. 516–528 (ISSN 1537-6605, PMID 21944045, PMCID 3188841, DOI 10.1016/j.ajhg.2011.09.005, lire en ligne, consulté le )
- (en) Richard E. Green, Johannes Krause, Adrian W. Briggs et Tomislav Maricic, « A Draft Sequence of the Neandertal Genome », Science, vol. 328, , p. 710–722 (ISSN 0036-8075 et 1095-9203, PMID 20448178, DOI 10.1126/science.1188021, lire en ligne, consulté le )
- (en-US) « NEANDERTALS LIVE! », sur john hawks weblog (consulté le )
- David Reich, Richard E. Green, Martin Kircher et Johannes Krause, « Genetic history of an archaic hominin group from Denisova Cave in Siberia », Nature, vol. 468, , p. 1053–1060 (ISSN 0028-0836, PMID 21179161, PMCID 4306417, DOI 10.1038/nature09710, lire en ligne, consulté le )
- (en-US) « The Denisova genome FAQ », sur john hawks weblog (consulté le )
- (en) D. J. Witherspoon, S. Wooding, A. R. Rogers et E. E. Marchani, « Genetic Similarities Within and Between Human Populations », Genetics, vol. 176, , p. 351–359 (ISSN 0016-6731 et 1943-2631, PMID 17339205, PMCID 1893020, DOI 10.1534/genetics.106.067355, lire en ligne, consulté le )
Bibliographie
- (en) AR Templeton, « Out of Africa again and again », Nature, vol. 416, , p. 45–51
- (en) Osbjorn M Pearson, « Has the Combination of Genetic and Fossil Evidence Solved the Riddle of Modern Human Origins? », Evolutionary Anthropology, vol. 13, , p. 145-159
- (en) J Adams, « Human Evolutionary Tree », Nature Education, Macmillian, vol. 1, no 1, (lire en ligne)
- (en) Donald C Johanson, « Origins of Modern Humans: Multiregional or Out of Africa? », ActionBioscience, (consulté le )