Nettie Stevens
Nettie Maria Stevens est une généticienne américaine, née le à Cavendish (Vermont) et morte à Baltimore (Maryland) le . Elle a fait l'une des grandes découvertes biologiques du début du XXe siècle : que le sexe de chaque individu est déterminé par des caractères chromosomiques. Vivant à une époque où toutes les femmes scientifiques étaient des exceptions et devaient tailler leur place[1], elle s'est fait reconnaître pour ses contributions scientifiques.

| Naissance |
Cavendish (Vermont) (États-Unis) |
|---|---|
| Décès |
Baltimore (Maryland) (États-Unis) |
| Nationalité |
|
| Domaines | Génétique, Embryologie |
|---|---|
| Institutions |
Westford Academy Westfield Normal School Université Stanford Bryn Mawr College |
| Diplôme |
Université Stanford Bryn Mawr College |
| Directeur de thèse | Thomas Hunt Morgan |
| Renommée pour | La découverte du système XY de détermination sexuelle |
La contribution de Nettie Stevens à la science génétique naissante fut très importante bien que largement inconnue, en partie à cause de la notoriété de ses mentors, Thomas Hunt Morgan et Edmund Beecher Wilson, et en partie à cause de son décès précoce.
Biographie
Nettie Maria Stevens naît le 7 juillet 1861 à Cavendish, petite ville du Vermont où Alexandre Soljenitsyne trouvera refuge un siècle plus tard. Elle est la troisième des quatre enfants d'Ephraïm Stevens et de Julia Adams. Ses deux frères aînés sont morts à l'âge de quelques mois en 1858 et 1860. Sa mère meurt à 27 ans, six mois après la naissance de sa sœur Emma, en 1863[2].
Ephraïm Stevens se remarie avec Ellen C. Thompson en 1865. Il s'installe à Westford[3], près de Lowell dans le Massuchusetts, à 40 km au Nord-Ouest de Boston. Il est charpentier et gagne suffisamment bien sa vie pour acquérir des propriétés diverses et pouvoir payer des études à ses enfants. Nettie Stevens fait ses études secondaires à la Westford Academy entre 1872 et 1880. Élève brillante, elle est toujours en tête de sa classe. À la fin de ses études, elle est institutrice dans un lycée à Lebanon dans le New Hampshire pendant trois ans. Elle enseigne les mathématiques, l'anglais et le latin, matières obligatoires, et aussi la zoologie et la physiologie[3]. Son intérêt pour la biologie est peut-être né à l'occasion d'un cours pour enseignants qu'elle a suivi à Martha's Vinyard en 1880.
Ayant pris goût à l'enseignement, elle entre en 1883 à la Westfield Normal School (aujourd’hui la Westfield State University (en)) en banlieue de Spingfield dans le Massachusetts. Elle accomplit en deux ans le cursus qui normalement dure quatre ans[3]. D'après sa courte notice nécrologique sur le site du cimetière de Westford, elle passe les dix années suivantes comme enseignante et assistante du Principal à Westford Academy[4]. Au cours de cette période, en 1888, sa mère d'adoption, Ellen Thomson meurt à 46 ans.
Elle a 35 ans quand, en 1896, elle décide d'entamer un cursus universitaire de biologie à l’université Stanford, en Californie, qui vient d'être inaugurée en 1891 et qui a la particularité d'accepter des étudiantes aussi bien que des étudiants. De plus à Stanford, les professeurs sont jeunes et dynamiques, et les étudiants peuvent choisir les cours qui les intéressent autour d'une matière principale[5]. Il est possible aussi que son choix de Stanford ait été conseillé par son ancien professeur et directeur de Westfield Normal School, biologiste et élève de Louis Agassiz, célèbre biologiste, ichtyologiste, paléontologue suisse établi aux États-Unis. En effet, le président de Stanford, David Starr Jordan, est aussi élève de Louis Agassiz[5].
En première année, Nettie Stevens a Oliver Peebles Jenkins, qui enseigne la physiologie, comme professeur principal. En deuxième année, elle travaille sous la direction du Professeur Frank Mace McFarland, qui a huit ans de moins qu'elle. McFarland est professeur associé d'histologie et sa recherche porte sur le rôle du noyau cellulaire dans l'hérédité. McFarland revient d'un séjour de deux ans en Europe où il a travaillé à l'université de Wurtzbourg dans le laboratoire de Théodore Boveri ainsi qu'à la Station Zoologique de Naples pour préparer sa thèse de doctorat[5].
Nettie Stevens obtient son Bachelor of Arts en 1899. Son père et sa sœur Emma font le déplacement pour assister à la soutenance de son diplôme. Ils sont séduits par la Californie et s'installent peu après à Mountain View, à 10 km de Stanford. Nettie quitte la résidence universitaire où elle loge et rejoint sa famille pour la fin de ses études[5]. Elle prépare son diplôme de Master of Arts qu'elle présente en 1900. Son travail est intitulé Studies on Ciliate Infusoria et constitue sa première publication scientifique[5].
Pendant ses études à Stanford, elle passe quatre étés (de 1897 à 1900) à la station marine Hopkins Seaside Laboratory installée dès 1891 à Pacific Grove, sur la baie de Monterey. L'été suivant, elle est à Woods Hole. En 1901-1902, elle passe un semestre à la Station Zoologique de Naples et elle y revient en 1909. Elle visite aussi le laboratoire de Cold Spring Harbour, Long Island, New York. Elle a donc toujours accès aux espèces marines auxquelles elle consacre de nombreuses études.
En 1900, Nettie Stevens a 39 ans. Elle revient sur la côte Est des États-Unis et poursuit ses recherches en cytologie au Bryn Mawr College, établi en périphérie de Philadelphie. C'est une université prestigieuse pour les filles qui peuvent acquérir des diplômes jusqu'au doctorat[3]. Bryn Mawr est un lieu de recherche réputé en biologie depuis qu'Edmund B. Wilson y a enseigné. Wilson est le fondateur de biologie cellulaire américaine. Son livre The cell in development and inheritance, paru en 1896, réédité en 1900 et en 1925, a eu un grand retentissement[6]. Wilson a quitté Bryn Mawr pour Columbia University à New York en 1891. C'est son élève et successeur, Thomas H. Morgan, le futur fondateur de la génétique classique, qui accueille Nettie Stevens et accepte de superviser sa thèse. Comme à Stanford, l'étudiante a quatre ans de plus que son professeur. Bryn Mawr lui accorde une bourse pour sa première année de recherche.
En 1901, Bryn Mawr lui offre une bourse d'études en Europe. Comme Wilson, Morgan et McFarland l'ont fait avant elle, elle va travailler dans le laboratoire de Théodore Boveri à l'Université de Wurtzbourg et à la Station Zoologique de Naples. Elle rencontre à Wurtzbourg Marcella O'Grady, ancienne élève de Wilson à Bryn Mawr, qui a épousé Boveri en 1898. De retour aux États-Unis, Nettie Stevens rejoint Bryn Mawr, passe son doctorat en 1903 et poursuit ses recherches sur place avec un statut précaire de Research Fellow. Elle tente d'obtenir un statut permanent de chercheuse à plein temps de la Carnegie Institution à Washington avec les recommandations chaleureuses de Wilson, Morgan et du directeur de Bryn Mawr College. Sa demande est refusée en 1903, faute d'argent dans les caisses, mais elle est acceptée l'année suivante.
Elle reste à Bryn Mawr avec les titres de Lectrice puis d'Associée en Morphologie Expérimentale. En 1908-1909, elle fait un nouveau voyage en Europe à la Station Zoologique de Naples avec un financement de l'American Association of University Women[7]. En reconnaissance de sa contribution à l'enseignement et à la recherche dans l'institution pendant de longues années, la direction de Bryn Mawr College décide en 1912, de la création, pour Nettie Stevens, d'une chaire de professeur avec peu d'obligations d'enseignement qui lui aurait permis d'avoir du temps pour ses recherches[3]. Malheureusement, elle n'a pas la possibilité d'occuper cette chaire. Nettie Stevens meurt d'un cancer du sein à l'âge de 50 ans à l'Hôpital John Hopkins de Baltimore, le 4 mai 1912[2]. Elle est inhumée dans le cimetière Fairview de Westford. Son père et sa sœur, morts respectivement en 1914 et 1945 à Mountain View, Californie, sont enterrés à ses côtés[8].
« Sa détermination et son dévouement, combinés à un grand sens de l'observation, son sérieux et sa patience, unis à un solide jugement, expliquent en partie sa remarquable réussite » (Thomas Hunt Morgan[9]).
Contributions
Dans son premier travail publié, Nettie Stevens décrit une nouvelle espèce d'infusoires (des protozoaires ciliés). Elle met en œuvre les techniques d'histologie les plus perfectionnées de l'époque. Elle étudie les modifications des noyaux au cours de la division cellulaire et met en évidence pour la première fois des chromosomes chez les protozoaires[5]. Elle poursuit ses travaux sur les infusoires à Bryn Mawr, travaux regroupés dans sa thèse sur Further studies on ciliate infusoria Lichnophora and Boveria.
Dans les années 1880, Hermann Henking (de), Privat-docent à l'université de Göttingen, remarqua un « élément » qui persistait durant la plus grande partie de la première division méiotique chez les cellules germinales de l'insecte appelé gendarme, ou pyrrhocore (Pyrrhocoris apterus). Cet élément fut appelé « chromosome accessoire »[10].
C'est au Bryn Mawr College que Nettie Stevens mène ses recherches, dont l'une aboutira, en 1905, à la découverte du rôle joué par le chromosome Y dans la détermination sexuelle. Edmund Beecher Wilson fait une découverte similaire, indépendamment, la même année[11]. Elle utilise, pour ses expériences, divers insectes et identifie le chromosome Y chez un coléoptère du genre Tenebrio. Elle déduit alors que la base de la détermination du sexe dépend de la présence ou de l’absence du chromosome Y.
Studies on spermatogenesis. Part I
.jpg.webp)
L'étude sur la spermatogenèse publiée en 1905 est la publication la plus célèbre de Nettie Stevens. Elle reprend l'hypothèse de McClung[12] et explore la présence du « chromosome accessoire » en relation avec le sexe de l'individu. Pour mener son enquête, elle examine les cellules des testicules de 5 espèces d'insectes de la classe des néoptères chez lesquels aucune étude de la spermatogenèse n'a encore été menée. Comme le fait remarquer McClung dans son introduction: « On a constaté qu'on trouve rarement dans une espèce tous les détails d'un problème avec la même clarté. Utiliser différentes espèces permet de mettre en évidence certains traits plus apparents dans une espèce que dans les autres. » Dans les cellules germinales, elle dénombre et observe les chromosomes au cours de la mitose et de la méiose. Elle utilise des techniques de coloration spécifiques de la chromatine pour distinguer les chromosomes des autres structures cellulaires. Elle mène ses observations avec un microscope monoculaire et dessine à la main toutes les images.
Termopsis angusticollis
Chez ce termite de Californie, Nettie Stevens trouve que les cellules des gonades sont petites et qu'il faut employer le plus fort grossissement possible de son microscope (x1500, avec l'objectif à immersion à huile) pour distinguer des choses intéressantes. Les chromosomes sont très nombreux (52). Elle repère des tétrades en losange. Elle n'observe pas de chromosome accessoire.
Stenopelmatus
Ce criquet des sables appartient à l'ordre des Orthoptères, étudié par McClung[2]. Il possède 46 chromosomes dont 2 sont plus longs que les autres. Nettie Stevens remarque un petit chromosome, qu'elle appelle "élément x", et qui n'est présent que dans la moitié des spermatides.
Blattella germanica
Dans ce cafard commun, il a été difficile de compter les chromosomes des cellules diploïdes . Dans les spermatides (haploïdes), Stevens en compte 23 dont un chromosome bizarre, élément x, qui n'est présent que dans la moitié des cellules.
Tenebrio molitor
La larve de ce coléoptère constitue les vers de farine. C'est avec cet animal que Nettie Stevens fait sa découverte. Ci-dessous, on trouvera la traduction intégrale de la section de l'article original consacré au Ténébrion meunier avec les figures qui s'y rattachent.

« Dans toutes les mitoses des spermatogonies en métaphase dans lesquelles il était possible de compter avec précision les chromosomes, on a trouvé 20 chromosomes, 19 grands de taille approximativement égale, et un petit sphérique (fig. 169 et 170, élément « s »). Dans les spermatocytes les plus jeunes, on trouve occasionnellement des cellules formant un kyste avec des noyaux comme ceux des figures 171 et 172, indiquant qu'une brève « synapse », ou étape de condensation, se produit à la fin de la dernière mitoses des spermatogonies. Pendant la plus grande partie de la période, la chromatine forme un lourd spirème irrégulier et parfois segmenté (fig. 173 et 174). Peu après la première division de maturation, des segments clivés de ce type peuvent être observés comme dans la figure 175. Certains apparaissent comme des tétrades avec des bandes de connexion étroites entre les éléments appariés. On trouve aussi quelques cas, comme dans la figure 176, où le spirème est segmenté en chromosomes bivalents, chaque composant montrant une fracture longitudinale. Cette figure montre aussi le petit chromosome. »

« Cependant, d'ordinaire, le spirème irrégulier et enchevêtré (fig. 173 et 174) se condense en une bande fortement segmentée disposée de diverses façons dans le noyau (fig. 177). Cette bande se fractionne rapidement en chromosomes bivalents montrés sur les figures 178 et 179, donnant 9 paires symétriques et une paire asymétrique, composée du petit chromosome et d'un plus grand. En prophase du fuseau, dans de rares cas, certains chromosomes sont clivés longitudinalement et condensés transversalement, formant des tétrades (fig. 180). Mais la plupart du temps, ils apparaissent comme dans la figure 181. La paire asymétrique qui apparaît dans chaque figure est marquée « s ». Elle est la dernière à s'aligner sur la plaque équatoriale en métaphase (fig. 182), peut-être en raison de son manque de symétrie. Le petit composant de cette paire est toujours dirigé vers l'équateur du fuseau. La figure 183 montre une coupe tangentielle d'un fuseau en métaphase contenant une paire inégale et une paire égale.
Dans la figure 184, la vue polaire d'une métaphase est montrée. La paire inégale est un peu décalée par rapport aux autres. Les figures 184a et 185 montrent que les composants inégaux de la paire asymétrique, ainsi que les composants égaux des paires symétriques, sont séparés en métakinèse, manifestant clairement une division de réduction. Deux plaques polaires sont montrées dans les figures 186 et 187, l'une contenant 10 éléments égaux et l'autre contenant 9 éléments égaux et un petit. La télophase est montrée à la figure 188. Il n'y a pas d'étape de repos. Le nouveau fuseau est formé des restes du précédent, et la masse de la chromatine en forme de fuseau, vue à la figure 188, ou bien passe dans le centre du nouveau fuseau ou bien est enveloppée par lui. Les chromosomes doubles se séparent comme dans les figures 189 et 190. »
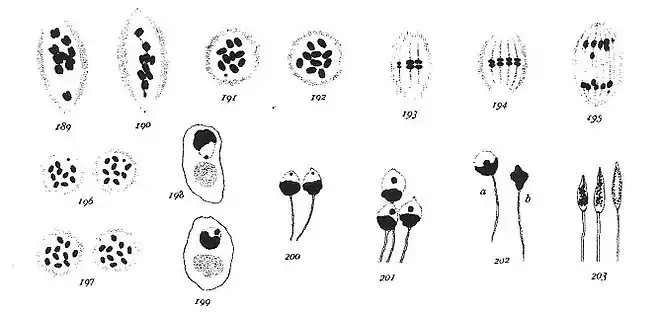
« La figure 190 montre la petite dyade et la figure 189 une dyade aberrante qui est peut-être sa jumelle. Dans les deux divisions, le fuseau est spécial en ce qu'il a, à l'extérieur, une masse dense de fibres qui, lorsque la préparation est fixée à l'acide osmique, se colore fortement avec l'Hématoxyline ferrique. Ces fibres apparaissent sur toutes les figures de 174 à 196. Les figures 191 et 192 sont les plaques équatoriales des deux types de spermatocytes de second ordre. Le petit chromosome apparaît sur la figure 191. Un début d'anaphase apparaît sur les figures 193 et 194 qui montrent toutes les deux le petit et les grands chromosomes en métakinèse. La figure 195 est une fin d'anaphase contenant le petit chromosome divisé.
Dans la figure 196, on voit les deux plaques polaires correspondant à la figure 195 et, dans la figure 197, les plaques polaires d'un fuseau dans lequel 10 chromosomes égaux ont été divisés. Chez Tenebrio molitor, les spermatides sont donc certainement de deux types différents en ce qui concerne leur contenu en chromatine. Dans la plupart des jeunes spermatides, après que la membrane nucléaire se soit formée, il apparaît un élément chromatinien isolé qui correspond assez bien soit au grand, soit au petit composant de la paire asymétrique, séparés lors de la première mitose et divisé dans la seconde. La portion claire du noyau contenant cet élément est d'abord tourné vers le reste du fuseau (fig. 198) mais, avant que la queue apparaisse, soit tout le noyau, soit son contenu, tourne à 180° (fig. 199). Différentes étapes du développement des spermatides sont montrées dans les figures 200 à 203.
Avant de déterminer, si possible, la valeur de la paire asymétrique d'éléments chromatiniens, des ovaires très jeunes, avec des tubes d'œufs, ont été sectionnés. Les chromosomes ont été comptés dans les cellules en division du follicule (cellules somatiques femelles) et dans les ovogonies. Dans les deux cas, on a trouvé 20 grands chromosomes. La figure 207 montre la plaque équatoriale d'une cellule somatique femelle d'un jeune follicule. Les figures 208a et b montrent deux coupes d'une ovogonie dans la prophase de la mitose. Pour déterminer le nombre et le type des chromosomes dans les cellules somatiques des mâles, plusieurs pupes mâles ont été sectionnés. Dans les spermatogonies on a trouvé 19 grands et un petit chromosomes. La figure 204 montre la plaque équatoriale d'une cellule somatique en division et les figures 205 et 206 montrent les plaques filles d'une cellule semblable (Trois grands chromosomes de la plaque de la figure 206 sont dans une autre coupe). »

« À partir de ces faits, il apparaît que le pro-noyau de l'œuf doit, dans tous les cas, contenir 10 grands chromosomes tandis que le spermatozoïde, à la fertilisation, apporte à l'œuf soit 10 grands chromosomes, soit 9 grands et un petit. Puisque les cellules somatiques des femelles contiennent 20 grands chromosomes tandis que celles des mâles contiennent 19 grands et un petit, cela semble un cas clair de détermination sexuelle, non pas par un chromosome accessoire, mais par une vraie différence de caractère des éléments d'une paire des chromosomes des spermatocytes de premier ordre. Les spermatozoïdes qui contiennent le petit chromosome déterminent le sexe mâle, pendant que ceux qui contiennent 10 chromosomes de taille égale détermine le sexe femelle. Ce résultat suggère qu'il peut y avoir, dans beaucoup de cas, une différence intrinsèque affectant le sexe dans le caractère de la chromatine de la moitié des spermatozoïdes, bien qu'il puisse ne pas toujours apparaître par des différences de taille des chromosomes aussi marquées que chez Tenebrio. Il est important que des espèces apparentées soient étudiées de façon à savoir si des conditions chromatiques semblables apparaissent dans d'autres espèces de ce genus ou peut-être chez les coléoptères en général. (Note: Le Professeur E.B. Wilson a récemment trouvé un dimorphisme similaire dans les spermatozoïdes de Lygeus et de Hemiptera heteroptera.) »
Aphis oenotherae
Nettie Stevens a publié une étude détaillée des cellules germinales des pucerons (Studies on the germ cells of aphids). Elle en donne un résumé dans l'étude présente. Elle observe des chromosomes de différentes longueurs mais ne détecte pas de chromosome à l'aspect ou au comportement qui pourrait ressembler à un chromosome accessoire.
Sagitta bipunctata
Sagitta est un vers aquatique. À certaines étapes de la prophase, un petit chromosome au comportement particulier pourrait correspondre au chromosome accessoire, élément x. Mais, comme Sagitta est hermaphrodite, on ne peut pas parler de détermination sexuelle par un chromosome particulier. La fonction de l'élément x reste à découvrir.
Discussion
La traduction de la partie de la discussion générale qui a trait à Tenebrio est reproduite ci-dessous.
| Position des taxons étudiés par Nettie Maria Stevens | |
|
« Du point de vue de la détermination sexuelle, nous avons chez Tenebrio molitor l'espèce la plus intéressante considérée dans ce papier. Dans les cellules somatiques aussi bien que germinales des deux sexes, il y a une différence, non pas du nombre d'éléments chromatiniens, mais de la taille de l'un qui est très petit chez le mâle et de la même grandeur que les 19 autres chez la femelle. Les noyaux des œufs de la femelle doivent être semblables quant au nombre et à la taille des chromosomes, tandis qu'il est absolument certain que les spermatozoïdes se répartissent en deux catégories égales relativement au contenu chromatinien des noyaux. Une moitié d'entre eux ont 9 grands chromosomes et un petit, tandis que l'autre moitié a 10 grands chromosomes.
Puisque les cellules somatiques des mâles ont 19 grands et un petit chromosome, tandis que les cellules somatiques des femelles ont 20 grands chromosomes, il semble certain qu'un œuf fertilisé par un spermatozoïde contenant le petit chromosome doit produire un mâle, et qu'un œuf fertilisé par un spermatozoïde contenant 10 grands chromosomes de même taille doit produire une femelle. Le petit chromosome lui-même peut ne pas être un déterminant sexuel mais les conditions trouvées chez Tenebrio indiquent que le sexe peut, dans certains cas, être déterminé par une différence dans la quantité ou la qualité de la chromatine de différents spermatozoïdes. Ceci constitue la partie la plus suggestive du travail et sera suivi de l'étude d'autres espèces apparentées.
Il apparaît qu'il y a très peu d'uniformité dans la présence d'hétérochromosomes, même chez les insectes, et dans leur apparence quand ils sont présents. La poursuite de la discussion doit donc être reportée en attendant que la spermatogenèse de beaucoup d'autres espèces soit minutieusement explorée.
Bryn Mawr College, 15 mai 1905 »
Autres travaux
Elle aborde aussi à Bryn Mawr des questions comme la régénération chez les organismes multicellulaires primitifs, la structure des organismes unicellulaires, le développement du sperme et des ovules, les cellules germinales des insectes et la division cellulaire chez les Echinoidea et les vers.
Publications (liste partielle)
On compte une quarantaine de publications de Nettie M.Stevens[13] - [14].
- (en) Nettie M. Stevens, Studies on ciliate infusoria, Palo Alto, Hopkins Marine Station,
- (en) Nettie M. Stevens, Further studies on ciliate infusoria, Lichnophora and Boveria (Thèse), Naumberg O.S.,
- (en) Nettie M. Stevens, « On the germ cells and the embryology of Planaria simplissima », Proc. Acad. Nat. Sci., , p. 208-220 + Planches 13-16
- (en) Nettie M. Stevens, « Studies on germ cells of Aphis rosae and Aphis oenotherae », J. Exp. Zool., 1905 vol.2, p. 313-333
- (en) Nettie M. Stevens, Studies in spermatogenesis (Part 1) with especial reference to the "Accessory Chromosome", Washington, Carnegie Institution of Washington, Publication n°36, , 33 p. (lire en ligne)
- (en) Nettie M. Stevens, Studies in spermatogenesis (Part 2) A comparative study of the heterochromosomes in certain species of coleoptera, hemiptera and lepidoptera, witl especial reference to sex determination, Washington, Carnegie Institution of Washington, Publication n°36 part 2, , 43 p.
- (en) Nettie M. Stevens, Studies on the germ cells of aphids, Washington, Carnegie Institution of Washington, , 44 p. (lire en ligne)
- (en) Nettie M. Stevens, « Studies of the germ cells of certain diptera with reference to the heterochromosomes and the phenomena of synapsis », J. Exp. Zool., 1908 vol.5, p. 359-574
- (en) Nettie M. Stevens, « A note on reduction in the maturation of male eggs in Aphis », Biol. Bull., 1910 vol.18, p. 72-75
- (en) Nettie M. Stevens, « A preliminary note on heterochromosomes in the guinea pig », Biol. Bull., 1911 vol.20, p. 121-122
- (en) Nettie M. Stevens, « Further studies on heterochromosomes in mosquitoes », Biol. Bull., 1911 vol.21, p. 109-120
- (en) Nettie M. Stevens, « Heterochromosomes in the guinea pig », Biol. Bull., 1911 vol.21, p. 155-167
- (en) Nettie M. Stevens, « Supernumerary chromosomes and synapsis in Ceuthophilus (SP?) », Biol. Bull., 1910 vol.22, p. 219-230
- (en) Nettie M. Stevens, « Further observations on supernumerary chromosomes and sex ratios in diabrotica soror », Biol. Bull., 1912 vol.22, p. 231-238
Bibliographie
- (en) « Nettie Maria Stevens (1861–1912) »
- (en) Stephen G. Brush, « Nettie M. Stevens and the discovery of sex determination by chromosomes », Isis, juin 1978, vol.69, p. 163-172[15]
- (en) Joel B. Hagen, Douglas Allchin and Fred Singer, Nettie Stevens and the problem of sex determination, in Doing Biology, Minneapolis, Pearson, , p. 37-47
Émission
- [(fr) Cherchez la femme ! (16/30) Nettie Stevens - La naissance de la génétique (page consultée le 17 janvier 2023)], série en stop motion de Julie Gavras (2021), Arte, durée 0 h 4.
Compléments
Hommages
- Nettie Stevens a reçu de son vivant l’Alice Freeman Palmer Fellowship, pour un an de recherches, de l’American Association of University Women (1908)[7].
- Elle est inscrite depuis 1994 au National Women's Hall of Fame[16].
- Le , Google lui dédie un doodle[17] pour le 155e anniversaire de sa naissance.
- La Westfield State University a nommé un nouveau bâtiment de laboratoires pour ses étudiants: Dr Nettie Maria Stevens Science and Innovation Center[18].
Notes et références
- Angeline Durand-Vallot, « Nettie Stevens et la découverte de la détermination chromosomique du sexe », dans Adeline Gargam, Femmes de sciences de l'Antiquité au XIXe siècle : réalités et représentations, Éditions universitaires de Dijon, , 274 p. (ISBN 978-2-36441-081-7), p. 274.
- Simone Gilgenkrantz, « Nettie Maria Stevens (1861-1912) », Médecine/Sciences, 2008, vol.24, p. 874-878 (lire en ligne)
- (en) « Nettie Maria Stevens », sur vermonthistory.org (consulté le )
- (en) Marilyn Bailey Ogilvie, Joy Dorothy Harvey and Margaret Rossiter, The biogaphical dictionary of women in science, New York(USA)/Oxon (GB), Rootledge, , 1491 p. (ISBN 978-0-415-92038-4, lire en ligne), p. 1230-1231
- (en) Patricia C. Cross and John P. Steward, « Nettie maria Stevens. Turn-of-the-century Stanford alumna paved path for women in biology », Sandstone & Tile, 1993 vol. 17, p. 3-12 (lire en ligne)
- (en) Edmund Beecher Wilson, The cell in develoment and inheritance, New York, MacMillan, , 483 p.
- (en) « American Association of University Women », sur aauw.com (consulté le )
- (en) « Find a grave », sur new.findagrave.com (consulté le )
- « Her single-mindedness and devotion, combined with keen powers of observation; her thoughtfulness and patience, united to a well-balanced judgment, account, in part, for her remarkable accomplishment. » T. H. Morgan, « The scientific work of Miss N. M. Stevens », Science, vol. 36, no 298, , p. 468–70 (DOI 10.1126/science.36.928.468, JSTOR 1636618). On se souviendra d'elle, ajoute Morgan, dans une notice nécrologique développée.
- (en) Frederic B. Churchill, August Weissmann, Devopment, Heredity and Evolution, Cambridge, Mass., Harvard University Press, , 722 p. (lire en ligne), p. 424
- E. B. Wilson, « The chromosomes in relation to determination of sex in insects », dans Science, 22, p. 500–502 (1905).
- (en) Clarence Erwin McClung, « The accessory chromosome - Sex Determinant? », Biol. Bull., 1902 vol.1 (lire en ligne)
- (en) Marilyn Bailey Ogilvie and Clifford J. Choquette, « Nettie Maria Stevens (1861-1912): Her life and contribution to cytogenetics », Proc. Am. Philos. Soc., 1981, vol.125, p. 292-311
- (en) « Résultats pour 'au:Stevens, N. M.' [WorldCat.org] », sur www.worldcat.org (consulté le )
- Accès payant, The University of Chicago Press Journals.
- « Nettie Stevens », sur le site de l'organisation.
- Doodle du .
- (en) « Dr. Nettie Maria Stevens | Westfield State University », sur www.westfield.ma.edu (consulté le )
Liens externes
- Ressource relative à la recherche :